5-HT1A-рецептор
5-HT1A-рецептор — подтип серотониновых рецепторов подсемейства 5-HT1-рецепторов. Эндогенным лигандом-агонистом для них является моноаминовый нейромедиатор серотонин (5-гидрокситриптамин, 5-HT). Рецепторы этого подтипа относятся к семейству трансмембранных метаботропных G-белок-связанных рецепторов и связаны с гетеротримерным ингибиторным G-белком, так называемым Gi / Go. Этот подтип рецепторов опосредует тормозящую нейротрансмиссию. Ген, кодирующий белок этого рецептора у человека, обозначается HTR1A.[1][2]
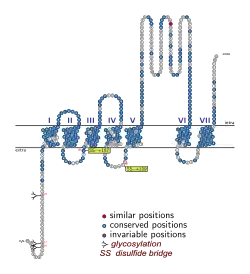
Структура 5-HT1A-рецептора
Серотониновый рецептор подтипа 5-HT1A является белком (вернее гликопротеином), состоящим у человека из 422 аминокислот (молекулярная масса 46107 дальтон). Подобно другим G-белок-связанным рецепторам, он имеет семь трансмембранных доменов и семь α-спиральных доменов, среди которых находится и активный сайт рецептора — место связывания с лигандами, такими, как серотонин. Обращённая к синапсу сторона имеет небольшой отрицательный электростатический заряд (что способствует электростатическому притяжению положительно заряженных лигандов), а обращённая внутрь клетки сторона имеет небольшой положительный электростатический заряд, способствующий связыванию с отрицательно заряженным сайтом Gi.
Гликозилируется по аспарагину в позициях 10, 11, 24 (Asn10, Asn11, Asn24). Связывается с убиквитином в позиции лизина 334 (Lys334).
Первичная, вторичная и третичная структуры белка 5-HT1A-рецептора проявляют высокую степень аминокислотно-последовательностной и структурной гомологичности с первичной, вторичной и третичной структурами других G-белок-связанных рецепторных белков, в частности родопсина и особенно β₂-адренорецептора. Именно на базе аминокислотной и структурной гомологичности с родопсином были построены первые пространственные модели 5-HT1A-рецептора. Позднее эти пространственные модели были усовершенствованы с использованием в качестве гомологичной модели β₂-адренорецептора, проявляющего более высокую степень аминокислотно-последовательностной, структурной и функциональной гомологичности с 5-HT1A-рецептором.[3]
Белок 5-HT1A-рецептора взаимодействует с липидами мембраны клетки, в частности холестерином и сфинголипидами,[4] приобретая при взаимодействии с холестерином более плотную пространственную конфигурацию и большее сродство к агонистам.[3]
Белок рецептора 5-HT1A подвергается и другим посттрансляционным модификациям, а именно — пальмитированию (ковалентному соединению тиоэфирной связью с остатками пальмитиновой кислоты) в специфических, эволюционно высоко консервативных участках аминокислотной последовательности (что подтверждает важность этого пальмитирования для функционирования 5-HT1A-рецептора) — в области остатков цистеина в позициях 417 и 420, находящихся в проксимальном С-терминальном домене рецептора. Показано, что отсутствие пальмитирования в любом из двух участков — 417 или 420 — значительно понижает функциональную активность 5-HT1A-рецептора, а именно его способность связываться с гетеротримерным G-белком Gi и угнетать активность аденилатциклазы. При отсутствии пальмитирования одновременно в обоих цистеинах — 417 и 420 — способность 5-HT1A-рецептора связываться с α-субъединицей Gi —белком Giα полностью утрачивается. При отсутствии пальмитирования одновременно в обоих цистеинах 417 и 420 также полностью утрачивается и функциональная активность 5-HT1A-рецептора, в частности его способность угнетать стимулированное форсколином повышение аденилатциклазной активности и накопление циклического аденозинмонофосфата (цАМФ) в клетке. Это даёт основания полагать, что пальмитирование цистеиновых остатков в позициях 417 и 420 критически необходимо для обеспечения функциональной активности 5-HT1A-рецептора и его способности связываться с Gi и оказывать влияние на активность аденилатциклазного нисходящего эффекторного пути. Кроме того, зависящая от активации 5-HT1A-рецептора активация ERK-сигнального пути также оказалась нарушенной у мутантного белка, лишённого способности к пальмитированию в цистеиновых остатках 417 и 420. Это заставляет предполагать, что пальмитирование белка 5-HT1A-рецептора в цистеиновых остатках 417 и 420 важно также для обеспечения его способности к передаче сигнала через βγ-субъединицы G-белка (димер Giβγ) и ERK-сигнальный путь, в дополнение к важности этого пальмитирования для обеспечения способности к передаче сигнала через Giα и аденилатциклазный путь.[5]
Также было показано, что пальмитирование белка 5-HT1A-рецептора в области цистеиновых остатков 417 и 420 необходимо для его правильного позиционирования в специфических местах клеточной мембраны, обогащённых холестерином и сфинголипидами — так называемых липидных рафтах. Показано также, что правильное позиционирование 5-HT1A-рецептора именно в этих специфических местах клеточной мембраны, а не в произвольных её местах (и, соответственно, необходимое для этого пальмитирование цистеиновых остатков 417 и 420) важно для правильного функционирования 5-HT1A-рецептора и эффективной передачи им сигнала внутрь клетки.[6]
Два идущих подряд остатка лейцина в позициях 414 и 415 C-терминального конца 5-HT1A-рецептора критически важны для правильного трёхмерного пространственного фолдинга этого гликопротеина, для распознавания им агонистов и для правильного размещения 5-HT1A-рецептора на поверхности тела нейрона и на его дендритах (в то время как 5-HT1B-рецепторы преимущественно находятся на аксонах). Двухточечная мутация с заменой 414 и 415 лейцинов на соответствующие остатки аланина приводит к образованию нефункционального белка, который секвестрируется в эндоплазматическом ретикулуме клетки (то есть не транспортируется к клеточной мембране и не встраивается в неё), не способен распознавать агонисты и имеет резко пониженную степень гликозилирования. В то же время замена пальмитированных цистеинов 417 и 420 на серины приводит к меньшим нарушениям функциональности 5-HT1A-рецептора.[7]
Белок 5-HT1A-рецептора в культуре клеток по-разному гликозилируется в разных типах клеток, что влияет на возможность распознавания его теми или иными антителами при иммуногистохимическом исследовании тканей.[8]
Эволюционно высоко консервативный остаток треонина в 149-й позиции C-терминального конца (внутриклеточная петля i2), являющийся также известным местом фосфорилирования 5-HT1A-рецептора протеинкиназой C играет роль в правильной передаче опосредованного G-белком Gi сигнала. В частности, мутантный белок 5-HT1A-рецептора с заменой треонина в 149-й позиции на аланин (T149A) проявляет резко пониженную способность регулировать уровень внутриклеточного кальция — эффект, опосредуемый βγ-субъединицами G-белка, а также несколько пониженную способность угнетать активность аденилатциклазы и снижать внутриклеточное накопление цАМФ — эффект, опосредуемый α-субъединицей G-белка. Это позволяет предположить, что именно этот участок рецептора ответствен за специфическое взаимодействие с G-белком.[9]
Специфические аминокислотные остатки в трансмембранных доменах 4 и 5 (TM4/TM5) — остаток триптофана в 175-й позиции (Trp175 (4.64)), остаток тирозина в 198-й позиции (Tyr198 (5.41)), идущие подряд два остатка аргинина в 151-й и 152-й позиции (Arg151 (4.40) и Arg152 (4.41)) являются специфическим интерфейсом для димеризации 5-HT1A-рецептора.[10]
Гетеродимеризация 5-HT1A-рецептора
Рецепторы подтипа 5-HT1A формируют G-белок-связанные гетеродимеры со следующими рецепторами: 5-HT7-рецептор,[11] 5-HT1B, 5-HT1D, ГАМКB2, GPCR26, LPA1, LPA3, S1P1, S1P3.[12]
Распределение в организме
Рецептор подтипа 5-HT₁A является наиболее распространённым в человеческом организме подтипом серотониновых рецепторов. В центральной нервной системе рецепторы подтипа 5-HT₁A в большом количестве обнаруживаются в коре больших полушарий головного мозга, в гиппокампе, септуме, миндалевидном теле (амигдале) и других структурах лимбической системы, в ядрах шва. Меньшие количества 5-HT₁A-рецепторов обнаруживаются также в базальных ганглиях и в таламусе.[13][14][15] Рецепторы подтипа 5-HT₁A в ядрах шва представлены в основном соматодендритными ауторецепторами, в то время как в других областях мозга, таких, как гиппокамп, они в основном представлены постсинаптическими рецепторами.[14]
Пути передачи внутриклеточного сигнала
Аденилатциклазный путь

Уже в ранних исследованиях было показано, что 5-HT₁A-рецептор связан с одним из семейства ингибиторных G-белков — Gi / Go.[16] Связывание агониста с рецептором вызывает замену гуанозиндифосфата (ГДФ) на гуанозинтрифосфат (ГТФ) в α-субъединице белка Gi / Go.[17] Это вызывает, соответственно, активацию этой α-субъединицы (её переход в активную форму). А одна из основных функций активной формы Giα / Goα (хотя и не единственная) — это угнетение активности внутриклеточной аденилатциклазы. Угнетение активности аденилатциклазы вызывает снижение образования в клетке циклического АМФ из АТФ. А это вызывает угнетение активности цАМФ-зависимой протеинкиназы — так называемой протеинкиназы A. Эксперименты с мембранами гиппокампальных нейронов млекопитающих показали, что серотонин, а также такие агонисты 5-HT₁A-рецепторов, как 8-OH-DPAT, 5-карбоксамидотриптамин (5-CT), урапидил, буспирон, ипсапирон, способны ингибировать вызванное форсколином увеличение активности аденилатциклазы и соответственно предотвращать накопление циклического АМФ именно благодаря воздействию на 5-HT₁A-рецепторы.[16][18] Позже этот эффект был также воспроизведён в культурах гиппокампальных и корковых нейронов,[19] а также в других клетках, экспрессирующих 5-HT₁A-рецептор.[20][21][22]
В мозгу, однако, лишь активация 5-HT₁A-гетерорецепторов, расположенных на не-серотониновых нейронах, приводит с Giα-связанному ингибированию активности аденилатциклазы и к угнетению образования цАМФ и активности протеинкиназы А. Показано, что 5-HT₁A-ауторецепторы, расположенные на серотониновых нейронах в ядрах шва, не ингибируют аденилатциклазную активность и не связываются с аденилатциклазой.[25] В разных областях мозга 5-HT₁A-рецепторы связаны с разными ингибиторными белками семейства Gi / Go. Например, в дорсальной части ядер шва 5-HT₁A-ауторецепторы на серотониновых нейронах чаще всего связаны с Gi3α, а 5-HT₁A-гетерорецепторы в гиппокампе — с Goα.[26] Кроме того, показано, что способность 5-HT₁A-ауторецепторов ядер шва к десенситизации значительно выше, чем способность 5-HT₁A-гетерорецепторов в лимбической системе к аналогичной десенситизации.[27][28][29] Это может отражать различия в их связывании с различными сигнальными G-белками, а также и само по себе может оказывать влияние на различия в их способе передачи внутриклеточного сигнала и на эффективность этой передачи.
Агонисты 5-HT₁A-рецепторов, такие, как 8-OH-DPAT, вызывая понижение активности протеинкиназы А в гиппокампе, вызывают вследствие этого повышение активности протеин фосфатазы 1 (одной из мишеней протеинкиназы А) и снижение активности фосфорилирования кальций/кальмодулин-зависимой протеинкиназы II (CaMKII) — а эта активность повышается при процессах обучения. Показано, что именно этот сигнальный каскад ответствен за наблюдаемые при воздействии 8-OH-DPAT нарушения памяти и обучаемости вследствие активации 5-HT1A-рецепторов.[30] Таким образом, ингибирование аденилатциклазы и активности протеинкиназы А и опосредованное через него ингибирование активности протеин-фосфатазы-1 и кальций/кальмодулин-зависимой протеинкиназы II (CaMKII) может опосредовать поведенческие эффекты активации 5-HT1A-рецепторов.
Модуляция активности калиевых каналов и электрической активности клеток
Активация 5-HT₁A-рецепторов также активирует G-белок-связанные калиевые ионные каналы внутреннего выпрямления — так называемые GIRK[31] — причём и в гиппокампе,[32][33][34] и в дорсальной части ядер шва.[25][35] То есть этот эффект активации G-белок-связанных калиевых каналов внутреннего выпрямления (GIRK) — одинаков у 5-HT₁A-ауторецепторов и у 5-HT₁A-гетерорецепторов. Учитывая тот факт, что активация 5-HT₁A-ауторецепторов в ядрах шва не вызывает ингибирования аденилатциклазы,[25] маловероятно, что выраженная активация входящего тока ионов калия в дорсальной части ядер шва, наблюдаемая при воздействии 5-HT₁A-агонистов, является цАМФ-зависимым эффектом. Активация GIRK и соответственно активация входящего тока ионов калия осуществляется другим механизмом — а именно, диссоциировавшими под воздействием активации рецептора из связи с α-субъединицей β- и γ-субъединицами Gi / Go — то есть димером Giβγ / G0βγ.[36] Способность активированных 5-HT₁A-рецепторов вызывать GIRK-индуцированный гиперполяризующий входящий ток ионов калия позволяет 5-HT₁A-рецепторам оказывать сильнейшее тормозящее влияние на электрическую возбудимость нейронов, на их способность к генерации электрических импульсов.[35] Этот физиологический процесс также может иметь прямое отношение к поведенческим эффектам, наблюдаемым при активации 5-HT₁A-рецепторов,[37] в частности к их анксиолитическому, анальгетическому, антидепрессивному эффектам.
Фосфолипазный путь
Активация 5-HT₁A-рецепторов также может вызывать активацию фосфолипазы C, которая катализирует образование из фосфатидилинозитол дифосфата (PIP2) двух вторичных мессенджеров — диацилглицерина (DAG) и инозитолтрифосфата (IP3). Далее происходит активация протеинкиназы C. А протеинкиназа C, в свою очередь, фосфорилирует многие белки нисходящего эффекторного каскада.
Помимо этого, в этом каскаде инозитолтрифосфат (IP3) связывается со специфическими IP3-рецепторами, многие из которых являются кальциевыми каналами (например, кальмодулин), что приводит к увеличению концентрации внутриклеточного кальция и активации ряда кальций-зависимых внутриклеточных процессов.
Взаимодействие с системой синтазы оксида азота
Активация 5-HT₁A-рецепторов оказывает стимулирующее влияние на активность эндогенной системы оксида азота (II) (моноксида азота, NO) и синтазы оксида азота (белка NOS). Так, например, в культуре клеток вентральной части простаты крысы, так же как и в живом организме крысы, активация 5-HT₁A-рецепторов приводит не только к ожидаемому (и типичному для разных клеточных систем) угнетению активности аденилатциклазы, но и к повышению активности синтазы оксида азота.[38]
Некоторые СИОЗС угнетают активность синтазы оксида азота, в том числе через вызываемую ими десенситизацию 5-HT₁A-рецепторов (понижение их чувствительности к стимуляции). Это может быть одной из причин наблюдаемых при воздействии СИОЗС нарушений сексуальной функции, поскольку активность эндотелиальной формы синтазы оксида азота и вызываемое ею выделение оксида азота (II) критически важны для вазодилатации кровеносных сосудов полового члена, и, соответственно, для акта эрекции.[39]
Вызываемая некоторыми β-адреноблокаторами, такими, как небиволол, вазодилатация также во многом опосредуется через их парциальный агонизм к 5-HT₁A-рецепторам, и соответственно вызываемую ими стимуляцию 5-HT₁A-рецепторов и индукцию синтазы оксида азота, приводящую к повышению биосинтеза оксида азота (II). Этот эффект предотвращается антагонистами 5-HT₁A-рецепторов, такими, как метерголин или NAN-190, или блокаторами синтазы оксида азота, такими, как метиловый эфир N-ω-нитро-L-аргинина (L-NAME), а также теми β-адреноблокаторами, которые обладают, напротив, антагонистической активностью в отношении 5-HT₁A-рецепторов, как, например, пропранолол.[40]
Регуляция циркадного ритма у млекопитающих происходит путём передачи информации об освещённости от специализированных клеток сетчатки в супрахиазматические ядра гипоталамуса (так называемые SCN) по ретиногипоталамическому (оптикохиазмальному) пути с участием глутаматергических нейронов, причём в процессе передачи сигнала важно участие как NMDA-рецепторов, так и не-NMDA глутаматных рецепторов и активация синтазы оксида азота, приводящая к выделению оксида азота (II). А этот процесс, включающий выделение оксида азота (II), как показано, модулируется активностью постсинаптических 5-HT₁A и 5-HT7-рецепторов и пресинаптических 5-HT₁B-гетерорецепторов.[41] Таким образом, вызываемая активацией 5-HT₁A-рецепторов индукция синтазы оксида азота и повышение биосинтеза оксида азота (II) может принимать непосредственное участие в реализации их поведенческих эффектов и, например, частично объяснять наблюдаемую при воздействии антидепрессантов нормализацию нарушенного при депрессиях циркадного ритма.
Система синтазы оксида азота может также принимать участие в опосредованной 5-HT₁A-рецепторами регуляции аппетита и пищевого поведения. В частности, у крыс наблюдаемая при воздействии агониста 5-HT₁A-рецепторов — 8-OH-DPAT стимуляция аппетита и гиперфагия, снимается воздействием блокатора синтазы оксида азота — метилового эфира N-ω-нитро-L-аргинина (L-NAME), однако этот эффект стимуляции аппетита и вызывания гиперфагии при помощи 8-OH-DPAT восстанавливается при введении получившим L-NAME крысам прекурсора оксида азота (II) — L-аргинина.[42]
Однако влияние стимуляции 5-HT₁A-рецепторов на активность синтазы оксида азота и на выделение оксида азота (II) различно в разных типах нейронов. Так, например, в гиппокампальных нейронах взрослой крысы стимуляция 5-HT₁A-рецепторов серотонином или селективным парциальным агонистом 5-HT₁A-рецепторов — буспироном — приводит к угнетению вызванной активацией NMDA-рецепторов активности синтазы оксида азота и к снижению выделения оксида азота (II) и накопления циклического ГМФ, а также к угнетению вызванного активацией NMDA-рецепторов высвобождения арахидоновой кислоты и накопления ионов кальция, причём этот эффект серотонина и буспирона снимается селективным антагонистом 5-HT₁A-рецепторов — NAN-190, но не снимается антагонистом 5-HT2-рецепторов — кетансерином. Предполагается, что этот эффект является одним из механизмов, благодаря которому серотонин и антидепрессанты оказывают нейропротективное влияние на нейроны гиппокампа, защищая их от NMDA-, кальций- и NO-опосредованной эксайтотоксичности.[43]
Эксперименты на мышах показывают, что повышение проницаемости кровеносных сосудов, вызываемое серотонином, как эндогенным медиатором воспаления, также частично опосредуется через вызываемую стимуляцией 5-HT₁A-рецепторов (но в ещё большей мере — стимуляцией 5-HT2-рецепторов) кровеносных сосудов индукцию синтазы оксида азота и выделение оксида азота (II). Этот эффект серотонина на проницаемость кровеносных сосудов сильно блокируется антагонистом 5-HT2-рецепторов — кетансерином и неселективным антагонистом 5-HT₁- и 5-HT2-рецепторов метисергидом, в меньшей степени блокируется селективным антагонистом 5-HT₁A-рецепторов — NAN-190, но не блокируется селективным антагонистом 5-HT3-рецепторов — гранисетроном (что означает вовлечение в реализацию эффекта как 5-HT2, так и 5-HT₁A-рецепторов, но неучастие 5-HT3-рецепторов), и предотвращается блокаторами синтазы оксида азота — L-NAME и метиленовым синим.[44]
Способность серотонина и агонистов 5-HT₁A-рецепторов, таких, как 5-карбоксамидотриптамин (5-CT) или 8-OH-DPAT, расширять кровеносные сосуды почек тоже опосредуется через стимуляцию 5-HT₁A-рецепторов эндотелия сосудов, индукцию синтазы оксида азота и выделение оксида азота (II). Этот эффект блокируется антагонистами 5-HT₁A-рецепторов BMY-7378 и метерголином, а также снимается ингибиторами синтазы оксида азота — L-NAME и метиленовым синим.[45]
Способность серотонина и агониста 5-HT₁A-рецепторов 8-OH-DPAT ингибировать прессорный ответ на симпатическую стимуляцию также опосредуется через индукцию синтазы оксида азота и выделение оксида азота (II) и блокируется ингибитором гуанилатциклазы ODQ и/или ингибитором синтазы оксида азота — L-NAME.[46]
И наоборот, воздействие на систему оксида азота (II) и синтазы оксида азота оказывает влияние на активность 5-HT₁A-рецепторов.
Сложности изучения сигнальных каскадов 5-HT₁A-рецептора
Несмотря на то, что связь 5-HT₁A-рецепторов с опосредованными Giα / Goα изменениями активности аденилатциклазы, концентрации внутриклеточного цАМФ и активности протеинкиназы А и с опосредованными Giβγ / Goβγ изменениями активности GIRK и тока ионов калия уже хорошо установлена, функция этих двух «канонических» сигнальных механизмов и их роль в опосредованной 5-HT₁A-рецепторами регуляции активности нейронов всё ещё не изучена достаточно полно. Сложность механизмов внутриклеточной передачи сигнала в нейронах требует комбинированных фармакологических, биохимических и молекулярно-биологических подходов к их изучению для детальной характеристики каждого компонента внутриклеточного сигнального каскада и его роли и места в общей схеме. А это сильно ограничивает возможности изучения этих каскадов in vivo и возможность охарактеризовать их влияние на функции нейронов в их естественной среде, на функцию мозга в целом и на поведение живых организмов как целого. Помимо этого, в работе 5-HT₁A-рецепторов могут играть роль также дополнительные, «неканонические» сигнальные каскады. Среди них в последнее время наибольшее внимание привлекают сигнальные каскады, работа которых традиционно ассоциировалась с активацией рецепторов для факторов роста, и которые также оказались связаны с работой 5-HT₁A-рецепторов.
Сигнальный путь MAPK/ERK
Роль MAP-киназ в регуляции роста клеток и в поддержании их жизнеспособности, в противодействии апоптозу разных типов клеток хорошо известна.[47] Более того, хорошо известно, что MAP-киназы являются критически важными в регуляции роста, развития и пластичности также и нейронов центральной нервной системы. Семейство MAP-киназ включает в себя активируемые внеклеточными сигналами киназы 1 и 2 (ERK1 и ERK2, известные также как белки MAP-киназ p42 и p44), MAP-киназу p38 (p38-MAPK) и c-Jun N-терминальную киназу (JNK). Из всего семейства MAP-киназ, активация 5-HT₁A-рецепторов наиболее сильно влияет на активность ERK1 и ERK2. Обычно ERK1/2 активируются тирозинкиназами, связанными с рецепторами факторов роста. Эти рецепторы активируют малую ГТФазу Ras. Белок Ras, в свою очередь, активирует белок Raf1. А уже этот белок фосфорилирует и активирует MAPK/ERK-киназы 1 и 2 (MEK1 и MEK2). А MEK1 и MEK2 — это протеинкиназы, регулирующие активность ERK1 и ERK2. Активация MEK приводит к фосфорилированию ERK. Активация ERK приводит к множественной активации протеинкиназ нисходящего ERK-сигнального пути, таких, как рибосомальная S6 киназа (RSK), и к активации факторов транскрипции белков, таких, как Myc, и белков-активаторов митоза, таких, как потенциальный онкоген en:Elk1. Фосфорилирование белков нисходящего ERK-сигнального пути в нейронах приводит к множественной активации разнообразных рецепторов и ионных каналов, множественной экспрессии различных генов и к проявлениям нейропластичности. Все эти явления могут иметь внешние поведенческие проявления. Одним из интересных примеров является активация фактора транскрипции CREB — одной из мишеней (субстратов) ERK-киназ — серин/треониновой протеинкиназой RSK. Белок фактора транскрипции CREB хорошо изучен. Изучена его роль в регуляции экспрессии генов и роль изменений в его активности и соответствующих изменений в регулируемой им экспрессии генов в развитии таких патологических состояний, как стресс, тревожные состояния, депрессия. Регуляция активности CREB при помощи ERK-сигнального пути заставляет предполагать, что сигнальный путь ERK может играть важную роль в регуляции настроения, уровня тревожности, уровня стресса. Поведенческие эффекты изменений в активности ERK-сигнального пути исследовались в нескольких лабораториях. Было показано, что ингибиторы MEK вызывают множественные изменения в поведении животных, в зависимости от вида животного вызывая гиперактивное состояние, повышение или снижение уровня тревоги и стресса и поведение, напоминающее проявления депрессии. Также было показано, что ингибиторы MEK блокируют поведенческие эффекты антидепрессантов. Столь большое разнообразие поведенческих эффектов ингибиторов МЕК может быть следствием того, что MEK/ERK-сигнальный путь влияет на активность одновременно множества регуляторных белков и факторов транскрипции, являющихся субстратами ERK-киназ. Для того, чтобы точнее определить поведенческие эффекты, связанные с активацией ERK-сигнального каскада, необходимо продолжение исследований и более точное определение роли каждого из этих белков в регуляции как функции отдельной клетки, так и поведения организма в целом.
Способность 5-HT₁A-рецепторов активировать ERK1/2 при помощи их фосфорилирования была впервые обнаружена в клетках, отличных от нейронов, но также экспрессирующих 5-HT₁A-рецепторы. Это воздействие 5-HT₁A-рецепторов на активность ERK1/2 чувствительно к ингибированию Gi-белков коклюшным токсином. Это заставляет предполагать, что в активации ERK-сигнального пути при стимуляции 5-HT₁A-рецепторов принимает участие G-белок-связанный механизм. Так же, как и при вызванной факторами роста активации ERK, вызванная активацией 5-HT₁A-рецепторов активация ERK опосредуется активацией малых ГТФ-аз Ras и Raf и активацией через них MEK. Этот сигнальный каскад требует кальмодулин-зависимого эндоцитоза рецепторов как промежуточного этапа. Кроме того, активация ERK1/2 при активации 5-HT₁A-рецепторов в ненейронных клетках может также опосредоваться фосфатидилинозитол-3-киназой (PI3K) и фосфатидилхолин-специфичной фосфолипазой С (PLC). Этот эффекторный каскад также является G-белок-зависимым. Однако детали процесса передачи сигнала от G-белок-зависимой фосфатидилинозитол-3-киназы (PI3K) к ERK в результате активации 5-HT₁A-рецепторов ещё до конца не понятны.
Несмотря на то, что в ненейронных клеточных системах активация 5-HT₁A-рецепторов систематически и закономерно приводит к активации сигнального каскада MEK/ERK, эффекты активации 5-HT₁A-рецепторов на активность MEK/ERK каскада и в частности на активность ERK1/2 в клетках нейронального происхождения вариабельны в зависимости от места происхождения и степени дифференцировки нейронов. Так, в частности, в высокодифференцированных HN2-5 клетках, происходящих из гиппокампа, агонисты 5-HT₁A-рецепторов вызывают усиление фосфорилирования ERK1/2 и повышение её активности. Показано, что этот эффект зависит от активности малых ГТФ-аз Ras и Raf, активации MEK и мобилизации внутриклеточного кальция. Однако этот эффект активации 5-HT₁A-рецепторов не обнаруживается в первичной культуре низкодифференцированных гиппокампальных нейронов или в ромбоэнцефалических нейронах эмбриона. Для них активация 5-HT₁A-рецепторов нейтральна в отношении активности ERK1/2 (не влияет на неё никак). А в высокодифференцированных нейрональных клетках из ядер шва активация 5-HT₁A-рецепторов приводит к прямо противоположному эффекту — опосредуемому Gβγ-субъединицей ингибированию активности MEK и снижению фосфорилирования и активности ERK. На то, каким образом отреагирует MEK/ERK-сигнальный каскад на активацию 5-HT₁A-рецепторов, влияют многие факторы. В частности, похоже на то, что для проявления активирующего действия 5-HT₁A-рецепторов на активность ERK необходима высокая плотность 5-HT1A-рецепторов на поверхности клеток (что чаще встречается у зрелых, высокодифференцированных клеток). Однако тип 5-HT₁A-рецепторов (ауторецепторы, как в ядрах шва, или гетерорецепторы, как, например, в гиппокампе), их предпочтительное связывание с тем или иным подтипом Gi / Go ингибиторных белков, и наличие и доступность этих белков в соответствующих клетках также, по-видимому, оказывают влияние на характер ERK-ответа на активацию 5-HT₁A-рецепторов. Хотя факторы, детерминирующие тот или иной тип ERK-ответа на активацию 5-HT₁A-рецепторов ещё до конца не изучены, важным уже достигнутым результатом исследований на сегодняшний день является понимание того, что опосредуемая 5-HT₁A-рецепторами регуляция активности MEK/ERK-сигнального пути высоко селективна и специфична в зависимости от области происхождения нейронов, их возраста, степени дифференцировки. Это особенно важно в свете того, что в мозгу существует огромное количество разнообразных типов нейронов, экспрессирующих 5-HT₁A-рецепторы и огромное количество разных областей с существенно разными физиологическими функциями, содержащих такие нейроны.
В нескольких исследованиях было убедительно показано, что активация MEK/ERK-сигнального пути при активации 5-HT₁A-рецепторов не является универсальной реакцией для любых нейронов мозга. В частности, в ряде исследований было показано, что 5-HT₁A-агонисты быстро, но кратковременно повышают фосфорилирование ERK в гипоталамусе, и что этот эффект, по всей вероятности, является промежуточным этапом в вызываемой 5-HT₁A-агонистами индукции биосинтеза и высвобождения окситоцина, АКТГ и пролактина. В контрасте с этим, 5-HT₁A-агонисты понижают фосфорилирование ERK и активность ERK в гиппокампе. И хотя значение факта этой отрицательной привязки активности ERK к активности 5-HT₁A-рецепторов в гиппокампе точно не известно, но сама по себе ERK известна как критически важный регулятор процессов мышления и памяти и как важный медиатор синаптической пластичности. Вероятно, ингибирование 5-HT₁A-рецепторами активности ERK в гиппокампе может играть роль в опосредуемых 5-HT₁A-рецепторами изменениях синаптической пластичности или в объяснении вызываемых активацией 5-HT₁A-рецепторов нарушений когнитивных функций. Находки, характеризующие регуляцию 5-HT₁A-рецепторами фосфорилирования ERK в других областях мозга, таких, как кора больших полушарий, миндалевидное тело, ядра шва, значительно более противоречивы. Так, например, по некоторым данным, во фронтальной коре 5-HT₁A-агонисты повышают фосфорилирование ERK, а по другим данным, там они не влияют на степень фосфорилирования ERK. В префронтальной коре активация 5-HT₁A-рецепторов или NMDA-рецепторов по отдельности никак не влияет на активность ERK, однако одновременная активация обоих типов рецепторов приводит к снижению фосфорилирования ERK и к понижению её активности. Это заставляет предполагать, что для регуляции активности ERK в нейронах коры больших полушарий критически важно кросс-общение между различными нейромедиаторами, рецепторами и сигнальными механизмами, совместно координирующее эффект. Для более чёткого понимания взаимосвязи между активностью 5-HT₁A-рецепторов и активностью сигнального каскада MEK/ERK в коре больших полушарий нужны более детальные исследования этих связей в различных областях коры и различных типах нейронов коры. По контрасту с исследованиями, которые показали, что в изолированных высокодифференцированных нейронах ядер шва активация 5-HT₁A-рецепторов приводит к снижению активности ERK, показано, что введение 5-HT₁A-агонистов в живой организм приводит к кратковременному увеличению активности ERK в дорсальной части ядер шва — эффект, полностью противоположный наблюдаемому в культуре тех же самых клеток. Кроме того, селективная активация 5-HT₁A-ауторецепторов ядер шва (а их там очень много) может также косвенно, опосредованно влиять на активность MEK/ERK-сигнального пути в мозгу через регуляцию высвобождения серотонина в других областях мозга. Таким образом, регуляция 5-HT₁A-рецепторами активности ERK в мозгу очень сложна и разнообразна и отличается в разных областях мозга и для разных типов нейронов. Для выяснения деталей специфической регуляции 5-HT1A-рецепторами активности ERK в разных областях мозга необходимы дальнейшие эксперименты с локальной активацией 5-HT₁A-рецепторов в тех или иных областях мозга и опыты с трансгенными мышами с пространственными и временными модификациями 5-HT₁A-рецепторов.
Учитывая выраженное влияние 5-HT₁A-рецепторов на регуляцию настроения, уровня тревожности, память и когнитивные функции, и наличие данных, показывающих выраженное регионарно-специфичное и типоспецифичное для разных типов нейронов влияние 5-HT₁A-рецепторов на активность ERK, роль ERK в опосредовании поведенческих эффектов 5-HT₁A-рецепторов и в регуляции активности нейронов заслуживает дальнейшего изучения, так же как и возможный терапевтический потенциал непосредственной фармакологической модуляции активности этого важного регулируемого 5-HT₁A-рецепторами сигнального каскада на более низких уровнях (на уровне тех или иных внутриклеточных звеньев, а не на уровне 5-HT₁A-рецептора).
Сигнальный путь PI3K/Akt
Серотониновые рецепторы подтипа 5-HT₁A также способны активировать другой сигнальный путь, обычно активируемый различными факторами роста, так называемый сигнальный путь PI3K / Akt. Когда тирозинкиназные рецепторы активируются факторами роста, они рекрутируют (активируют, фосфорилируя по тирозину) PI3K. Активированная PI3K, в свою очередь, активирует фосфоинозитид-зависимую киназу (PDK). А активированная PDK фосфорилирует и активирует Akt. Белок Akt хорошо известен как регулятор клеточного роста и выживаемости и как фактор, противодействующий апоптозу и способствующий митотической активности. Активация белка Akt ростовыми факторами опосредует вызываемый инсулином рост тканей и повышает устойчивость клеток к про-апоптотическим воздействиям. В центральной нервной системе белок Akt является важнейшим эффекторным белком в сигнальных каскадах, реализующих эффект нейротрофинов и различных нейромедиаторов. Также как и ERK, белок Akt является мультиспецифичной протеинкиназой широкого назначения, которая фосфорилирует множество мишеней-субстратов, в том числе протеинкиназы нисходящего эффекторного сигнального пути PI3K / Akt, различные факторы транскрипции белков и другие регуляторные белки.
Киназа гликоген-синтетазы 3 (GSK3) — это одна из протеинкиназ, которая в основном фосфорилируется и инактивируется белком Akt, но также может фосфорилироваться и инактивироваться несколькими другими протеинкиназами, такими, как протеинкиназа C или протеинкиназа А (цАМФ-зависимая). Киназа гликоген-синтетазы 3 (GSK3) является потенциальной молекулярной мишенью при некоторых психических заболеваниях, прежде всего заболеваниях аффективной сферы. В частности, известно, что нормотимик литий является селективным ингибитором GSK3. Ингибирование GSK3 фармакологическим методом или выведение лишённой GSK3 трансгенной мыши имитирует эффект антидепрессантов и антиманиакальных средств (нормотимиков и антипсихотиков). И наоборот, нарушенная регуляция GSK3 приводит к поведенческим аномалиям у животных, напоминающим состояния мании и депрессии.
Другая интересная группа субстратов фосфорилирования белком Akt — это так называемые транскрипционные факторы Forkhead-box-O (также называемые FoxOs). Активированный в результате действия факторов роста белок Akt фосфорилирует FoxOs и инактивирует их, вызывая их экспорт из ядра клетки в цитоплазму, где они не могут влиять на транскрипцию ДНК. И у беспозвоночных (в частности, насекомых, как фруктовая мушка дрозофила), и у высших животных белки FoxOs также могут подвергаться фосфорилированию и инактивации при воздействии серотонина через 5-HT₁A-рецептор и нисходящий эффекторный PI3K / Akt-зависимый механизм. В частности, показано, что подтип FoxO3a этого белка, распространённый в мозгу, может инактивироваться при воздействии имипрамина и подвергаться даунрегуляции (понижению активности и степени экспрессии) при воздействии ионов лития. Кроме того, трансгенные мыши с дефицитом FoxO проявляют поведение, характерное для мышей, получающих антидепрессанты и/или анксиолитики.
Таким образом, регуляция активности различных белков при помощи сигнального пути PI3K / Akt играет важную роль не только в росте и развитии нейронов и в их устойчивости к апоптозу, но и в поддержании активности нейронов и в регулировании поведенческих реакций, и этот сигнальный путь, по-видимому, тоже является важным для реализации антидепрессивного и анксиолитического эффектов 5-HT₁A-рецепторов.
В клетках, отличных от нейронов, активация гетерологичных 5-HT₁A-рецепторов закономерно повышает фосфорилирование белка Akt и его активность. Подобно регуляции белка ERK, регуляция Akt при помощи 5-HT₁A-рецепторов опосредуется Gi / Go и нисходящими эффекторными каскадами PI3K и Ras. Кроме того, активация Akt при активации 5-HT₁A-рецепторов предотвращается при повышении концентрации циклического АМФ в клетке (например вызванном одновременной активацией каких-то других, стимулирующих, а не ингибирующих аденилатциклазную активность, рецепторов) и соответствующей активации протеинкиназы A. Это влияние накопления цАМФ на активность Akt предотвращается или снимается ингибированием или инактивированием протеинкиназы А. Это заставляет предполагать, что вызываемое активацией 5-HT₁A-рецепторов ингибирование активности аденилатциклазы с соответствующим снижением содержания цАМФ в клетке и активности протеинкиназы A также является одним из механизмов, при помощи которых активация 5-HT₁A-рецепторов повышает активность Akt. Это согласуется с другими данными, которые показывают, что накопление цАМФ в клетке может вызывать дефосфорилирование и инактивацию Akt за счёт активации протеинкиназы A (PKA) и PKA-зависимой активации протеин-фосфатаз, в том числе протеин-фосфатазы 1.
Было показано, что агонисты 5-HT₁A-рецепторов закономерно повышают фосфорилирование и активность Akt в нейронах разного происхождения, включая происходящие из гиппокампа HN2-5 нейрональные клетки разной степени дифференцировки и зрелости, первичные низкодифференцированные гиппокампальные нейроны, первичные ромбоэнцефалические нейроны плода. Так же, как и в клетках не-нейронного происхождения, вызываемая активацией 5-HT₁A-рецепторов активация белка Akt является G-белок-зависимым процессом, чувствительным к ингибированию Gi и повышению активности аденилатциклазы коклюшным токсином и к концентрации цАМФ и опосредуемой ею активности протеинкиназы A. Этот процесс активации белка Akt под влиянием активации 5-HT₁A-рецепторов медиируется повышением активности PI3K, а также угнетением активности аденилатциклазы, снижением концентрации циклического АМФ и угнетением активности протеинкиназы А. Таким образом, опосредуемая отчасти PI3K, отчасти «каноническим» аденилатциклазным механизмом регуляция белка Akt и нижележащих звеньев сигнального каскада PI3K / Akt, вероятно, также является одним из важных механизмов, которыми опосредуются поведенческие и физиологические эффекты активации 5-HT₁A-рецепторов.
Прямые доказательства регуляции белка Akt активностью 5-HT₁A-рецепторов в мозге млекопитающих в нормальных физиологических условиях пока не получены. Однако существует немало косвенных доказательств участия 5-HT₁A-рецепторов в регуляции активности белка Akt и в целом сигнального каскада PI3K / Akt. Так, например, введение мыши 5-HT₁A-агониста 8-OH-DPAT значительно повышало фосфорилирование N-терминальных остатков серина у белка GSK3 — а белок GSK3 является одной из основных мишеней-субстратов белка Akt. Этот эффект наблюдался в том числе и в мозге мыши, в нескольких областях. Более того, было показано, что вызванное серотонином повышение фосфорилирования GSK3 опосредуется именно через 5-HT₁A-рецепторы (не через другие типы серотониновых рецепторов) и блокируется антагонистами 5-HT₁A-рецепторов. Кроме того, было показано, что в мозге мыши повышение уровня серотонина (при введении антидепрессантов, лития, L-триптофана) приводит к повышению фосфорилирования и активности белка Akt — эффект, который предотвращался введением в желудочки мозга ингибитора PI3K. Однако ни одно из этих исследований не предоставляет прямых доказательств того, что 5-HT₁A-рецепторы регулируют активность белка Akt в нормальных физиологических условиях в мозге млекопитающих. Также неизвестно, является ли влияние 5-HT₁A-рецепторов на активность белка Akt регионарно-специфичным и/или типоспецифичным для типа нейронов, как это имеет место быть с белками ERK. Дополнительные исследования также необходимы и для того, чтобы окончательно определить, какую роль играет регуляция PI3K / Akt сигнального пути 5-HT₁A-рецепторами в общей картине действия 5-HT₁A-рецепторов на нейрональную активность и на поведение животных.
Взятые вместе, все эти данные свидетельствуют о том, что 5-HT₁A-рецепторы сопряжены с активацией не только канонического Gi / Go-опосредованного сигнального каскада, включающего аденилатциклазу, цАМФ и протеинкиназу A, а также G-белок-зависимые каналы ионов калия (GIRK), но и с активацией добавочных сигнальных каскадов, обычно ассоциируемых с факторами роста клеток и с модуляцией нейрональной пластичности, роста и развития клеток и с их устойчивостью к апоптозу — в частности, с активацией ERK-сигнального пути и сигнального пути PI3K / Akt. Для детального изучения того, каким образом работа этих сигнальных каскадов зависит от типа клеток и от регионарной специфичности в разных областях мозга, и каким образом они медиируют разнообразные физиологические и поведенческие аспекты действия 5-HT₁A-рецепторов, требуются дополнительные исследования. Лучшее понимание механизмов передачи сигнала от 5-HT₁A-рецептора внутрь клетки может способствовать обнаружению новых мишеней для лекарств, и в конечном итоге — разработке новых лекарств (в частности, анксиолитиков и антидепрессантов) для лечения патологических состояний, ассоциирующихся с аномальным функционированием 5-HT₁A-рецепторов или их внутриклеточных каскадов (а это такие состояния, как тревожность, депрессия).
Физиологическая роль
Эффекты 5-HT1A-рецептора в центральной нервной системе
Рецепторы подтипа 5-HT₁A вовлечены в нейромодуляцию (модуляцию активности центральной нервной системы). В частности, активация этих рецепторов агонистами приводит к снижению артериального давления и урежению частоты сердечных сокращений благодаря центральным механизмам действия, а также благодаря индуцированию периферической вазодилатации и за счёт стимуляции активности блуждающего нерва.[48] Эти эффекты являются результатом активации 5-HT₁A-рецепторов, находящихся в сосудодвигательном (прессорном) центре продолговатого мозга, в его рострально-вентролатеральной части.[48] Симпатолитическое гипотензивное лекарство урапидил является не только антагонистом α₁-адренорецепторов и агонистом α₂-адренорецепторов, но и агонистом 5-HT₁A-рецепторов. И было показано, что его свойства как агониста 5-HT₁A-рецепторов привносят свой вклад в его гипотензивный эффект.[49][50] Вазодилатация поверхностных сосудов кожи через активацию центральных 5-HT₁A-рецепторов увеличивает теплоотдачу (отдачу тепла) с поверхности тела и тем самым способствует снижению температуры тела.[51][52]
Активация центральных 5-HT₁A-рецепторов вызывает либо усиление, либо ослабление выделения норадреналина в голубоватом пятне, в зависимости от вида животного — у человека и других приматов ослабление, у крыс и других грызунов — усиление. А нисходящие норадренергические влияния из голубоватого пятна модулируют активность постсинаптических α₂-адренергических рецепторов в глазодвигательных ядрах Эдингера-Вестфаля, а те, в свою очередь, через симпатические волокна глазодвигательных нервов передают команды цилиарным мышцам глаз. Таким образом, стимуляция центральных 5-HT₁A-рецепторов у человека и других приматов приводит к ослаблению норадренергической нейротрансмиссии в глазодвигательных ядрах Эдингера-Вестфаля и к сужению зрачка (миозу), а у крыс и других грызунов — к усилению норадренергической нейротрансмиссии в ядрах Эдингера-Вестфаля и к расширению зрачков (мидриазу).[53][54][55]
Агонисты 5-HT₁A-рецепторов, такие, как буспирон[56] и флезиноксан[57] проявляют эффективность в уменьшении симптомов тревоги[58] и депрессии[59]. Буспирон и тандоспирон разрешены к применению для этих целей в разных странах мира. Другие агонисты 5-HT₁A-рецепторов, такие, как гепирон[60], флезиноксан[61], флибансерин[62], или налузотан[63] находятся на различных стадиях разработки и исследования (либо разрабатывались и исследовались в прошлом, но разработка была по тем или иным причинам прекращена), однако на данный момент ни один из них ещё не закончил полный цикл разработки и исследований и не получил разрешения на клиническое применение у человека. Некоторые из атипичных антипсихотиков, такие, как арипипразол[64] также являются парциальными агонистами 5-HT₁A-рецепторов и иногда используются в малых дозах в качестве средств для аугментации (усиления действия, потенцирования) стандартных антидепрессантов, в частности СИОЗС, ТЦА[65].
Десенситизация 5-HT₁A- пресинаптических ауторецепторов и усиление активности постсинаптических 5-HT₁A-рецепторов в результате вызванного теми или иными механизмами повышения общего содержания серотонина в синапсах мозга (будь то повышение биосинтеза серотонина под влиянием препаратов лития или под влиянием усиленного поступления в организм прекурсоров серотонина — триптофана или 5-гидрокситриптофана, или блокада обратного захвата серотонина при применении СИОЗС, ТЦА, или блокада разрушения серотонина ферментом моноаминоксидазой при применении ИМАО, или блокада пресинаптических «тормозных» α₂-гетероадренорецепторов на серотониновых нейронах при применении миртазапина и др.) — считается одним из важных механизмов реализации антидепрессивного действия всех вышеперечисленных соединений, как лекарств, так и пищевых добавок.[66] В дополнение к этому, некоторые антидепрессанты, такие, как тразодон, нефазодон, вилазодон, вортиоксетин — обладают прямым и непосредственным 5-HT₁A-агонистическим действием, что, вероятно, также вносит свой вклад в их антидепрессивную активность, наряду с блокадой обратного захвата моноаминов и другими эффектами.
Также считается, что активация 5-HT₁A-рецепторов и сопряжённое с ней усиление секреции окситоцина, по всей вероятности, играет важную роль в реализации «просоциального» (социализирующего), быстрого антидепрессивного, анксиолитического и эйфоризирующего эффекта агентов, вызывающих выброс серотонина, таких, как МДМА.[67][68]
Рецепторы подтипа 5-HT₁A в дорсальной части ядер шва ко-локализованы (находятся рядом на одних и тех же клетках) с NK₁-рецепторами (рецепторами к нейрокинину-1, также называемому субстанцией P). Было показано, что 5-HT₁A-рецепторы способны угнетать выделение NK₁ из пептидергических нейронов, а также понижать чувствительность NK₁-рецепторов к их эндогенному лиганду. А уменьшение опосредованной NK₁ нейротрансмиссии, в свою очередь, вторично уменьшает норадренергическую нейротрансмиссию в соответствующих областях мозга.[69][70] А опосредованная NK₁ нейротрансмиссия принимает участие в регуляции тошноты и рвоты, болевой чувствительности, а также возбуждения и тревоги. В результате, в дополнение к антидепрессивному и анксиолитическому эффектам, активация 5-HT₁A-рецепторов при помощи агонистов приводит также к противорвотному эффекту[71][72] и анальгетическому эффекту,[73][74]. Все или часть этих эффектов могут полностью или частично зависеть от угнетения активированными 5-HT₁A-рецепторами опосредованной NK₁ нейропередачи. И наоборот, новейшие NK₁-антагонисты, которые в основном используются в настоящее время как противорвотные средства для лечения тошноты и рвоты при лучевой терапии и химиотерапии злокачественных опухолей, в последнее время исследуются также как потенциальные новые агенты для лечения тревоги и депрессии, а также болевых синдромов.[75]
Показано, что активация 5-HT₁A-рецепторов способна повышать выделение дофамина в медиальной префронтальной коре, стриатуме и гиппокампе. Эти эффекты могут оказаться полезными для улучшения состояния больных с шизофренией и болезнью Паркинсона.[76][77] Как уже упоминалось выше, некоторые из атипичных антипсихотиков являются парциальными агонистами 5-HT₁A-рецепторов. Было показано, что это свойство повышает их клиническую эффективность при шизофрении, особенно в отношении негативной, когнитивной и депрессивной симптоматики, и повышает их «атипичность» (уменьшает вероятность экстрапирамидных побочных явлений).[76][78][79]
Усиление дофаминергической нейротрансмиссии в этих областях мозга может также играть важную роль в реализации антидепрессивного и анксиолитического эффектов, наблюдаемых при активации постсинаптических 5-HT₁A-рецепторов (напрямую, как при применении агонистов 5-HT₁A-рецепторов, или косвенно через повышение уровня серотонина, как при применении антидепрессантов, лития, прекурсоров серотонина).[80][81]
Вместе с тем сверхстимуляция 5-HT₁A-рецепторов, возникающая при приёме серотонинергических препаратов, ответственна за возникновение редкого, но опасного побочного эффекта — серотонинового синдрома (в частности, таких его проявлений, как гиперактивность, гиперрефлексия и беспокойство)[82].
Было показано, что активация 5-HT₁A-рецепторов может нарушать некоторые функции памяти (это касается как декларативной, так и не-декларативной памяти) и способность к обучению (вследствие нарушения работы механизмов кодирования и хранения новой информации в памяти), за счёт ингибирования высвобождения глутамата и ацетилхолина в различных областях мозга. Возможно, что этот эффект лежит в основе иногда наблюдаемого неблагоприятного влияния антидепрессантов на память.[83] В то же время известно, что активация 5-HT₁A-рецепторов способна улучшать когнитивные функции, ассоциирующиеся с префронтальной корой (и нарушенные при шизофрении и при депрессиях), вероятно, вследствие повышения содержания в префронтальной коре дофамина и ацетилхолина.[84] И напротив, было показано, что антагонисты 5-HT₁A-рецепторов, такие, как лекозотан, способны улучшать определённые аспекты обучаемости и памяти, по крайней мере в опытах на крысах, и, как следствие, в настоящее время изучаются в качестве потенциальных кандидатов на лекарства для лечения болезни Альцгеймера.[85]
Другие наблюдавшиеся в научных исследованиях результаты активации 5-HT₁A-рецепторов включают в себя:
- Уменьшение агрессивности[86][87]
- Повышение социальности и социальной активности[88]
- Уменьшение импульсивности[89]
- Уменьшение влечения к алкоголю и психоактивным веществам[90][91][92]
- Повышение либидо и сексуальной возбудимости[93][94]
- Угнетение эректильной функции[95][96]
- Снижение аппетита[97]
- Пролонгирование латентной фазы до наступления REM-фазы сна[98][99]
- Уменьшение вызываемой опиоидами депрессии дыхания.[100]
Эндокринные эффекты 5-HT1A-рецептора
Активация 5-HT₁A-рецепторов вызывает секрецию различных гормонов, включая кортизол, АКТГ, окситоцин, пролактин, соматотропин, β-эндорфин.[101][102][103][104] В отличие от 5-HT2-рецепторов, активация 5-HT₁A-рецепторов не оказывает влияния на секрецию вазопрессина или ренина.[101][102] Предполагается, что секреция окситоцина может привносить свой вклад в просоциальные, антиагрессивные, анксиолитические эффекты, наблюдаемые при активации 5-HT₁A-рецепторов.[68] Секреция β-эндорфина может привносить свой вклад в антидепрессивный, анксиолитический и анальгетический эффекты.[105]
Ауторецепторы и гетерорецепторы
Рецепторы (в том числе рецепторы подтипа 5-HT₁A) могут находиться на теле клетки, её дендритах, аксонах, а также могут находиться как на пресинаптической, так и на постсинаптической сторонах нервных окончаний и синапсов. Рецепторы, расположенные на теле, аксонах и дендритах, называются соматодендритными. Те, которые расположены на пресинаптической стороне синаптической щели, называют пресинаптическими, а те, которые расположены на постсинаптической стороне — соответственно, постсинаптическими. Рецепторы на пресинаптической стороне, которые чувствительны к тому же самому нейромедиатору, который высвобождается данным нейроном в данную синаптическую щель, называют [пресинаптическими] [тормозными], ауторегуляторными рецепторами (или просто [пресинаптическими] [тормозными] ауторецепторами). Эти пресинаптические ауторецепторы играют роль ключевого компонента в ультракороткой петле отрицательной обратной связи: выделение нейроном того или иного нейромедиатора стимулирует «тормозные» ауторецепторы на нём самом, вследствие чего ингибируется, предотвращается дальнейшее выделение этого нейромедиатора в синапс и/или активируется его обратный захват. Таким образом, стимуляция 5-HT₁A пресинаптических ауторецепторов ингибирует высвобождение серотонина в нервных окончаниях и синапсах. Ситуация осложняется ещё и тем, что пресинаптические (регуляторные) ауторецепторы чувствительнее постсинаптических (эффекторных) рецепторов, к тому же их на пресинаптической стороне больше (их плотность выше), поэтому для их активации достаточно меньшей концентрации серотонина (или другого агониста) в синапсе. Это эффективно ограничивает сверху максимально возможный выброс серотонина пресинаптической клеткой и максимально возможную серотонинергическую стимуляцию постсинаптической клетки в каждом конкретном случае. По этой причине агонисты 5-HT₁A-рецепторов имеют тенденцию к двухфазности кривой «доза-эффект». В малых дозах они преимущественно стимулируют более чувствительные и более обильно представленные пресинаптические 5-HT₁A-ауторецепторы, тем самым уменьшая высвобождение серотонина в синапс и в конечном итоге уменьшая серотонинергическую стимуляцию постсинаптических клеток и в целом серотонинергическую нейропередачу. А в более высоких дозах они ещё сильнее уменьшают выброс эндогенного серотонина (за счёт ещё более сильной активации пресинаптических ауторецепторов, а также за счёт включения других, менее чувствительных и более «длинных» петель обратной связи), но одновременно с этим усиливают активность постсинаптических 5-HT₁A-рецепторов, непосредственно стимулируя их в этих (высоких) концентрациях вместо эндогенного агониста — серотонина. Аналогичную двухфазность проявляют и некоторые антагонисты 5-HT₁A-рецепторов, например пиндолол — в малых дозах они преимущественно блокируют пресинаптические 5-HT₁A тормозные ауторецепторы, усиливая тем самым выброс серотонина в синапс и увеличивая серотонинергическую нейропередачу, а в больших дозах начинают блокировать также и постсинаптические 5-HT₁A-рецепторы, препятствуя действию эндогенного нейромедиатора серотонина и снижая серотонинергическую передачу.
Это опосредованное пресинаптическими тормозными ауторецепторами угнетение высвобождения серотонина в синапс постулировано в качестве одной из главных причин наличия характерной 2-3-недельной задержки проявления терапевтического действия серотонинергических антидепрессантов, таких, как СИОЗС, ТЦА, ИМАО.[106] Пресинаптические ауторецепторы должны сначала в достаточной степени десенситизироваться (подвергнуться даунрегуляции), прежде чем концентрация серотонина в синапсах, во внеклеточном пространстве, сможет сколько-нибудь существенно вырасти под влиянием антидепрессантов.[106][107] И хотя чувствительность пресинаптических ауторецепторов несколько уменьшается при длительном, хроническом лечении антидепрессантами, они всё равно остаются весьма эффективными ограничителями, предотвращающими значительное повышение концентрации серотонина в синапсах.[106] По этой причине современные исследователи изыскивают антидепрессанты, которые бы сочетали свойства ингибиторов обратного захвата серотонина и антагонистов или парциальных агонистов пресинаптических 5-HT₁A-ауторецепторов (чтобы «выключать» этот мешающий механизм отрицательной обратной связи) и/или агонистов постсинаптических 5-HT₁A-рецепторов (чтобы эффективно обходить этот механизм, непосредственно стимулируя нужные рецепторы). Предполагается, что препараты, сочетающие такие свойства, могут обладать преимуществом в эффективности действия и/или в скорости наступления эффекта, или в частоте некоторых побочных эффектов (например, сексуальных) по сравнению с уже имеющимися антидепрессантами. Примерами таких препаратов являются вилазодон и вортиоксетин.[108]
В отличие от большинства антидепрессантов, которые повышают уровень серотонина в синапсах только при достаточно длительном применении, а в начале терапии его даже снижают из-за срабатывания ауторегуляторных механизмов, серотонин-высвобождающие агенты (СВА), такие, как МДМА и фенфлюрамин, способны обходить серотониновые ауторегуляторные механизмы, такие, как 5-HT₁A-ауторецепторы. Они делают это благодаря прямому действию на механизмы высвобождения серотонина в нейронах, и вызывают выброс серотонина вне зависимости от ингибирования ауторецепторами.[109] Вследствие этого, серотонин-высвобождающие агенты вызывают немедленное и значительно большее, чем при применении антидепрессантов, повышение уровня серотонина в синапсах. По контрасту с СВА, антидепрессанты, будь то СИОЗС, ИМАО или ТЦА, на самом деле в начале терапии обычно понижают уровень серотонина в синапсах из-за включения ауторегуляторных механизмов, и требуют нескольких недель терапии для того, чтобы уровень серотонина начал повышаться, и ещё нескольких недель для того, чтобы уровень серотонина достиг максимально возможного при данной дозе антидепрессанта «плато», и клинический эффект в отношении таких состояний, как депрессия или тревога, проявился в полной мере.[110][111] По этой причине так называемые «селективные серотонин-высвобождающие агенты» (ССВА), такие, как MDAI и MMAI (модифицированные аналоги или производные МДМА) предложены в качестве возможных новых кандидатов в антидепрессивные лекарства с, вероятно, более быстрым наступлением эффекта и большей клинической эффективностью по сравнению с существующими антидепрессантами.[110]
Достаточно высокие дозы агонистов 5-HT1A-рецепторов также могут эффективно обходить вызываемое пресинаптическими 5-HT₁A-ауторецепторами ингибирование высвобождения серотонина и увеличивать серотонинергическую нейропередачу, непосредственно стимулируя постсинаптические (эффекторные) 5-HT₁A-рецепторы, вместо серотонина. Однако, по контрасту с СВА, агонисты 5-HT₁A-рецепторов не в состоянии обойти ингибирующие эффекты пресинаптических 5-HT₁A-рецепторов, расположенных в качестве гетерорегуляторных рецепторов (гетерорецепторов) в не-серотонинергических синапсах (например, в норадренергических, дофаминергических или глутаматергических), в которых постсинаптических 5-HT₁A-рецепторов нет. В этих синапсах в норме серотонин ингибирует через 5-HT₁A гетерорецепторы высвобождение других медиаторов, таких, как норадреналин, дофамин или глутамат. Агонисты 5-HT₁A-рецепторов делают то же самое вместо серотонина. Клиническое значение этого гетеро-ингибирования неизвестно, однако предполагается, что оно должно ограничивать клиническую эффективность «чистых» 5-HT₁A-агонистов и их применимость в реальной практике (а не в экспериментах на животных) в качестве антидепрессантов. Вероятно, для того, чтобы быть эффективным антидепрессантом, препарат должен, помимо 5-HT₁A-агонистической активности, обладать ещё и другими полезными для антидепрессанта свойствами (например, ингибировать обратный захват моноаминов или быть агонистом либо антагонистом ещё каких-то подтипов серотониновых рецепторов, как вилазодон или вортиоксетин). Вместе с тем, относительно «чистые» 5-HT₁A-агонисты (такие, как буспирон) или средства, обладающие среди прочих свойств 5-HT₁A-агонистической активностью (ряд атипичных антипсихотиков, особенно арипипразол, а также йохимбин, лизурид) могут быть эффективны в качестве адъюванта (дополнения) к традиционным антидепрессантам, для усиления (потенцирования) их действия и/или ускорения наступления эффекта, то есть в качестве потенцирующих агентов (аугментаторов).
Генетика
Белок рецептора подтипа 5-HT₁A кодируется геном HTR1A. У человека обнаружены несколько вариантов полиморфизма этого гена. Исследование 2007 года перечисляет обнаруженные к тому времени 27 однонуклеотидных полиморфизмов гена HTR1A.[112] Наиболее изученные на сегодня однонуклеотидные полиморфизмы — это C-1019G (rs6295), C-1018G,[113] Ile28Val (rs1799921), Arg219Leu (rs1800044) и Gly22Ser (rs1799920).[112] Некоторые другие изученные однонуклеотидные полиморфизмы — это Pro16Leu, Gly272Asp и синонимичный полиморфизм G294A (rs6294). Эти генетические полиморфизмы подвергались изучению на предмет их возможной связи с психическими заболеваниями, однако чётких и однозначно трактуемых результатов при этом не было получено.[112]
Взаимодействия рецептора с другими белками
Рецепторы подтипа 5-HT₁A взаимодействуют с BDNF (мозговым нейротрофическим фактором), что может играть существенную роль в их способности регулировать настроение, эмоциональное состояние и уровень тревожности, а также память и когнитивные функции.[114][115]
Лиганды
Распределение 5-HT₁A-рецепторов в мозге человека может быть визуализировано при помощи позитронно-эмиссионной томографии с меченым радиоактивным изотопом углерода — углеродом-11 (11C) радиолигандом WAY-100,635.[116]
В частности, в одном из исследований было обнаружено повышенное связывание меченого 11C радиолиганда при ожирении и сахарном диабете 2-го типа. Учитывая, что такое же повышение количества 5-HT₁A-рецепторов (их сенситизация или апрегуляция) обнаруживается при депрессиях и тревожных состояниях, эта находка, возможно, может отчасти объяснять повышенную частоту депрессий и тревожных состояний у больных ожирением или сахарным диабетом 2-го типа, а также повышенную предрасположенность депрессивных и тревожных больных к развитию ожирения и СД 2-го типа. Возможно, в этих состояниях играют роль общие клеточные механизмы.[117] В другом ПЭТ-исследовании была обнаружена отрицательная корреляция между количеством 5-HT₁A-рецепторов в ядрах шва, гиппокампе и коре больших полушарий и сведениями, которые предоставляли добровольцы относительно наличия у них внетелесного или иного «духовного» опыта.[118] Меченый тритием (радиоактивным изотопом водорода — 3H) этот же лиганд WAY-100,635 может также быть использован в ауторадиографическом методе. Показано повышенное количество 5-HT₁A-рецепторов во фронтальной коре больных шизофренией.[119]
Агонисты
Полные
Парциальные
Антидепрессанты
- Тразодон
- Нефазодон
- Вилазодон
- Вортиоксетин
Небензодиазепиновые анксиолитики
- Биноспирон
- Буспирон
- Гепирон
- Залоспирон
- Каннабидиол
- Ипсапирон
- Пероспирон
- Тандоспирон
- Тиоспирон
Атипичные антипсихотики
- Арипипразол
- Азенапин
- Зипрасидон
- Кветиапин
- Клозапин
- Луразидон
- Оланзапин
Типичные антипсихотики
Гипотензивные препараты
- Урапидил
- Раувольсцин
Антимигренозные препараты
Галлюциногены
Эмпатогены
Дофаминергические антипаркинсонические препараты
- Лизурид
Препараты, улучшающие либидо и сексуальную функцию
Разные и исследовательские препараты
- 5-карбоксамидотриптамин (5-CT)
- 5-метокситриптамин (5-MT)
- 5-MeO-DMT
- Адатансерин
- Флибансерин
- Альфа-этилтриптамин (αET)
- Альфа-метилтриптамин (αMT)
- Bay R 1531
- Бефирадол
- Буфотенин
- Элтопразин
- Этоперидон
- F-11,461
- F-12,826
- F-13,714
- F-14,679
- Флезиноксан
- Гинкго Билоба[141]
- LY-301,317
- Эбалзотан
- Налузотан
- Пиклозотан
- Саризотан
- NBUMP
- RU-24,969
- S-15,535
- SSR-181,507
- Сунепитрон
- Трифторметилфенилпиперазин
- Ксалипроден
- Лимонен[142]
Антагонисты
β-адреноблокаторы
Антимигренозные препараты
Атипичные антипсихотики
Разные и исследовательские препараты
Примечания
- T. Conrad Gilliam, Nelson B. Freimer, Charles A. Kaufmann, Peter P. Powchik, Anne S. Bassett, Ulla Bengtsson, John J. Wasmuth. Deletion Mapping of DNA Markers to a Region of Chromosome 5 That Cosegregates with Schizophrenia (англ.) // PubMed Central Genomics. — Academic Press, Nov 1989. — Iss. 5(4). — P. 940—944. — doi:10.1016/0888-7543(89)90138-9. — PMID 2591972.
- HTR1A 5-hydroxytryptamine (serotonin) receptor 1A, G protein-coupled [ Homo sapiens (human) ]. Дата обращения: 24 февраля 2015.
- Paila YD1, Tiwari S, Sengupta D, Chattopadhyay A. Molecular modeling of the human serotonin1Areceptor: role of membrane cholesterol in ligand binding of the receptor (англ.) // Molecular BioSystems. — 2011. — Vol. 7, iss. 7(1), no. 1. — P. 224—234. — doi:10.1039/C0MB00148A. — PMID 20967314.
- Md. Jafurulla, A. Chattopadhyay. Membrane Lipids in the Function of Serotonin and Adrenergic Receptors (англ.) // Current Medicinal Chemistry. — 2013. — Vol. 20, iss. 20(1), no. 1. — P. 47—55. — doi:10.2174/0929867311302010006. — PMID 23151002.
- Ekaterina Papoucheva, Aline Dumuis, Michèle Sebben, Diethelm W. Richter, Evgeni G. Ponimaskin. The 5-Hydroxytryptamine(1A) Receptor Is Stably Palmitoylated, and Acylation Is Critical for Communication of Receptor with Gi Protein (англ.) // The Journal of Biological Chemistry. — 30 Jan 2004. — Vol. 279, iss. 279(5), no. 5. — doi:10.1074/jbc.M308177200. — PMID 14604995.
- Ute Renner, Konstantin Glebov, Thorsten Lang, Ekaterina Papusheva, Saju Balakrishnan, Bernhard Keller, Diethelm W. Richter, Reinhard Jahn, Evgeni Ponimaskin. Localization of the Mouse 5-Hydroxytryptamine1A Receptor in Lipid Microdomains Depends on Its Palmitoylation and Is Involved in Receptor-Mediated Signaling (англ.) // Molecular Pharmacology. — May 31, 2007. — Vol. 72, iss. 72(3), no. 3. — P. 502—513. — doi:10.1124/mol.107.037085. — PMID 17540717.
- Damien Carrel, Michel Hamon, Michèle Darmon. Role of the C-terminal di-leucine motif of 5-HT1A and 5-HT1B serotonin receptors in plasma membrane targeting (англ.) // Journal of Cell Science. — The Company of Biologists, September 26, 2006. — Vol. 119, iss. 119(20), no. 20. — P. 4276—4284. — PMID 17003106.
- Anthony TE, Azmitia EC. Molecular characterization of antipeptide antibodies against the 5-HT1A receptor: evidence for state-dependent antibody binding (англ.) // Molecular Brain Research. — 15 Oct 1997. — Vol. 50, iss. 50(1—2), no. 1—2. — P. 277—284. — PMID 9406944.
- Paola M. C. Lembo, Mohammad H. Ghahremani, Stephen J. Morris, Paul R. Albert. A Conserved Threonine Residue in the Second Intracellular Loop of the 5-Hydroxytryptamine 1A Receptor Directs Signaling Specificity (англ.) // Molecular Pharmacology. — July 1, 1997. — Vol. 52, iss. 52(1), no. 1. — P. 164—171. — PMID 9224826.
- Nataliya Gorinski, Noga Kowalsman, Ute Renner, Alexander Wirth, Michael T. Reinartz, Roland Seifert, Andre Zeug, Evgeni Ponimaskin, Masha Y. Niv. Computational and Experimental Analysis of the Transmembrane Domain 4/5 Dimerization Interface of the Serotonin 5-HT1A Receptor (англ.) // Molecular Pharmacology. — June 5, 2012. — Vol. 82, iss. 82(3), no. 3. — P. 448—463. — doi:10.1124/mol.112.079137. — PMID 22669805.
- Ute Renner, Andre Zeug, Andrew Woehler, Marcus Niebert, Alexander Dityatev, Galina Dityateva, Nataliya Gorinski, Daria Guseva, Dalia Abdel-Galil, Matthias Fröhlich, Frank Döring, Erhard Wischmeyer, Diethelm W. Richter, Erwin Neher, Evgeni G. Ponimaskin. Heterodimerization of serotonin receptors 5-HT1A and 5-HT7 differentially regulates receptor signalling and trafficking (англ.) // Journal of Cell Science. — The Company of Biologists, February 22, 2012. — Vol. 125, iss. 125, no. Pt(10). — P. 2486—2499. — doi:10.1242/jcs.101337. — PMID 22357950.
- Kamran Salim, Tim Fenton, Jamil Bacha, Hector Urien-Rodriguez, Tim Bonnert, Heather A. Skynner, Emma Watts, Julie Kerby, Anne Heald, Margaret Beer, George McAllister, Paul C. Guest. Oligomerization of G-protein-coupled Receptors Shown by Selective Co-immunoprecipitation (англ.) // The Journal of Biological Chemistry. — February 19, 2002. — Vol. 277, iss. 277(18), no. 18. — P. 15482—15485. — doi:10.1074/jbc.M201539200. — PMID 11854302.
- Hiroshi Ito, Christer Halldin, Lars Farde. Localization of 5-HT1A Receptors in the Living Human Brain Using Carbonyl-11C-WAY-100635: PET with Anatomic Standardization Technique (англ.) // Journal of Nuclear Medicine. — Jan 1999. — Vol. 40, iss. 40(1), no. 1. — P. 102—109. — PMID 9935065.
- Richard A. Glennon; Malgorzata Dukat, Richard B. Westkaemper. Serotonin Receptor Subtypes and Ligands (англ.). Psychopharmacology - 4th Generation of Progress. American College of Neurophyscopharmacology (1 января 2000). Дата обращения: 24 февраля 2015. Архивировано 21 апреля 2008 года.
- Julián de Almeida, Guadalupe Mengod. Serotonin 1A receptors in human and monkey prefrontal cortex are mainly expressed in pyramidal neurons and in a GABAergic interneuron subpopulation: implications for schizophrenia and its treatment (англ.) // Journal of Neurochemistry. — 29 Aug 2008. — Vol. 107, iss. 107(2), no. 2. — P. 488—496. — doi:10.1111/j.1471-4159.2008.05649.x. — PMID 18761712.
- De Vivo M, Maayani S. Characterization of the 5-hydroxytryptamine1a receptor-mediated inhibition of forskolin-stimulated adenylate cyclase activity in guinea pig and rat hippocampal membranes (англ.) // J Pharmacol Exp Ther.. — Jul 1986. — Vol. 238, iss. 238(1), no. 1. — P. 248—253. — PMID 2941565.
- Lutz Birnbaumer. Expansion of signal transduction by G proteins The second 15 years or so: From 3 to 16 α subunits plus βγ dimers (англ.) // PubMed Central Biochim Biophys Acta. — PubMed Central, Apr 2007. — Vol. 1768, iss. 1768(4), no. 4. — P. 772—793. — doi:10.1016/j.bbamem.2006.12.002. — PMID 17258171.
- P. Schoeffter, D. Hoyer. Centrally acting hypotensive agents with affinity for 5-HT1A binding sites inhibit forskolin-stimulated adenylate cyclase activity in calf hippocampus (англ.) // PubMed Central British Journal of Pharmacology. — PubMed Central, Nov 1988. — Vol. 95, iss. 95(3), no. 3. — P. 975—985. — PMID 3207999.
- Dumuis A1, Sebben M, Bockaert J. Pharmacology of 5-hydroxytryptamine-1A receptors which inhibit cAMP production in hippocampal and cortical neurons in primary culture (англ.) // Molecular Pharmacology. — Feb 1988. — Iss. 33(2). — P. 178—186. — PMID 2828913.
- Sprouse JS1, Aghajanian GK. Responses of hippocampal pyramidal cells to putative serotonin 5-HT1A and 5-HT1B agonists: a comparative study with dorsal raphe neurons (англ.) // Neuropharmacology. — Jul 1988. — Vol. 27, iss. 27(7), no. 7. — P. 707—715. — PMID 2901680.
- Harrington MA, Shaw K, Zhong P, Ciaranello RD. Agonist-induced desensitization and loss of high-affinity binding sites of stably expressed human 5-HT1A receptors (англ.) // Journal of Pharmacology and Experimental Therapeutics.. — Mar 1994. — Vol. 268, iss. 268(3), no. 3. — P. 1098—1106. — PMID 8138923.
- John R. Raymond, Frank J. Albers, John P. Middleton, Robert J. Lefkowitz, Marc G. Caron, Lina M. Obeid, Vincent W. Dennis. 5-HT1A and Histamine H1 Receptors in HeLa Cells Stimulate Phosphoinositide Hydrolysis and Phosphate Uptake via Distinct G Protein Pools (англ.) // The Journal of Biological Chemistry. — 5 Jan 1991. — Vol. 266, iss. 266(1), no. 1. — P. 372—379. — PMID 1845968.
- Cell-specific Signaling of the 5-HT1A Receptor
- Cell-specific physical and functional coupling of human 5-HT1A receptors to inhibitory G protein alpha-subunits and lack of coupling to Gs alpha.
- Lack of 5-hydroxytryptamine1A-mediated inhibition of adenylyl cyclase in dorsal raphe of male and female rats.
- Regional Differences in the Coupling of 5-Hydroxytryptamine-1A Receptors to G Proteins in the Rat Brain
- Functional Consequences of 5-HT Transporter Gene Disruption on 5-HT1A Receptor-Mediated Regulation of Dorsal Raphe and Hippocampal Cell Activity
- Differential adaptation of brain 5-HT1A and 5-HT1B receptors and 5-HT transporter in rats treated chronically with fluoxetine.
- In vivo electrophysiological evidence for the regulatory role of autoreceptors on serotonergic terminals.
- Role of Hippocampal CaMKII in Serotonin 5-HT1A Receptor-Mediated Learning Deficit in Rats
- G Protein-Coupled Inwardly Rectifying K+ Channels (GIRKs) Mediate Postsynaptic but Not Presynaptic Transmitter Actions in Hippocampal Neurons
- Pharmacologically distinct actions of serotonin on single pyramidal neurones of the rat hippocampus recorded in vitro.
- G alpha o1 decapeptide modulates the hippocampal 5-HT1A potassium current.
- Differential modulation of three separate K-conductances in hippocampal CA1 neurons by serotonin.
- Serotonin decreases population spike amplitude in hippocampal cells through a pertussis toxin substrate.
- Gi Irks GIRKs.
- Suppression of conditioning to ambiguous cues by pharmacogenetic inhibition of the dentate gyrus
- 5-hydroxytryptamine1A receptor-mediated effects on adenylate cyclase and nitric oxide synthase activities in rat ventral prostate.
- Effects of SSRIs on sexual function: a critical review.
- Effects of Vasodilatory β-Adrenoceptor Antagonists on Endothelium-Derived Nitric Oxide Release in Rat Kidney
- Photic entrainment of circadian rhythms in rodents.
- Effects of a nitric oxide synthase inhibitor on 5-HT1A receptor agonist 8-OH-DPAT-induced hyperphagia in rats.
- Activation of serotonergic 5-HT1A receptor reduces Ca(2+)- and glutamatergic receptor-evoked arachidonic acid and No/cGMP release in adult hippocampus.
- Possible role of nitric oxide in 5-hydroxytryptamine-induced increase in vascular permeability in mouse skin.
- 5-Hydroxytryptamine-induced vasodilatation in the isolated perfused rat kidney: are endothelial 5-HT1A receptors involved?
- The nitric oxide synthesis/pathway mediates the inhibitory serotoninergic responses of the pressor effect elicited by sympathetic stimulation in diabetic pithed rats.
- Cell Survival Promoted by the Ras-MAPK Signaling Pathway by Transcription-Dependent and -Independent Mechanisms
- Dabiré H. Central 5-hydroxytryptamine (5-HT) receptors in blood pressure regulation (англ.) // Therapie : journal. — 1991. — Vol. 46, no. 6. — P. 421—429. — PMID 1819150.
- Ramage A.G. The mechanism of the sympathoinhibitory action of urapidil: role of 5-HT₁A receptors (англ.) // Br. J. Pharmacol. : journal. — 1991. — April (vol. 102, no. 4). — P. 998—1002. — doi:10.1111/j.1476-5381.1991.tb12290.x. — PMID 1855130.
- Kolassa N., Beller K.D., Sanders K.H. Involvement of brain 5-HT₁A receptors in the hypotensive response to urapidil (англ.) // Am. J. Cardiol. : journal. — 1989. — Vol. 64, no. 7. — P. 7D—10D. — doi:10.1016/0002-9149(89)90688-7. — PMID 2569265.
- Ootsuka Y., Blessing W.W. Activation of 5-HT₁A receptors in rostral medullary raphé inhibits cutaneous vasoconstriction elicited by cold exposure in rabbits (англ.) // Brain Res. : journal. — 2006. — Vol. 1073—1074. — P. 252—261. — doi:10.1016/j.brainres.2005.12.031. — PMID 16455061.
- Rusyniak D.E., Zaretskaia M.V., Zaretsky D.V., DiMicco J.A. 3,4-Methylenedioxymethamphetamine- and 8-hydroxy-2-di-n-propylamino-tetralin-induced hypothermia: role and location of 5-hydroxytryptamine 1A receptors (англ.) // J. Pharmacol. Exp. Ther. : journal. — 2007. — Vol. 323, no. 2. — P. 477—487. — doi:10.1124/jpet.107.126169. — PMID 17702902.
- Yu Y., Ramage A.G., Koss M.C. Pharmacological studies of 8-OH-DPAT-induced pupillary dilation in anesthetized rats (англ.) // Eur. J. Pharmacol. : journal. — 2004. — Vol. 489, no. 3. — P. 207—213. — doi:10.1016/j.ejphar.2004.03.007. — PMID 15087245.
- Prow M.R., Martin K.F., Heal D.J. 8-OH-DPAT-induced mydriasis in mice: a pharmacological characterisation (англ.) // Eur. J. Pharmacol. : journal. — 1996. — Vol. 317, no. 1. — P. 21—8. — doi:10.1016/S0014-2999(96)00693-0. — PMID 8982715.
- Fanciullacci M., Sicuteri R., Alessandri M., Geppetti P. Buspirone, but not sumatriptan, induces miosis in humans: relevance for a serotoninergic pupil control (англ.) // Clin. Pharmacol. Ther. : journal. — 1995. — March (vol. 57, no. 3). — P. 349—355. — doi:10.1016/0009-9236(95)90161-2. — PMID 7697953.
- Cohn J.B., Rickels K. A pooled, double-blind comparison of the effects of buspirone, diazepam and placebo in women with chronic anxiety (англ.) // Curr Med Res Opin : journal. — 1989. — Vol. 11, no. 5. — P. 304—320. — doi:10.1185/03007998909115213. — PMID 2649317.
- Cryan J.F., Redmond A.M., Kelly J.P., Leonard B.E. The effects of the 5-HT1A agonist flesinoxan, in three paradigms for assessing antidepressant potential in the rat (англ.) // European Neuropsychopharmacology : journal. — Elsevier, 1997. — Vol. 7, no. 2. — P. 109—114. — doi:10.1016/S0924-977X(96)00391-4. — PMID 9169298.
- Parks C.L., Robinson P.S., Sibille E., Shenk T., Toth M. Increased anxiety of mice lacking the serotonin1A receptor (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1998. — Vol. 95, no. 18. — P. 10734—10739. — doi:10.1073/pnas.95.18.10734. — PMID 9724773.
- Kennett G.A., Dourish C.T., Curzon G. Antidepressant-like action of 5-HT1A agonists and conventional antidepressants in an animal model of depression (англ.) // Eur. J. Pharmacol. : journal. — 1987. — Vol. 134, no. 3. — P. 265—274. — doi:10.1016/0014-2999(87)90357-8. — PMID 2883013.
- Keller M.B., Ruwe F.J., Janssens C.J., Sitsen J.M., Jokinen R., Janczewski J. Relapse prevention with gepirone ER in outpatients with major depression (англ.) // J Clin Psychopharmacol : journal. — 2005. — February (vol. 25, no. 1). — P. 79—84. — doi:10.1097/01.jcp.0000150221.53877.d9. — PMID 15643103.
- Cryan J.F., Redmond A.M., Kelly J.P., Leonard B.E. The effects of the 5-HT1A agonist flesinoxan, in three paradigms for assessing antidepressant potential in the rat (англ.) // European Neuropsychopharmacology : journal. — Elsevier, 1997. — May (vol. 7, no. 2). — P. 109—114. — doi:10.1016/S0924-977X(96)00391-4. — PMID 9169298.
- Invernizzi R.W., Sacchetti G., Parini S., Acconcia S., Samanin R. Flibanserin, a potential antidepressant drug, lowers 5-HT and raises dopamine and noradrenaline in the rat prefrontal cortex dialysate: role of 5-HT(1A) receptors (англ.) // Br. J. Pharmacol. : journal. — 2003. — August (vol. 139, no. 7). — P. 1281—1288. — doi:10.1038/sj.bjp.0705341. — PMID 12890707.
- de Paulis T. Drug evaluation: PRX-00023, a selective 5-HT1A receptor agonist for depression (англ.) // Curr Opin Investig Drugs : journal. — 2007. — Vol. 8, no. 1. — P. 78—86. — PMID 17263189.
- Stark A.D., Jordan S., Allers K.A., Bertekap R.L., Chen R., Mistry Kannan T., Molski T.F., Yocca F.D., Sharp T., Kikuchi T., Burris K.D. Interaction of the novel antipsychotic aripiprazole with 5-HT1A and 5-HT2A receptors: functional receptor-binding and in vivo electrophysiological studies (англ.) // Psychopharmacology : journal. — Springer, 2007. — Vol. 190, no. 3. — P. 373—382. — doi:10.1007/s00213-006-0621-y. — PMID 17242925.
- Wheeler Vega J.A., Mortimer A.M., Tyson P.J. Conventional antipsychotic prescription in unipolar depression, I: an audit and recommendations for practice (англ.) // J Clin Psychiatry : journal. — Physicians Postgraduate Press, 2003. — May (vol. 64, no. 5). — P. 568—574. — doi:10.4088/JCP.v64n0512. — PMID 12755661. Архивировано 20 июня 2009 года.
- Blier P., Abbott F.V. Putative mechanisms of action of antidepressant drugs in affective and anxiety disorders and pain (англ.) // Journal of Psychiatry & Neuroscience : journal. — Canadian Medical Association, 2001. — January (vol. 26, no. 1). — P. 37—43. — PMID 11212592. Архивировано 6 марта 2016 года.
- Morley K.C., Arnold J.C., McGregor I.S. Serotonin (1A) receptor involvement in acute 3,4-methylenedioxymethamphetamine (MDMA) facilitation of social interaction in the rat (англ.) // Prog. Neuropsychopharmacol. Biol. Psychiatry : journal. — 2005. — June (vol. 29, no. 5). — P. 648—657. — doi:10.1016/j.pnpbp.2005.04.009. — PMID 15908091.
- Thompson M.R., Callaghan P.D., Hunt G.E., Cornish J.L., McGregor I.S. A role for oxytocin and 5-HT(1A) receptors in the prosocial effects of 3,4 methylenedioxymethamphetamine ("ecstasy") (англ.) // Neuroscience : journal. — Elsevier, 2007. — May (vol. 146, no. 2). — P. 509—514. — doi:10.1016/j.neuroscience.2007.02.032. — PMID 17383105.
- Gobbi G., Cassano T., Radja F., Morgese M.G., Cuomo V., Santarelli L., Hen R., Blier P. Neurokinin 1 receptor antagonism requires norepinephrine to increase serotonin function (англ.) // European Neuropsychopharmacology : journal. — Elsevier, 2007. — April (vol. 17, no. 5). — P. 328—338. — doi:10.1016/j.euroneuro.2006.07.004. — PMID 16950604.
- Baker K.G., Halliday G.M., Hornung J.P., Geffen L.B., Cotton R.G., Törk I. Distribution, morphology and number of monoamine-synthesizing and substance P-containing neurons in the human dorsal raphe nucleus (англ.) // Neuroscience : journal. — Elsevier, 1991. — Vol. 42, no. 3. — P. 757—775. — doi:10.1016/0306-4522(91)90043-N. — PMID 1720227.
- Lucot J.B. Antiemetic effects of flesinoxan in cats: comparisons with 8-hydroxy-2-(di-n-propylamino)tetralin (англ.) // Eur. J. Pharmacol. : journal. — 1994. — February (vol. 253, no. 1—2). — P. 53—60. — doi:10.1016/0014-2999(94)90756-0. — PMID 8013549.
- Oshima T., Kasuya Y., Okumura Y., Terazawa E., Dohi S. Prevention of nausea and vomiting with tandospirone in adults after tympanoplasty (англ.) // Anesth. Analg. : journal. — 2002. — November (vol. 95, no. 5). — P. 1442—1445. — doi:10.1097/00000539-200211000-00063. — PMID 12401641.
- Bardin L., Tarayre J.P., Malfetes N., Koek W., Colpaert F.C. Profound, non-opioid analgesia produced by the high-efficacy 5-HT(1A) agonist F 13640 in the formalin model of tonic nociceptive pain (англ.) // Pharmacology : journal. — 2003. — April (vol. 67, no. 4). — P. 182—194. — doi:10.1159/000068404. — PMID 12595749.
- Colpaert F.C. 5-HT(1A) receptor activation: new molecular and neuroadaptive mechanisms of pain relief (англ.) // Curr Opin Investig Drugs : journal. — 2006. — January (vol. 7, no. 1). — P. 40—7. — PMID 16425670.
- Blier P., Gobbi G., Haddjeri N., Santarelli L., Mathew G., Hen R. Impact of substance P receptor antagonism on the serotonin and norepinephrine systems: relevance to the antidepressant/anxiolytic response (англ.) // Journal of Psychiatry & Neuroscience : journal. — Canadian Medical Association, 2004. — Vol. 29, no. 3. — P. 208—218. — PMID 15173897.
- Li Z., Ichikawa J., Dai J., Meltzer H.Y. Aripiprazole, a novel antipsychotic drug, preferentially increases dopamine release in the prefrontal cortex and hippocampus in rat brain (англ.) // Eur. J. Pharmacol. : journal. — 2004. — Vol. 493, no. 1—3. — P. 75—83. — doi:10.1016/j.ejphar.2004.04.028. — PMID 15189766.
- Bantick R.A., De Vries M.H., Grasby P.M. The effect of a 5-HT1A receptor agonist on striatal dopamine release (англ.) // Synapse : journal. — 2005. — Vol. 57, no. 2. — P. 67—75. — doi:10.1002/syn.20156. — PMID 15906386.
- Rollema H., Lu Y., Schmidt A.W., Sprouse J.S., Zorn S.H. 5-HT(1A) receptor activation contributes to ziprasidone-induced dopamine release in the rat prefrontal cortex (англ.) // Biol. Psychiatry : journal. — 2000. — Vol. 48, no. 3. — P. 229—237. — doi:10.1016/S0006-3223(00)00850-7. — PMID 10924666.
- Rollema H., Lu Y., Schmidt A.W., Zorn S.H. Clozapine increases dopamine release in prefrontal cortex by 5-HT1A receptor activation (англ.) // Eur. J. Pharmacol. : journal. — 1997. — Vol. 338, no. 2. — P. R3—5. — doi:10.1016/S0014-2999(97)81951-6. — PMID 9456005.
- Yoshino T., Nisijima K., Katoh S., Yui K., Nakamura M. Tandospirone potentiates the fluoxetine-induced increases in extracellular dopamine via 5-HT(1A) receptors in the rat medial frontal cortex (англ.) // Neurochem. Int. : journal. — 2002. — April (vol. 40, no. 4). — P. 355—360. — doi:10.1016/S0197-0186(01)00079-1. — PMID 11792466.
- Chojnacka-Wójcik E., Tatarczyńska E., Gołembiowska K., Przegaliński E. Involvement of 5-HT1A receptors in the antidepressant-like activity of gepirone in the forced swimming test in rats (англ.) // Neuropharmacology : journal. — 1991. — July (vol. 30, no. 7). — P. 711—717. — doi:10.1016/0028-3908(91)90178-E. — PMID 1681449.
- Волков В.П. Ятрогенные психонейросоматические синдромы. — Тверь: Триада, 2014. — 320 с.
- Ogren S.O., Eriksson T.M., Elvander-Tottie E., D'Addario C., Ekström J.C., Svenningsson P., Meister B., Kehr J., Stiedl O. The role of 5-HT(1A) receptors in learning and memory (англ.) // Behav. Brain Res. : journal. — 2008. — Vol. 195, no. 1. — P. 54—77. — doi:10.1016/j.bbr.2008.02.023. — PMID 18394726.
- Meltzer H.Y., Sumiyoshi T. Does stimulation of 5-HT(1A) receptors improve cognition in schizophrenia? (англ.) // Behav. Brain Res. : journal. — 2008. — December (vol. 195, no. 1). — P. 98—102. — doi:10.1016/j.bbr.2008.05.016. — PMID 18707769.
- Spreitzer H. Neue Wirkstoffe - Lecozotan (нем.) // Österreichische Apothekerzeitung : magazin. — 2008. — 13 August (Nr. 17/2007). — S. 805.
- de Boer S.F., Koolhaas J.M. 5-HT1A and 5-HT1B receptor agonists and aggression: a pharmacological challenge of the serotonin deficiency hypothesis (англ.) // Eur. J. Pharmacol. : journal. — 2005. — Vol. 526, no. 1—3. — P. 125—139. — doi:10.1016/j.ejphar.2005.09.065. — PMID 16310183.
- Olivier B., Mos J., Rasmussen D. Behavioural pharmacology of the serenic, eltoprazine (англ.) // Drug Metabol Drug Interact : journal. — 1990. — Vol. 8, no. 1—2. — P. 31—83. — doi:10.1515/DMDI.1990.8.1-2.31. — PMID 2091890.
- Thompson M.R., Callaghan P.D., Hunt G.E., Cornish J.L., McGregor I.S. A role for oxytocin and 5-HT(1A) receptors in the prosocial effects of 3,4 methylenedioxymethamphetamine ("ecstasy") (англ.) // Neuroscience : journal. — Elsevier, 2007. — Vol. 146, no. 2. — P. 509—514. — doi:10.1016/j.neuroscience.2007.02.032. — PMID 17383105.
- Winstanley C.A., Theobald D.E., Dalley J.W., Robbins T.W. Interactions between serotonin and dopamine in the control of impulsive choice in rats: therapeutic implications for impulse control disorders (англ.) // Neuropsychopharmacology : journal. — Nature Publishing Group, 2005. — Vol. 30, no. 4. — P. 669—682. — doi:10.1038/sj.npp.1300610. — PMID 15688093.
- Tomkins D.M., Higgins G.A., Sellers E.M. Low doses of the 5-HT1A agonist 8-hydroxy-2-(di-n-propylamino)-tetralin (8-OH DPAT) increase ethanol intake (англ.) // Psychopharmacology : journal. — Springer, 1994. — Vol. 115, no. 1—2. — P. 173—179. — doi:10.1007/BF02244769. — PMID 7862892.
- Müller C.P., Carey R.J., Huston J.P., De Souza Silva M.A. Serotonin and psychostimulant addiction: focus on 5-HT1A-receptors (англ.) // Prog. Neurobiol. : journal. — 2007. — Vol. 81, no. 3. — P. 133—178. — doi:10.1016/j.pneurobio.2007.01.001. — PMID 17316955.
- Carey R.J., DePalma G., Damianopoulos E., Shanahan A., Müller C.P., Huston J.P. Evidence that the 5-HT1A autoreceptor is an important pharmacological target for the modulation of cocaine behavioral stimulant effects (англ.) // Brain Res. : journal. — 2005. — Vol. 1034, no. 1—2. — P. 162—171. — doi:10.1016/j.brainres.2004.12.012. — PMID 15713268.
- Fernández-Guasti A., Rodríguez-Manzo G. 8-OH-DPAT and male rat sexual behavior: partial blockade by noradrenergic lesion and sexual exhaustion (англ.) // Pharmacol. Biochem. Behav. : journal. — 1997. — January (vol. 56, no. 1). — P. 111—116. — doi:10.1016/S0091-3057(96)00165-7. — PMID 8981617.
- Haensel S.M., Slob A.K. Flesinoxan: a prosexual drug for male rats (англ.) // Eur. J. Pharmacol. : journal. — 1997. — July (vol. 330, no. 1). — P. 1—9. — doi:10.1016/S0014-2999(97)00170-2. — PMID 9228408.
- Simon P., Guardiola B., Bizot-Espiard J., Schiavi P., Costentin J. 5-HT1A receptor agonists prevent in rats the yawning and penile erections induced by direct dopamine agonists (англ.) // Psychopharmacology : journal. — Springer, 1992. — Vol. 108, no. 1—2. — P. 47—50. — doi:10.1007/BF02245284. — PMID 1357709.
- Millan M.J., Perrin-Monneyron S. Potentiation of fluoxetine-induced penile erections by combined blockade of 5-HT1A and 5-HT1B receptors (англ.) // Eur. J. Pharmacol. : journal. — 1997. — Vol. 321, no. 3. — P. R11—3. — doi:10.1016/S0014-2999(97)00050-2. — PMID 9085055.
- Ebenezer I.S., Arkle M.J., Tite R.M. 8-Hydroxy-2-(di-n-propylamino)-tetralin inhibits food intake in fasted rats by an action at 5-HT1A receptors (англ.) // Methods Find Exp Clin Pharmacol : journal. — 2007. — Vol. 29, no. 4. — P. 269—272. — doi:10.1358/mf.2007.29.4.1075362. — PMID 17609739.
- Monti J.M., Jantos H. Dose-dependent effects of the 5-HT1A receptor agonist 8-OH-DPAT on sleep and wakefulness in the rat (англ.) // J Sleep Res : journal. — 1992. — Vol. 1, no. 3. — P. 169—175. — doi:10.1111/j.1365-2869.1992.tb00033.x. — PMID 10607047.
- Marc Ansseau, William Pitchot, Antonio Gonzalez Moreno, Jacques Wauthy, Patrick Papart. Pilot study of flesinoxan, a 5-HT1A agonist, in major depression: Effects on sleep REM latency and body temperature (англ.) // Human Psychopharmacology: Clinical and Experimental : journal. — 2004. — Vol. 8, no. 4. — P. 279—283. — doi:10.1002/hup.470080407. Архивировано 17 декабря 2012 года.
- Meyer L.C., Fuller A., Mitchell D. Zacopride and 8-OH-DPAT reverse opioid-induced respiratory depression and hypoxia but not catatonic immobilization in goats (англ.) // American Physiological Society : journal. — 2006. — February (vol. 290, no. 2). — P. R405—13. — doi:10.1152/ajpregu.00440.2005. — PMID 16166206.
- Van de Kar L.D., Levy A.D., Li Q., Brownfield M.S. A comparison of the oxytocin and vasopressin responses to the 5-HT1A agonist and potential anxiolytic drug alnespirone (S-20499) (англ.) // Pharmacol. Biochem. Behav. : journal. — 1998. — Vol. 60, no. 3. — P. 677—683. — doi:10.1016/S0091-3057(98)00025-2. — PMID 9678651.
- Lorens S.A., Van de Kar L.D. Differential effects of serotonin (5-HT1A and 5-HT2) agonists and antagonists on renin and corticosterone secretion (англ.) // Neuroendocrinology : journal. — 1987. — Vol. 45, no. 4. — P. 305—310. — doi:10.1159/000124754. — PMID 2952898.
- Koenig J.I., Gudelsky G.A., Meltzer H.Y. Stimulation of corticosterone and beta-endorphin secretion in the rat by selective 5-HT receptor subtype activation (англ.) // Eur. J. Pharmacol. : journal. — 1987. — Vol. 137, no. 1. — P. 1—8. — doi:10.1016/0014-2999(87)90175-0. — PMID 2956114.
- Pitchot W., Wauthy J., Legros J.J., Ansseau M. Hormonal and temperature responses to flesinoxan in normal volunteers: an antagonist study (англ.) // European Neuropsychopharmacology : journal. — Elsevier, 2004. — March (vol. 14, no. 2). — P. 151—155. — doi:10.1016/S0924-977X(03)00108-1. — PMID 15013031.
- Navinés R., Martín-Santos R., Gómez-Gil E., Martínez de Osaba M.J., Gastó C. Interaction between serotonin 5-HT1A receptors and beta-endorphins modulates antidepressant response (англ.) // Prog. Neuropsychopharmacol. Biol. Psychiatry : journal. — 2008. — December (vol. 32, no. 8). — P. 1804—1809. — doi:10.1016/j.pnpbp.2008.07.021. — PMID 18725263.
- Hjorth S., Bengtsson H.J., Kullberg A., Carlzon D., Peilot H., Auerbach S.B. Serotonin autoreceptor function and antidepressant drug action (англ.) // J. Psychopharmacol. (Oxford) : journal. — 2000. — Vol. 14, no. 2. — P. 177—185. — doi:10.1177/026988110001400208. — PMID 10890313.
- Briley M., Moret C. Neurobiological mechanisms involved in antidepressant therapies (англ.) // Clin Neuropharmacol : journal. — 1993. — Vol. 16, no. 5. — P. 387—400. — doi:10.1097/00002826-199310000-00002. — PMID 8221701.
- Starr K.R., Price G.W., Watson J.M., Atkinson P.J., Arban R., Melotto S., Dawson L.A., Hagan J.J., Upton N., Duxon M.S. SB-649915-B, a novel 5-HT1A/B autoreceptor antagonist and serotonin reuptake inhibitor, is anxiolytic and displays fast onset activity in the rat high light social interaction test (англ.) // Neuropsychopharmacology : journal. — Nature Publishing Group, 2007. — Vol. 32, no. 10. — P. 2163—2172. — doi:10.1038/sj.npp.1301341. — PMID 17356576.
- Rothman R.B., Baumann M.H. Therapeutic potential of monoamine transporter substrates (англ.) // Curr Top Med Chem : journal. — 2006. — Vol. 6, no. 17. — P. 1845—1859. — doi:10.2174/156802606778249766. — PMID 17017961. Архивировано 26 марта 2017 года.
- Scorza C., Silveira R., Nichols D.E., Reyes-Parada M. Effects of 5-HT-releasing agents on the extracellullar hippocampal 5-HT of rats. Implications for the development of novel antidepressants with a short onset of action (англ.) // Neuropharmacology : journal. — 1999. — July (vol. 38, no. 7). — P. 1055—1061. — doi:10.1016/S0028-3908(99)00023-4. — PMID 10428424.
- Marona-Lewicka D., Nichols D.E. Drug discrimination studies of the interoceptive cues produced by selective serotonin uptake inhibitors and selective serotonin releasing agents (англ.) // Psychopharmacology : journal. — Springer, 1998. — July (vol. 138, no. 1). — P. 67—75. — doi:10.1007/s002130050646. — PMID 9694528. Архивировано 12 января 2002 года.
- Drago A., Ronchi D.D., Serretti A. 5-HT1A gene variants and psychiatric disorders: a review of current literature and selection of SNPs for future studies (англ.) // Int. J. Neuropsychopharmacol. : journal. — 2008. — August (vol. 11, no. 5). — P. 701—721. — doi:10.1017/S1461145707008218. — PMID 18047755.
- Wu S., Comings D.E. A common C-1018G polymorphism in the human 5-HT1A receptor gene (англ.) // Genetics : journal. — Genetics Society of America, 1999. — June (vol. 9, no. 2). — P. 105—106. — doi:10.1097/00041444-199906000-00010. — PMID 10412191.
- Anttila S., Huuhka K., Huuhka M., Rontu R., Hurme M., Leinonen E., Lehtimäki T. Interaction between 5-HT1A and BDNF genotypes increases the risk of treatment-resistant depression (англ.) // J Neural Transm : journal. — 2007. — Vol. 114, no. 8. — P. 1065—1068. — doi:10.1007/s00702-007-0705-9. — PMID 17401528.
- Guiard B.P., David D.J., Deltheil T., Chenu F., Le Maître E., Renoir T., Leroux-Nicollet I., Sokoloff P., Lanfumey L., Hamon M., Andrews A.M., Hen R., Gardier A.M. Brain-derived neurotrophic factor-deficient mice exhibit a hippocampal hyperserotonergic phenotype (англ.) // Int. J. Neuropsychopharmacol. : journal. — 2008. — Vol. 11, no. 1. — P. 79—92. — doi:10.1017/S1461145707007857. — PMID 17559709.
- Pike V.W., McCarron J.A., Lammerstma A.A., Hume S.P., Poole K., Grasby P.M., Malizia A., Cliffe I.A., Fletcher A., Bench C.J. First delineation of 5-HT1A receptors in human brain with PET and [11C]WAY-100635 (англ.) // Eur. J. Pharmacol. : journal. — 1995. — Vol. 283, no. 1—3. — P. R1—3. — doi:10.1016/0014-2999(95)00438-Q. — PMID 7498295.
- Price J.C., Kelley D.E., Ryan C.M., Meltzer C.C., Drevets W.C., Mathis C.A., Mazumdar S., Reynolds C.F. Evidence of increased serotonin-1A receptor binding in type 2 diabetes: a positron emission tomography study (англ.) // Brain Res. : journal. — 2002. — Vol. 927, no. 1. — P. 97—103. — doi:10.1016/S0006-8993(01)03297-8. — PMID 11814436.
- Borg J., Andrée B., Soderstrom H., Farde L. The serotonin system and spiritual experiences (англ.) // American Journal of Psychiatry : journal. — 2003. — November (vol. 160, no. 11). — P. 1965—1969. — doi:10.1176/appi.ajp.160.11.1965. — PMID 14594742.
- Burnet P.W., Eastwood S.L., Harrison P.J. [3H]WAY-100635 for 5-HT1A receptor autoradiography in human brain: a comparison with [3H]8-OH-DPAT and demonstration of increased binding in the frontal cortex in schizophrenia (англ.) // Neurochem. Int. : journal. — 1997. — Vol. 30, no. 6. — P. 565—574. — doi:10.1016/S0197-0186(96)00124-6. — PMID 9152998.
- Winsauer P.J., Rodriguez F.H., Cha A.E., Moerschbaecher J.M. Full and partial 5-HT1A receptor agonists disrupt learning and performance in rats (англ.) // J. Pharmacol. Exp. Ther. : journal. — 1999. — January (vol. 288, no. 1). — P. 335—347. — PMID 9862788.
- Griebel G., Misslin R., Pawlowski M., Guardiola Lemaître B., Guillaumet G., Bizot-Espiard J. Anxiolytic-like effects of a selective 5-HT1A agonist, S20244, and its enantiomers in mice (англ.) // Neuroreport. : journal. — 1992. — Vol. 3, no. 1. — P. 84—86. — doi:10.1097/00001756-199201000-00022. — PMID 1351756.
- Simon P., Guardiola B., Bizot-Espiard J., Schiavi P., Costentin J. 5-HT1A receptor agonists prevent in rats the yawning and penile erections induced by direct dopamine agonists (англ.) // Psychopharmacology : journal. — Springer, 1992. — Vol. 108, no. 1—2. — P. 47—50. — doi:10.1007/BF02245284. — PMID 1357709.
- Astier B., Lambás Señas L., Soulière F., Schmitt P., Urbain N., Rentero N., Bert L., Denoroy L., Renaud B., Lesourd M., Muñoz C., Chouvet G. In vivo comparison of two 5-HT1A receptors agonists alnespirone (S-20499) and buspirone on locus coeruleus neuronal activity (англ.) // Eur J Pharmacol. : journal. — 2003. — Vol. 459, no. 1. — P. 17—26. — doi:10.1016/S0014-2999(02)02814-5. — PMID 12505530.
- Bruins Slot L.A., Koek W., Tarayre J.P., Colpaert F.C. Tolerance and inverse tolerance to the hyperalgesic and analgesic actions, respectively, of the novel analgesic, F 13640 (англ.) // European Journal of Pharmacology : journal. — 2003. — April (vol. 466, no. 3). — P. 271—279. — doi:10.1016/S0014-2999(03)01566-8. — PMID 12694810.
- Bardin L., Assié M.B., Pélissou M., Royer-Urios I., Newman-Tancredi A., Ribet J.P., Sautel F., Koek W., Colpaert F.C. Dual, hyperalgesic, and analgesic effects of the high-efficacy 5-hydroxytryptamine 1A (5-HT1A) agonist F 13640 [(3-chloro-4-fluoro-phenyl)-[4-fluoro-4-{[(5-methyl-pyridin-2-ylmethyl)-amino]-methyl}piperidin-1-yl]methanone, fumaric acid salt]: relationship with 5-HT1A receptor occupancy and kinetic parameters (англ.) // The Journal of Pharmacology and Experimental Therapeutics : journal. — 2005. — March (vol. 312, no. 3). — P. 1034—1042. — doi:10.1124/jpet.104.077669. — PMID 15528450.
- Colpaert F.C., Deseure K., Stinus L., Adriaensen H. High-efficacy 5-hydroxytryptamine 1A receptor activation counteracts opioid hyperallodynia and affective conditioning (англ.) // The Journal of Pharmacology and Experimental Therapeutics : journal. — 2006. — February (vol. 316, no. 2). — P. 892—899. — doi:10.1124/jpet.105.095109. — PMID 16254131.
- Deseure K., Bréand S., Colpaert F.C. Curative-like analgesia in a neuropathic pain model: parametric analysis of the dose and the duration of treatment with a high-efficacy 5-HT(1A) receptor agonist (англ.) // European Journal of Pharmacology : journal. — 2007. — July (vol. 568, no. 1—3). — P. 134—141. — doi:10.1016/j.ejphar.2007.04.022. — PMID 17512927.
- Bernard Vacher, Bernard Bonnaud, Wouter Koek. Pyridin-2-yl-methylamine derivatives, method of preparing and application as medicine. US Patent 6020345, May 21, 1999.
- Koek W., Patoiseau J.F., Assié M.B., et al. F 11440, a potent, selective, high efficacy 5-HT1A receptor agonist with marked anxiolytic and antidepressant potential (англ.) // The Journal of Pharmacology and Experimental Therapeutics : journal. — 1998. — October (vol. 287, no. 1). — P. 266—283. — PMID 9765347.
- Prinssen E.P., Colpaert F.C., Koek W. 5-HT1A receptor activation and anti-cataleptic effects: high-efficacy agonists maximally inhibit haloperidol-induced catalepsy (англ.) // European Journal of Pharmacology : journal. — 2002. — October (vol. 453, no. 2—3). — P. 217—221. — doi:10.1016/S0014-2999(02)02430-5. — PMID 12398907.
- Maurel J.L., Autin J.M., Funes P., Newman-Tancredi A., Colpaert F., Vacher B. High-efficacy 5-HT1A agonists for antidepressant treatment: a renewed opportunity (англ.) // Journal of Medicinal Chemistry : journal. — 2007. — October (vol. 50, no. 20). — P. 5024—5033. — doi:10.1021/jm070714l. — PMID 17803293.
- Newman-Tancredi A., Martel J.C., Assié M.B., et al. Signal transduction and functional selectivity of F15599, a preferential post-synaptic 5-HT1A receptor agonist (англ.) // British Journal of Pharmacology : journal. — 2009. — January (vol. 156, no. 2). — P. 338—353. — doi:10.1111/j.1476-5381.2008.00001.x. — PMID 19154445.
- Haj-Dahmane S., Jolas T., Laporte A.M., et al. Interactions of lesopitron (E-4424) with central 5-HT1A receptors: in vitro and in vivo studies in the rat (англ.) // European Journal of Pharmacology : journal. — 1994. — April (vol. 255, no. 1—3). — P. 185—196. — doi:10.1016/0014-2999(94)90097-3. — PMID 8026543.
- Monte A.P., Marona-Lewicka D., Lewis M.M., Mailman R.B., Wainscott D.B., Nelson D.L., Nichols D.E. Substituted naphthofurans as hallucinogenic phenethylamine-ergoline hybrid molecules with unexpected muscarinic antagonist activity (англ.) // Journal of Medicinal Chemistry : journal. — 1998. — June (vol. 41, no. 12). — P. 2134—2145. — doi:10.1021/jm980076u. — PMID 9622555.
- Foreman M.M., Fuller R.W., Rasmussen K., Nelson D.L., Calligaro D.O., Zhang L., Barrett J.E., Booher R.N., Paget C.J., Flaugh M.E. Pharmacological characterization of LY293284: A 5-HT1A receptor agonist with high potency and selectivity (англ.) // The Journal of Pharmacology and Experimental Therapeutics : journal. — 1994. — September (vol. 270, no. 3). — P. 1270—1281. — PMID 7523657.
- Matsuda T., Yoshikawa T., Suzuki M., Asano S., Somboonthum P., Takuma K., Nakano Y., Morita T., Nakasu Y., Kim H. S. Novel benzodioxan derivative, 5-(3-[((2S)-1,4-benzodioxan-2- ylmethyl)aminopropoxy)-1,3-benzodioxole HCl (MKC-242), with a highly potent and selective agonist activity at rat central serotonin1A receptors.] (англ.) // Japanese journal of pharmacology. — 1995. — Vol. 69, no. 4. — P. 357—366. — PMID 8786639.
- De Vry J., Schohe-Loop R., Heine H. G., Greuel J. M., Mauler F., Schmidt B., Sommermeyer H., Glaser T. Characterization of the aminomethylchroman derivative BAY x 3702 as a highly potent 5-hydroxytryptamine1A receptor agonist. (англ.) // The Journal of pharmacology and experimental therapeutics. — 1998. — Vol. 284, no. 3. — P. 1082—1094. — PMID 9495870.
- Dong J., de Montigny C., Blier P. Full agonistic properties of BAY x 3702 on presynaptic and postsynaptic 5-HT1A receptors electrophysiological studies in the rat hippocampus and dorsal raphe. (англ.) // The Journal of pharmacology and experimental therapeutics. — 1998. — Vol. 286, no. 3. — P. 1239—1247. — PMID 9732384.
- Romero A.G., Leiby J.A., McCall R.B., Piercey M.F., Smith M.W., Han F. Novel 2-substituted tetrahydro-3H-benz[e]indolamines: highly potent and selective agonists acting at the 5-HT1A receptor as possible anxiolytics and antidepressants (англ.) // J Med Chem. : journal. — 1993. — Vol. 36, no. 15. — P. 2066—2074. — doi:10.1021/jm00067a003. — PMID 8101876.
- McCall R.B., Romero A.G., Bienkowski M.J., Harris D.W., McGuire J.C., Piercey M.F., Shuck M.E., Smith M.W., Svensson K.A., Schreur P.J., et al. Characterization of U-92016A as a selective, orally active, high intrinsic activity 5-hydroxytryptamine1A agonist (англ.) // J Pharmacol Exp Ther. : journal. — 1994. — Vol. 271, no. 2. — P. 875—883. — PMID 7965808.
- Winter J.C., Timineri D. The discriminative stimulus properties of EGb 761, an extract of Ginkgo biloba (англ.) // Pharmacol. Biochem. Behav. : journal. — 1999. — March (vol. 62, no. 3). — P. 543—547. — doi:10.1016/S0091-3057(98)00190-7. — PMID 10080249.
- Dr Beth, PhD. Limonene strains can produce and anti-anxiety effect. (англ.). Torrey Holistics (13 июля 2019). Дата обращения: 7 сентября 2020.
- Bjorvatn B, Neckelmann D, Ursin R. The 5-HT1A antagonist (-)-alprenolol fails to modify sleep or zimeldine-induced sleep-waking effects in rats // Pharmacol Biochem Behav.. — May 1992. — Т. 42, вып. 42(1), № 1. — С. 49—56. — PMID 1388278.
- Sánchez-López A, Centurión D, Lozano-Cuenca J, Muñoz-Islas E, Cobos-Puc LE, Villalón CM. Role of serotonin receptors in vascular tone in the pithed rat (исп.) // Arch Cardiol Mex.. — Dec 2009. — V. 79, fasc. 79 (Suppl 2), no Suppl 2. — PMID 20361490.
- Adham N1, Tamm JA, Salon JA, Vaysse PJ, Weinshank RL, Branchek TA. A single point mutation increases the affinity of serotonin 5-HT1D alpha, 5-HT1D beta, 5-HT1E and 5-HT1F receptors for beta-adrenergic antagonists (англ.) // Neuropharmacology. — Mar-Apr 1994. — Vol. 33, iss. 33(3—4), no. 3—4. — P. 387—391. — PMID 7984276.
- Waldmeier PC1, Williams M, Baumann PA, Bischoff S, Sills MA, Neale RF. Interactions of isamoltane (CGP 361A), an anxiolytic phenoxypropanolamine derivative, with 5-HT1 receptor subtypes in the rat brain // Naunyn Schmiedebergs Arch Pharmacol.. — Jun 1988. — Т. 337, вып. 337(6), № 6. — С. 609—620. — PMID 2905765.
- Seletti B, Benkelfat C, Blier P, Annable L, Gilbert F, de Montigny C. Serotonin1A receptor activation by flesinoxan in humans. Body temperature and neuroendocrine responses (англ.) // Neuropsychopharmacology. — Nature Publishing Group, Oct 1995. — Vol. 13, iss. 13(2), no. 2. — P. 93—104. — PMID 8597527.
- Newman-Tancredi A, Gavaudan S, Conte C, Chaput C, Touzard M, Verrièle L, Audinot V, Millan MJ. Agonist and antagonist actions of antipsychotic agents at 5-HT1A receptors: a [35SGTPgammaS binding study] (англ.) // Eur J Pharmacol.. — 21 Aug 1998. — Vol. 355, iss. 355(2—3), no. 2—3. — P. 245—256. — PMID 9760039.
- Roth, BL; Driscol, J. PDSP Ki Database. Psychoactive Drug Screening Program (PDSP). University of North Carolina at Chapel Hill and the United States National Institute of Mental Health (12 января 2011). Дата обращения: 4 ноября 2013. Архивировано 8 ноября 2013 года.
- Monachon M.A., Burkard W.P., Jalfre M., Haefely W. Blockade of central 5-hydroxytryptamine receptors by methiothepin (англ.) // Naunyn Schmiedebergs Arch. Pharmacol. : journal. — 1972. — Vol. 274, no. 2. — P. 192—197. — doi:10.1007/BF00501854. — PMID 4340797.
- Goetz A.S., King H.K., Ward S.D., True T.A., Rimele T.J., Saussy D.L. BMY 7378 is a selective antagonist of the D subtype of alpha 1-adrenoceptors (англ.) // European Journal of Pharmacology : journal. — 1995. — January (vol. 272, no. 2—3). — P. R5—6. — doi:10.1016/0014-2999(94)00751-R. — PMID 7713154.
- Farré, M; Roset, PN; Llorente, M; Márquez, M; Albet, C; Pérez, JA; Herrero, E; Ortíz, J.A. Clinical pharmacokinetics and tolerability of dotarizine in healthy subjects after single and multiple oral administration (англ.) // Methods and findings in experimental and clinical pharmacology : journal. — 1997. — Vol. 19, no. 5. — P. 343—350. — PMID 9379783.
- Montiel, C; Herrero, CJ; García-Palomero, E; Renart, J; García, AG; Lomax, R.B. Serotonergic effects of dotarizine in coronary artery and in oocytes expressing 5-HT2 receptors (англ.) // European Journal of Pharmacology : journal. — 1997. — Vol. 332, no. 2. — P. 183—193. — doi:10.1016/S0014-2999(97)01073-X. — PMID 9286620.
- Dictionary of Organic Compounds ... - Google Books.
- Rényi L., Larsson L.G., Berg S., Svensson B.E., Thorell G., Ross SB. Biochemical and behavioural effects of isamoltane, a beta-adrenoceptor antagonist with affinity for the 5-HT1B receptor of rat brain (англ.) // Naunyn Schmiedebergs Arch Pharmacol. : journal. — 1991. — Vol. 343, no. 1. — P. 1—6. — doi:10.1007/bf00180669. — PMID 1674359.
- Waldmeier P.C., Williams M., Baumann P.A., Bischoff S., Sills M.A., Neale RF. Interactions of isamoltane (CGP 361A), an anxiolytic phenoxypropanolamine derivative, with 5-HT1 receptor subtypes in the rat brain (англ.) // Naunyn Schmiedebergs Arch Pharmacol. : journal. — 1988. — Vol. 337, no. 6. — P. 609—620. — doi:10.1007/bf00175785. — PMID 2905765.
- Schlechter, LE; Smith, DL; Rosenzweig-Lipson, S; Sukoff, SJ; Dawson, LA; Marquis, K; Jones, D; Piesla, M; Andree, T; Nawoschik, S; Harder, J. A.; Womack, M. D.; Buccafusco, J; Terry, A. V.; Hoebel, B; Rada, P; Kelly, M; Abou-Gharbia, M; Barrett, J. E.; Childers, W. Lecotozan (SRA-333): A selective serotonin1A receptor antagonist that enhances the stimulated release of glutamate and acetylcholine in the hippocampus and promotes procognitive effects (англ.) // Journal of Pharmacology and Experimental Therapeutics : journal. — 2005. — 10 June (vol. 314, no. 3). — P. 1274—1289. — doi:10.1124/jpet.105.086363. — PMID 15951399.
- D. Le Bars. High-Yield Radiosynthesis and Preliminary In Vivo Evaluation of p-18FMPPF, a Fluoro Analog of WAY-100635 (англ.) // Nuclear Medicine and Biology : journal. — 1998. — May (vol. 25, no. 4). — P. 343—350. — doi:10.1016/S0969-8051(97)00229-1.
- Foong J.P., Bornstein JC. 5-HT antagonists NAN-190 and SB 269970 block alpha2-adrenoceptors in the guinea pig (англ.) // Neuroreport. : journal. — 2009. — Vol. 20, no. 3. — P. 325—330. — doi:10.1097/WNR.0b013e3283232caa. — PMID 19190523.
- Bell, R; Hobson, H. Effects of pindobind 5-hydroxytryptamine1A (5-HT1A), a novel and potent 5-HT1A antagonist, on social and agonistic behaviour in male albino mice (англ.) // Pharmacology, Biochemistry, and Behavior : journal. — 1993. — Vol. 46, no. 1. — P. 67—72. — doi:10.1016/0091-3057(93)90318-N. — PMID 8255924.
- Jerning E., Svantesson G.T., Mohell N. Receptor binding characteristics of [3H]NAD-299, a new selective 5-HT1A receptor antagonist (англ.) // Eur J Pharmacol. : journal. — 1998. — Vol. 360, no. 2—3. — P. 219—225. — doi:10.1016/S0014-2999(98)00667-0. — PMID 9851589.
- Starr K.R., Price G.W., Watson J.M., Atkinson P.J., Arban R., Melotto S., Dawson L.A., Hagan J.J., Upton N., Duxon MS. SB-649915-B, a novel 5-HT1A/B autoreceptor antagonist and serotonin reuptake inhibitor, is anxiolytic and displays fast onset activity in the rat high light social interaction test (англ.) // Neuropsychopharmacology. : journal. — 2007. — Vol. 32, no. 10. — P. 2163—2172. — doi:10.1038/sj.npp.1301341. — PMID 17356576.
- Czoty P.W., Howell L.L. Behavioral effects of AMI-193, a 5-HT(2A)- and dopamine D(2)-receptor antagonist, in the squirrel monkey (англ.) // Pharmacology, Biochemistry, and Behavior : journal. — 2000. — October (vol. 67, no. 2). — P. 257—264. — doi:10.1016/S0091-3057(00)00321-X. — PMID 11124389.
- Luparini M.R., Garrone B., Pazzagli M., Pinza M., Pepeu G. A cortical GABA-5HT interaction in the mechanism of action of the antidepressant trazodone (англ.) // Progress in Neuro-psychopharmacology & Biological Psychiatry : journal. — 2004. — November (vol. 28, no. 7). — P. 1117—1127. — doi:10.1016/j.pnpbp.2004.05.046. — PMID 15610924.
- Hamada K., Yoshida M., Isayama H., Yagi Y., Kanazashi S., Kashihara Y., Takeuchi K., Yamaguchi I. Possible involvement of endogenous 5-HT in aggravation of cerulein-induced acute pancreatitis in mice (англ.) // Journal of Pharmacological Sciences : journal. — 2007. — November (vol. 105, no. 3). — P. 240—250. — doi:10.1254/jphs.FP0071049. — PMID 17965538.
- Terrón JA, Ibarra M, Ransanz V, Hong E, Villalón CM. The alpha-antiadrenergic properties of spiroxatrine, a ligand of serotonergic 5-HT1A receptors (исп.) // Arch Inst Cardiol Mex.. — Jul-Aug 1993. — V. 63, fasc. 63(4), no 4. — P. 289—295. — PMID 8105762.
- Björk L., Cornfield L.J., Nelson D.L., Hillver S.E., Andén N.E., Lewander T., Hacksell U. Pharmacology of the novel 5-hydroxytryptamine1A receptor antagonist (S)-5-fluoro-8-hydroxy-2-(dipropylamino)tetralin: inhibition of (R)-8-hydroxy-2-(dipropylamino)tetralin-induced effects (англ.) // J Pharmacol Exp Ther. : journal. — 1991. — Vol. 258, no. 1. — P. 58—65. — PMID 1830099.
- Cliffe I.A., Brightwell C.I., Fletcher A., et al. (S)-N-tert-butyl-3-(4-(2-methoxyphenyl)-piperazin-1-yl)-2-phenylpropanamide [(S)-WAY-100135]: a selective antagonist at presynaptic and postsynaptic 5-HT1A receptors (англ.) // Journal of Medicinal Chemistry : journal. — 1993. — May (vol. 36, no. 10). — P. 1509—1510. — doi:10.1021/jm00062a028. — PMID 8496920.
- A. Fletcher, E. A. Forster, D. J. Bill, G. Brown, I. A. Cliffe, J. E. Hartley, D. E. Jones, A. McLenachan, K. J. Stanhope, D. J. Critchley, K. J. Childs, V. C. Middlefell, L. Lanfumey, R. Corradetti, A. M. Laporte, H. Gozlan, M. Hamon & C. T. Dourish. Electrophysiological, biochemical, neurohormonal and behavioural studies with WAY-100635, a potent, selective and silent 5-HT1A receptor antagonist (англ.) // Behavioural Brain Research : journal. — 1996. — Vol. 73, no. 1—2. — P. 337—333. — doi:10.1016/0166-4328(96)00118-0. — PMID 8788530.
- Fuller, RW; Kurz, KD; Mason, NR; Cohen, M.L. Antagonism of a peripheral vascular but not an apparently central serotonergic response by xylamidine and BW 501C67 (англ.) // European Journal of Pharmacology : journal. — 1986. — Vol. 125, no. 1. — P. 71—7. — doi:10.1016/0014-2999(86)90084-1. — PMID 3732393.
- Dedeoğlu, A; Fisher, L.A. Central and peripheral injections of the 5-HT2 agonist, 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane, modify cardiovascular function through different mechanisms (англ.) // The Journal of Pharmacology and Experimental Therapeutics : journal. — 1991. — Vol. 259, no. 3. — P. 1027—1034. — PMID 1762059.
- Baker, BJ; Duggan, JP; Barber, DJ; Booth, D.A. Effects of dl-fenfluramine and xylamidine on gastric emptying of maintenance diet in freely feeding rats (англ.) // European Journal of Pharmacology : journal. — 1988. — Vol. 150, no. 1—2. — P. 137—142. — doi:10.1016/0014-2999(88)90759-5. — PMID 3402534.
- Saigal N., Pichika R., Easwaramoorthy B., Collins D., Christian B. T., Shi B., Narayanan T. K., Potkin S. G., Mukherjee J. Synthesis and biologic evaluation of a novel serotonin 5-HT1A receptor radioligand, 18F-labeled mefway, in rodents and imaging by PET in a nonhuman primate. (англ.) // Journal of nuclear medicine : official publication, Society of Nuclear Medicine. — 2006. — Vol. 47, no. 10. — P. 1697—1706. — PMID 17015907.
Для дополнительного чтения
- el Mestikawy S., Fargin A., Raymond J.R., Gozlan H., Hnatowich M. The 5-HT1A receptor: an overview of recent advances (англ.) // Neurochem. Res. : journal. — 1991. — Vol. 16, no. 1. — P. 1—10. — doi:10.1007/BF00965820. — PMID 2052135.
- Hensler J.G. Regulation of 5-HT1A receptor function in brain following agonist or antidepressant administration (англ.) // Life Sci. : journal. — 2003. — Vol. 72, no. 15. — P. 1665—1682. — doi:10.1016/S0024-3205(02)02482-7. — PMID 12559389.
- Van Oekelen D., Luyten W.H., Leysen J.E. 5-HT2A and 5-HT2C receptors and their atypical regulation properties (англ.) // Life Sci. : journal. — 2003. — Vol. 72, no. 22. — P. 2429—2449. — doi:10.1016/S0024-3205(03)00141-3. — PMID 12650852.
- Lesch K.P., Gutknecht L. Focus on The 5-HT1A receptor: emerging role of a gene regulatory variant in psychopathology and pharmacogenetics (англ.) // Int. J. Neuropsychopharmacol. : journal. — 2004. — Vol. 7, no. 4. — P. 381—385. — doi:10.1017/S1461145704004845. — PMID 15683551.
- Kalipatnapu S., Chattopadhyay A. Membrane protein solubilization: recent advances and challenges in solubilization of serotonin1A receptors (англ.) // IUBMB Life : journal. — 2005. — Vol. 57, no. 7. — P. 505—512. — doi:10.1080/15216540500167237. — PMID 16081372.
- Varrault A., Bockaert J., Waeber C. Activation of 5-HT1A receptors expressed in NIH-3T3 cells induces focus formation and potentiates EGF effect on DNA synthesis (англ.) // Molecular Biology of the Cell : journal. — 1992. — Vol. 3, no. 9. — P. 961—969. — doi:10.1091/mbc.3.9.961. — PMID 1330092.
- Levy F.O., Gudermann T., Perez-Reyes E., Birnbaumer M., Kaumann A.J., Birnbaumer L. Molecular cloning of a human serotonin receptor (S12) with a pharmacological profile resembling that of the 5-HT1D subtype (англ.) // J. Biol. Chem. : journal. — 1992. — Vol. 267, no. 11. — P. 7553—7562. — PMID 1559993.
- Melmer G., Sherrington R., Mankoo B., Kalsi G., Curtis D., Gurling H.M. A cosmid clone for the 5HT1A receptor (HTR1A) reveals a TaqI RFLP that shows tight linkage to dna loci D5S6, D5S39, and D5S76 (англ.) // Genomics : journal. — Academic Press, 1991. — Vol. 11, no. 3. — P. 767—769. — doi:10.1016/0888-7543(91)90088-V. — PMID 1685484.
- Parks C.L., Chang L.S., Shenk T. A polymerase chain reaction mediated by a single primer: cloning of genomic sequences adjacent to a serotonin receptor protein coding region (англ.) // Nucleic Acids Res. : journal. — 1991. — Vol. 19, no. 25. — P. 7155—7160. — doi:10.1093/nar/19.25.7155. — PMID 1766875.
- Gilliam T.C., Freimer N.B., Kaufmann C.A., Powchik P.P., Bassett A.S., Bengtsson U., Wasmuth J.J. Deletion mapping of DNA markers to a region of chromosome 5 that cosegregates with schizophrenia (англ.) // Genomics : journal. — Academic Press, 1989. — Vol. 5, no. 4. — P. 940—944. — doi:10.1016/0888-7543(89)90138-9. — PMID 2591972.
- Kobilka B.K., Frielle T., Collins S., Yang-Feng T., Kobilka T.S., Francke U., Lefkowitz R.J., Caron M.G. An intronless gene encoding a potential member of the family of receptors coupled to guanine nucleotide regulatory proteins (англ.) // Nature : journal. — 1987. — Vol. 329, no. 6134. — P. 75—9. — doi:10.1038/329075a0. — PMID 3041227.
- Fargin A., Raymond J.R., Lohse M.J., Kobilka B.K., Caron M.G., Lefkowitz R.J. The genomic clone G-21 which resembles a beta-adrenergic receptor sequence encodes the 5-HT1A receptor (англ.) // Nature : journal. — 1988. — Vol. 335, no. 6188. — P. 358—360. — doi:10.1038/335358a0. — PMID 3138543.
- Nakhai B., Nielsen D.A., Linnoila M., Goldman D. Two naturally occurring amino acid substitutions in the human 5-HT1A receptor: glycine 22 to serine 22 and isoleucine 28 to valine 28 (англ.) // Biochem. Biophys. Res. Commun. : journal. — 1995. — Vol. 210, no. 2. — P. 530—536. — doi:10.1006/bbrc.1995.1692. — PMID 7755630.
- Aune T.M., McGrath K.M., Sarr T., Bombara M.P., Kelley K.A. Expression of 5HT1a receptors on activated human T cells. Regulation of cyclic AMP levels and T cell proliferation by 5-hydroxytryptamine (англ.) // J. Immunol. : journal. — 1993. — Vol. 151, no. 3. — P. 1175—1183. — PMID 8393041.
- Parks C.L., Shenk T. The serotonin 1a receptor gene contains a TATA-less promoter that responds to MAZ and Sp1 (англ.) // J. Biol. Chem. : journal. — 1996. — Vol. 271, no. 8. — P. 4417—4430. — doi:10.1074/jbc.271.8.4417. — PMID 8626793.
- Stockmeier C.A., Shapiro L.A., Dilley G.E., Kolli T.N., Friedman L., Rajkowska G. Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression-postmortem evidence for decreased serotonin activity (англ.) // J. Neurosci. : journal. — 1998. — Vol. 18, no. 18. — P. 7394—7401. — PMID 9736659.
- Kawanishi Y., Harada S., Tachikawa H., Okubo T., Shiraishi H. Novel mutations in the promoter and coding region of the human 5-HT1A receptor gene and association analysis in schizophrenia (англ.) // Am. J. Med. Genet. : journal. — 1998. — Vol. 81, no. 5. — P. 434—439. — doi:10.1002/(SICI)1096-8628(19980907)81:5<434::AID-AJMG13>3.0.CO;2-D. — PMID 9754630.
- Salim K., Fenton T., Bacha J., Urien-Rodriguez H., Bonnert T., Skynner H.A., Watts E., Kerby J., Heald A., Beer M., McAllister G., Guest P.C. Oligomerization of G-protein-coupled receptors shown by selective co-immunoprecipitation (англ.) // J. Biol. Chem. : journal. — 2002. — Vol. 277, no. 18. — P. 15482—15485. — doi:10.1074/jbc.M201539200. — PMID 11854302.
- Panesar, Kiran 5-HT1A Receptors in Psychopharmacology. Web Article. Psychopharmacology Institute. Дата обращения: 11 апреля 2013.
Ссылки
- 5-HT1A. IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.