Таламус
Тала́мус, иногда — зри́тельные бугры (лат. thalamus; от др.-греч. θάλαμος «комната, камера, отсек») — отдел головного мозга, представляющий собой большую массу серого вещества, расположенную в верхней части таламической области промежуточного мозга хордовых животных, в том числе и человека. Впервые описан древнеримским врачом и анатомом Галеном. Таламус — это парная структура, состоящая из двух половинок, симметричных относительно межполушарной плоскости. Таламус находится глубже структур большого мозга, в частности коры или плаща. Под таламусом расположены структуры среднего мозга. Срединная (медиальная) поверхность обеих половинок таламуса одновременно является верхней боковой стенкой третьего желудочка головного мозга[2][3][4].
| Таламус | |
|---|---|
 Таламус человека на МРТ-снимке, отмечен стрелкой | |
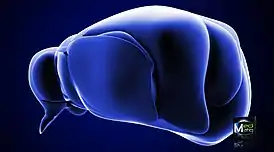 Таламус в передне-боковой проекции | |
| Часть | промежуточный мозг |
| Компоненты | см. список ядер таламуса |
| Артерия | см. артерии таламуса |
| Вена | см. вены таламуса |
| Каталоги | |
Таламус выполняет несколько важных физиологических функций. Он отвечает за передачу сенсорной и двигательной информации от органов чувств (кроме информации от органов обоняния) к соответствующим областям коры больших полушарий млекопитающих или плаща мозга низших хордовых. Таламус играет важную роль в регуляции уровня сознания, процессов сна и бодрствования, концентрации внимания[3].
Таламус — один из основных продуктов эмбрионального развития зародышевого промежуточного мозга. Этот факт был впервые установлен основоположником эмбриологии швейцарским анатомом Вильгельмом Гисом в 1893 году[4].
Ранее таламус считался структурой головного мозга, характерной только для хордовых. Ещё ранее его существование признавалось только у позвоночных. Учёные полагали, что таламус в принципе отсутствует у беспозвоночных, даже наиболее высокоорганизованных, таких как членистоногие. Однако в 2013 году в центральном нервном узле или головном мозге членистоногих обнаружена структура, гомологичная таламусу в головном мозге хордовых — так называемые «боковые вспомогательные дольки» (англ. lateral accessory lobes, LAL). У этих структур было выявлено сходство как в процессах эмбрионального развития и паттернах экспрессии генов, так и в анатомическом расположении в головном мозге. Сходство обнаружилось и в их физиологических функциях (сбор информации и передача её от различных сенсорных путей в более передне расположенные части головного мозга или центрального нервного узла)[5][6]. Таким образом, таламус, возможно, является эволюционно очень древней структурой мозга. Зачатки или предшественники таламуса, вероятно, возникли у общего предка хордовых и членистоногих около 550—600 млн лет назад[5][6].
Анатомия таламуса
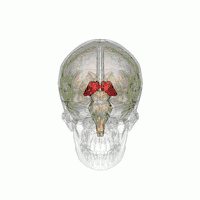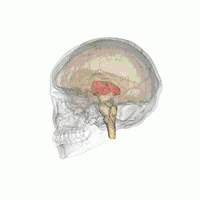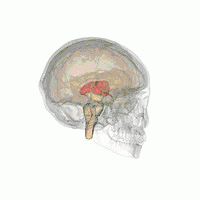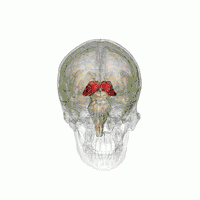
Таламус расположен вблизи центра мозга и входит в число структур таламической области промежуточного мозга. Он залегает под структурами большого мозга, но возвышается над структурами среднего мозга. Восходящие аксоны, исходящие из нейронов ядер таламуса, формируют пучки миелинизированных нервных волокон. Эти пучки нервных волокон обильно проецируются на различные области коры больших полушарий головного мозга во всех направлениях. Медиальная поверхность обеих половинок таламуса одновременно является верхней частью боковой стенки третьего желудочка головного мозга. Она соединена с соответствующей медиальной поверхностью противоположной половинки таламуса плоской полосой белого вещества. Эта полоса представляет собой пучок миелинизированных нервных волокон и называется межталамическим сращением, или промежуточной массой третьего желудочка, или срединной комиссурой (срединной спайкой) таламуса.
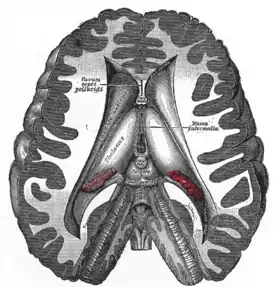
Ядра таламуса
| Ядра таламуса | |
|---|---|
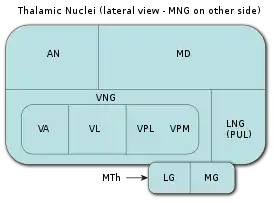 Список ядер таламуса: MNG = Срединная группа ядер AN = Передние ядра таламуса MD = Медиальное дорсальное ядро VNG = Вентральная группа ядер VA = Вентральное переднее ядро VL = Вентральное латеральное ядро VPL = Вентральное постеролатеральное ядро VPM = Вентральное постеромедиальное ядро LNG = Латеральная группа ядер PUL = Ядра подушки MTh = Метаталамус LG = Латеральное коленчатое тело MG = Медиальное коленчатое тело | |
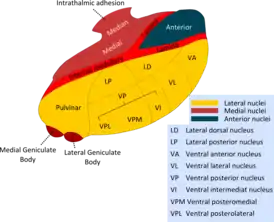 Ядра таламуса | |
| Каталоги | |
Таламус является частью более общей структуры — комплекса нейронных ядер, состоящего из четырёх частей: гипоталамуса, эпиталамуса, субталамуса (ранее называвшегося также преталамусом, периталамусом, вентральным таламусом или подбугорной областью), и собственно таламуса, который ранее называли (нередко называют для уточнения и в настоящее время) дорсальным таламусом[7].
Мозговые структуры, развивающиеся из разных частей эмбрионального промежуточного мозга приблизительно одновременно с собственно таламусом, включают в себя расположенный выше таламуса эпиталамус, состоящий из поводков мозга, эпифиза и прилегающих к ним ядер, а также расположенный ниже таламуса субталамус, содержащий так называемую неопределённую зону и таламическое ретикулярное ядро. Из-за их происхождения в ходе онтогенеза из разных, хотя и близко расположенных, частей зародышевого промежуточного мозга, эпиталамус и субталамус принято формально отличать от собственно таламуса (дорсального таламуса)[7].
Таламус состоит из нескольких отдельных областей серого вещества. Эти области представляют собой группы таламических ядер, разделённых областями белого вещества. Белое вещество, разделяющее и облегающее отдельные таламические ядра и группы ядер, представляет собой пучки миелинизированных нервных волокон. Кроме того, в таламусе выделяют также особые группы нейронов, отличающихся по своему гистологическому строению и биохимическому составу от остальной части таламуса, такие, как перивентрикулярное ядро, внутрипластинчатые ядра, так называемое ограниченное ядро и другие[4]. Эти отличающиеся по своей гистологической структуре и биохимическому составу от остальных ядер таламуса особенные ядра обычно группируют в так называемый аллоталамус, в противоположность «типичным» таламическим ядрам, которые группируют в так называемый изоталамус[8].
Ядра таламуса на основании особенностей их анатомо-гистологической структуры и цитоархитектоники можно подразделить на шесть групп: передние, медиальные, боковые, ретикулярные, внутрипластинчатые ядра и ядра средней линии. Тонкий Y-образный слой миелинизированных нервных волокон, так называемая внутренняя мозговая пластинка, разграничивает между собой переднюю, срединную и боковую группы таламических ядер. У человека передняя и срединная группы таламических ядер содержат всего по одному большому ядру, называемому соответственно передним дорсальным и медиальным дорсальным ядрами таламуса. Между тем боковая группа ядер таламуса человека подразделяется на дорсальный и вентральный уровни. Дорсальный уровень боковых ядер таламуса человека состоит из бокового дорсального, бокового заднего ядер и ядер подушки таламуса. Вентральный уровень боковых ядер таламуса человека, в свою очередь, состоит из вентрального переднего, вентрального бокового, вентрального задне-бокового и вентральных задних медиальных ядер[9].
Боковая группа ядер таламуса покрыта другим тонким слоем миелинизированных нервных волокон — так называемой наружной мозговой пластинкой. Между этим пучком нервных волокон и внутренней капсулой находится тонкий слой ГАМКергических нейронов, образующих ретикулярное ядро. Группа внутрипластинчатых ядер таламуса, среди которых самым крупным является так называемое центральное срединное ядро таламуса, как следует из её названия, располагается внутри Y-образной внутренней мозговой пластинки[9].
Наконец, последняя группа ядер таламуса, так называемые медиальные ядра, или «ядра средней линии», покрывает собой часть медиальной поверхности обеих половинок таламуса, и примыкает к верхней боковой стенке третьего желудочка головного мозга. Эти ядра средней линии (медиальные ядра) каждой из половинок таламуса могут соединяться между собой тонким и плоским серым пучком нервных волокон, так называемым межталамическим сращением. Выраженность межталамического сращения вариабельна не только у разных видов позвоночных, но даже у разных особей одного и того же вида (в том числе у разных людей), вплоть до её полного отсутствия. Отсутствие или слабая выраженность межталамического сращения практически не влияет на функциональность таламуса и является не патологией, а лишь анатомической особенностью (такой же, как наличие артерии Першерона)[9].
Функциональные различия ядер таламуса
Хотя анатомо-гистологическое подразделение ядер таламуса, безусловно, полезно, большее значение для понимания его функционирования имеет их функциональная группировка, проводимая на основании типа их связей с другими частями мозга и типа передаваемой по этим связям и обрабатываемой этими ядрами информации. С этой точки зрения, среди ядер таламуса функционально выделяют первичные сенсорные и моторные ретрансляционные ядра, ядра ретрансляции ассоциативной информации, диффузно-проекционные ядра, и тормозное, ауторегулирующее активность таламуса, ретикулярное ядро, состоящее, в отличие от остальных ядер таламуса, из ингибирующих ГАМКергических нейронов, а не из возбуждающих глутаматергических[9].
Первичные сенсорные и двигательные ретрансляционные ядра

Первичные сенсорные и двигательные ретрансляционные ядра таламуса передают информацию по своим проекциям в определённые области сенсорной и двигательной коры больших полушарий головного мозга, и получают от этих областей обратную связь. Эта обратная связь позволяет сенсорной и двигательной коре больших полушарий мозга регулировать активность соответствующих таламических ядер. Благодаря наличию системы отрицательной обратной связи сенсорные и двигательные области коры больших полушарий могут регулировать интенсивность получаемого этими областями коры входящего сенсорного сигнала от соответствующих ядер таламуса. Кроме того, система отрицательной обратной связи позволяет коре больших полушарий регулировать работу соответствующих фильтров в таламусе, степень и характер фильтрации ядрами таламуса входящего сигнала перед передачей его в кору[9]. Каждое конкретное первичное сенсорное или двигательное ретрансляционное ядро таламуса получает и обрабатывает информацию только от одной определённой системы органов чувств или от одной определённой части двигательной системы. Так, например, латеральное коленчатое ядро таламуса получает зрительный сигнал от зрительного тракта, производит его простейшую первичную обработку, фильтрует от помех и затем передаёт обработанный и отфильтрованный зрительный сигнал первичной зрительной коре, расположенной в затылочной доле коры больших полушарий мозга[9]. Нейроны первичных сенсорных ретрансляционных ядер таламуса для ряда систем органов чувств формируют топографически организованные проекции в определённые зоны коры больших полушарий. Например, для слуховой сенсорной системы определённые частоты звуковых сигналов отображаются соответствующим ядром таламуса на определённые участки первичной слуховой коры. То же самое верно для зрительной, соматосенсорной и висцеросенсорной систем, в которых отображается карта окружающего пространства или схема тела, соответственно. Аналогичную топографическую организацию проекций в двигательную кору, отображающую схему тела, проявляют нейроны первичных двигательных ретрансляционных ядер таламуса. Вместе с тем, некоторые первичные сенсорные и двигательные зоны коры больших полушарий получают информацию более чем от одного первичного сенсорного или двигательного ядра таламуса[9].
Ядра ретрансляции ассоциативной информации
В отличие от первичных сенсорных и двигательных ретрансляционных ядер таламуса, каждое из которых получает довольно простую и «сырую» (почти необработанную) информацию только от одного источника сенсорной или двигательной информации, и передаёт её в довольно ограниченные по своим размерам области первичной сенсорной или двигательной коры, ядра ретрансляции ассоциативной информации таламуса получают предварительно уже подвергшуюся высокой степени переработки информацию более чем из одного источника, ассоциируют одну информацию с другой, и передают её по своим проекциям в значительно более крупные по своим размерам области ассоциативной коры. Так, например, медиальное дорсальное ядро таламуса получает информацию одновременно от гипоталамуса и от миндалины, и связано двусторонними связями с префронтальной корой, а также с определёнными областями премоторной и височной коры[9].
Диффузно-проекционные ядра
В отличие от ретрансляционных ядер обоих типов (как первичных сенсорных и двигательных ретрансляторов, так и ядер ретрансляции ассоциативной информации), диффузно-проекционные ядра получают менее специфическую входящую информацию, но зато одновременно от множества различных источников. Они также широковещательно (диффузно) ретранслируют информацию по многообразным своим восходящим проекциям во многие различные области коры больших полушарий и в другие ядра таламуса, а по нисходящим проекциям — во множество различных подкорковых структур. Такой диффузный характер их проекций, широковещательный охват ретранслируемой по этим проекциям информации, а также неспецифичность получаемой ими входной информации, заставляет предполагать, что диффузно-проекционные ядра играют важную роль в регуляции общего уровня корковой и подкорковой возбудимости, уровня сознания, уровня активности и возбуждения ЦНС, концентрации внимания, и в регуляции смены состояний сна и бодрствования. И действительно, разрушение или повреждение этих ядер приводит в лёгких случаях к нарушениям концентрации внимания, к сонливости, а в более тяжёлых случаях — к летаргическому сну или перманентной коме. И наоборот, дегенеративные прионные изменения в них, такие, какие наблюдаются при фатальной семейной бессоннице — приводят к развитию упорной хронической, в пределе полной, бессонницы и в конечном итоге к смерти[9].
Ауторегуляторное ретикулярное ядро
Ретикулярное ядро таламуса уникально тем, что оно, в отличие от всех других ядер таламуса, содержит не возбуждающие глутаматергические, а, наоборот, тормозящие ГАМКергические нейроны. Ретикулярное ядро получает входящую информацию от ответвлений аксонов, которые взаимно соединяют другие таламические ядра с корой больших полушарий головного мозга. Каждый нейрон ретикулярного ядра затем направляет свой единственный исходящий аксон именно в то ядро таламуса, от которого он получает входящую информацию. Такая схема связности нейронов ретикулярного ядра с остальными ядрами таламуса позволяет предполагать, что нейроны ретикулярного ядра таламуса осуществляют постоянный контроль уровня активности других таламических ядер и, косвенно, иннервируемых ими корковых областей, получая копии входящей и исходящей информации, поступающей от этих ядер таламуса в кору больших полушарий и обратно, и затем использует эту информацию для регулирования уровня активности соответствующих таламических ядер[9].
Связи таламуса с другими структурами мозга
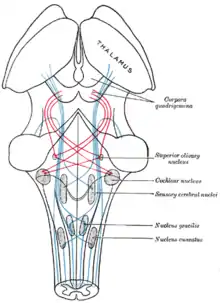
Сосцевидно-таламический путь
Таламус соединён с гиппокампом множеством двусторонних нервных связей, образующих так называемый сосцевидно-таламический путь, или сосцевидно-таламический тракт. В состав сосцевидно-таламического пути входят, в частности, сосцевидные тельца, а также свод мозга[10][11].
Корково-таламические и таламо-корковые пути
Таламус также соединён множеством двусторонних (восходящих и нисходящих) нервных связей с различными областями коры больших полушарий головного мозга. Эти связи образуют многообразные корково-таламические и таламо-корковые проекции[12]. Эти двусторонние связи образуют замкнутые кольцеобразно (нередко не напрямую, а с вовлечением базальных ядер) системы с отрицательной обратной связью, называемые таламо-корковыми или корково-таламо-корковыми системами[13]. Среди таламо-корковых систем особое значение имеет взаимодействие таламуса с теменными дольками коры больших полушарий, образующее так называемые таламо-теменные волокна[14].
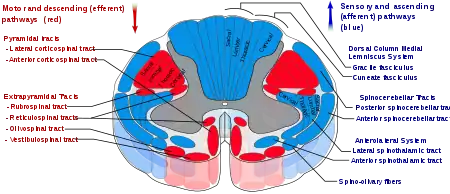
Спиноталамический путь
Восходящие нервные пути, соединяющие спинной мозг с таламусом, образуют спиноталамический путь, или спиноталамический тракт. В нём от спинного мозга к таламусу передаётся сенсорная информация о болевых, температурных и тактильных ощущениях, а также об ощущении зуда[15]. Спиноталамический путь подразделяется на две части: боковой, или латеральный, или дорсальный, спиноталамический путь[16], который передаёт информацию о болевых и температурных ощущениях, и передний, или вентральный, спиноталамический путь[17], который передаёт ощущения грубого прикосновения или сдавливания, надавливания[9].
В свою очередь, в боковом спиноталамическом пути выделяют эволюционно более молодой неоспиноталамический путь и более древний палеоспиноталамический путь. Первый состоит из большого количества тонких нервных волокон, которые быстро проводят болевые ощущения, а второй содержит меньшее количество более толстых и более медленно проводящих нервных волокон. Быстро проводящий неоспиноталамический путь играет большую роль в передаче в мозг хорошо локализованного ощущения острой боли непосредственно или вскоре после травмы, повреждения тканей, и в принятии организмом защитных мер, таких, например, как отдёргивание руки от горячего предмета. Более медленно проводящий палеоспиноталамический путь передаёт менее локализованную, более разлитую, тупую, давящую или сжимающую, реже жгучую или сверлящую хроническую боль, и играет большую роль в патогенезе различных хронических болевых синдромов[18][19].
Таламостриарные и таламо-оливарные пути
Таламус также тесно взаимодействует с полосатым телом, обмениваясь с ним информацией по так называемым таламостриарным волокнам[20]. С оливой таламус образует так называемый таламо-оливарный путь, он же центральный покрышечный путь[21].
Мозжечково-таламо-корковый путь
Мозжечково-таламо-корковый путь соединяет задние доли мозжечка, через зубчатое ядро и верхнюю ножку мозжечка, с вентральными ядрами таламуса и затем с двигательной и премоторной корой больших полушарий мозга[22].
Артериальное кровоснабжение и венозный отток от таламуса
При изучении кровоснабжения таламуса обращает на себя внимание то, что таламус интенсивнее кровоснабжается и имеет более развитую сеть коллатералей, чем некоторые другие близко расположенные структуры мозга. Это объясняется как важностью таламуса для функционирования мозга в целом, так и высокими метаболическими потребностями этой структуры, не намного уступающими метаболическим потребностям коры больших полушарий[23].
Артерии таламуса
Общепринятая международная анатомическая терминология для артерий таламуса до сих пор отсутствует. Одним из наиболее полных и подробных описаний артериальной сети таламуса является схема, выполненная Бенно Шлезингером ещё в 1976 году. Этот учёный предложил для упрощения классификации таламических артерий разделить их на две большие группы: парамедианные (парамедиальные, или срединные) артерии таламуса, они же центральные артерии таламуса, или таламо-проникающие артерии, и окружные, или огибающие (обходящие) артерии таламуса, они же поверхностные артерии таламуса[23].
Шлезингер указал, что таламо-коленчатые артерии относятся к промежуточному подтипу между упомянутыми двумя основными типами артерий таламуса. Они отходят из мест, типичных для мест отхождения огибающих артерий. Будучи короткими на протяжении субарахноидального пространства, у основания мозга эти артерии глубоко проникают в ткани промежуточного мозга под углом, характерным для таламо-проникающих артерий[23].
Парамедианные артерии таламуса по Шлезингеру
К основным парамедианным артериям таламуса Шлезингер отнёс туберо-таламические и глубокие межножковые артерии, а также одну из групп артерий подушки таламуса, а именно задние артерии подушки[23]. Все эти артерии являются ответвлениями задней мозговой артерии либо задней соединительной артерии[23][24].
Огибающие артерии таламуса по Шлезингеру
К основным представителям огибающих артерий таламуса Шлезингер отнёс передние и задние ворсинчатые артерии, а также нижние артерии подушки, и так называемые претектальные артерии, цингуло-таламические, спленио-таламические и латеральные мезэнцефальные артерии (латеральные артерии среднего мозга)[23].
Вариантная анатомия
У некоторых людей имеется так называемая «артерия Першерона» — редкая анатомическая вариация, при которой от одной из двух задних мозговых артерий, левой или правой, отходит единственная парамедианная таламическая артерия, кровоснабжающая парамедианные части обеих половинок таламуса. Обычно же от левой задней мозговой артерии отходит артерия, кровоснабжающая парамедианную часть левой половинки таламуса, а от правой — аналогичная артерия, кровоснабжающая парамедианную часть правой половинки[25][23].
Вены таламуса
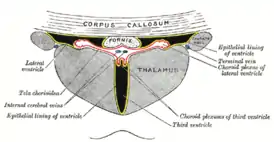
Вены таламуса собирают насыщенную карбгемоглобином и другими продуктами обмена веществ кровь. По этим венам она далее отекает в систему глубоких вен головного мозга[23].
Общепринятая международная анатомическая классификация вен таламуса также до сих пор отсутствует. Например, «Анатомия по Пирогову. Атлас анатомии человека» от 2011 года (авторы-составители В. В. Шилкин и В. И. Филимонов) из всех вен таламуса упоминает лишь самые крупные верхние вены таламуса, и только в виде однократного схематического изображения на картинке на стр. 351. В таблице вен головного мозга на стр. 347 эти вены — единственные из всего списка приведённых в нём вен мозга — помечены звёздочкой, означающей, что термин не включен в IAT (Международную анатомическую классификацию)[26].
Одним из наиболее полных и подробных описаний вен таламуса является описание Бенно Шлезингера (1976). Он предложил сгруппировать все найденные и описанные им вены таламуса в две большие группы — центральную группу и латеральную, или боковую, группу. Латеральная группа вен таламуса подразделялась на меньшую по количеству входящих в неё вен подгруппу латеральных верхних вен, и более крупную подгруппу латеральных нижних вен[23].
Шлезингер показал, что функционирование венозного кровеносного русла таламуса невозможно понять, если рассматривать только вены собственно таламуса. Необходимо рассматривать также вены соседних анатомических структур. Венозная кровь от некоторых частей таламуса отекает также во внеталамические вены. Зоны дренирования разных вен отчасти перекрываются. Вентральная задняя часть таламуса дренируется, наряду с прочими венами, также венами субталамо-мезэнцефалической группы, дренирующими расположенные рядом структуры — неопределённую зону, субталамус, чёрную субстанцию и красное ядро. Верхняя часть таламуса дренируется, наряду с прочими венами, также эпиталамическими венами, дренирующими структуры эпиталамуса. Прилегающие к стенке третьего желудочка мозга области таламуса дренируются, наряду с прочими венами, также маргинальными (пограничными) венами третьего желудочка, пролегающими непосредственно под его эпендимой[23].
Центральная группа вен таламуса по Шлезингеру
Согласно определению Шлезингера, центральные вены таламуса включают в себя вены, формирующиеся (берущие начало) в глубине таламического ядерного комплекса, и впадают либо в одну из малых вен Галена, либо в базальную вену (вену Розенталя)[23].
К центральной группе вен таламуса Шлезингер относил следующие вены:
- Фронтально-полярные вены таламуса собирают кровь от передних ядер таламуса и впадают во внутреннюю мозговую вену[23].
- Медиальные передние вены таламуса собирают кровь от медиальной передней части таламуса и впадают во внутреннюю мозговую вену[23].
- Самыми крупными венами таламуса являются вены, которые Шлезингер называл главными, или основными, или центромедиальными венами таламуса. Они собирают кровь от латеральных, вентральных и ретикулярной групп ядер таламуса, а также от ядер гипоталамуса, и впадают во внутреннюю мозговую вену[23].
- Таламо-коленчатые вены, которые Шлезингер иногда также называл коленчато-таламическими, выходят у заднего конца (заднего полюса) таламуса, собирают кровь преимущественно от метаталамуса (состоящего из латерального и медиального коленчатых тел) и впадают либо в базальную вену (вену Розенталя), либо в вены преддверия третьего желудочка головного мозга[23].
- К центральной группе вен таламуса он относил также вены подушки таламуса, заслуживающие отдельного подраздела.
Вены подушки таламуса
Вены подушки таламуса, а именно нижние и срединные (или медиальные) вены подушки таламуса, как следует из их названия, собирают кровь от ядер подушки таламуса. Нижняя вена подушки таламуса впадает в базальную вену. Срединная вена подушки таламуса впадает во внутреннюю мозговую вену[23].
Латеральная группа вен таламуса по Шлезингеру
Согласно определению Шлезингера, к латеральной группе вен таламуса относятся вены, которые формируются (берут своё начало) в латеральной области таламуса, или, иначе говоря, в таламо-капсулярной области, то есть в той области таламуса, которая прилегает к внутренней капсуле[23]. Подгруппа латеральных верхних вен таламуса впадает в верхнюю таламостриарную вену. Подгруппа латеральных нижних вен таламуса впадает в базальную вену (вену Розенталя) или в один из её межножковых притоков[23].
К латеральной группе вен таламуса Шлезингер относил следующие вены:
- Латеральная верхняя подгруппа
- Парные латеродорсальные вены таламуса собирают кровь от латеродорсальных ядер соответствующих половинок таламуса, и впадают в верхнюю таламостриарную вену[23].
- Латеральная нижняя подгруппа
- Парные латерокаудальные и латеровентральные вены таламуса собирают кровь, соответственно, от каудальной и вентральной частей латеральной группы ядер соответствующей половинки таламуса, и впадают в базальную вену[23].
Физиологические функции таламуса
Таламус выполняет множество физиологических функций. В частности, ранее считалось, что таламус является всего лишь центральным «реле» или ретрансляторной станцией, которая просто передаёт различные сенсорные и двигательные сигналы (кроме сигналов от органов обоняния) в кору больших полушарий головного мозга. Более поздние исследования показали, что функции таламуса гораздо сложнее, многообразнее и селективнее. Они не сводятся лишь к простой ретрансляции информации от нижележащих подкорковых областей и структур головного мозга к коре больших полушарий. Таламус также производит некую её первичную обработку и фильтрацию. Каждое из ядер таламуса, специализирующееся на первичной ретрансляции информации от органов чувств того или иного типа в кору больших полушарий, получает сильные обратные связи от соответствующей зоны коры больших полушарий, регулирующие активность этого ядра и степень фильтрации им входящего потока информации[9][27][28].
Нервные пути, передающие информацию от таких систем органов чувств, как зрение, слух, ощущение вкуса, устроены следующим образом: информация от чувствительных рецепторов (будь то палочки и колбочки сетчатки глаза, вкусовые рецепторы сосочков языка, или волосковые клетки улитки) поступает по нервным волокнам соответствующего нерва (зрительного, слухового или вкусового) сначала в ядро этого нерва, расположенное в определённой области промежуточного мозга. Затем информация по волокнам соответствующего нервного пути передаётся в структуры, расположенные в среднем мозге и традиционно называемые «первичными анализаторами ствола мозга» для соответствующей системы органов чувств. Например, для системы зрения таким «первичным стволовым анализатором» являются верхние холмики четверохолмия. А для системы восприятия звука, то есть для слуха, таким «первичным стволовым анализатором» являются нижние холмики четверохолмия. Эти первичные стволовые анализаторы проводят простейшую обработку и интеграцию сенсорной информации, поступающей от соответствующего органа чувств. На следующем этапе обработанная сенсорная информация поступает от первичного стволового анализатора в соответствующее специализированное ядро таламуса. Для зрения таким ядром является латеральное коленчатое тело, для слуха — медиальное коленчатое тело. А для ощущения вкуса — парвоцеллюлярная (мелкоклеточная) часть вентрального постеромедиального ядра, называемая ещё иногда «вкусовым ядром таламуса». Эти ядра производят уже более сложную обработку и фильтрацию поступающей сенсорной информации, а затем передают обработанную и отфильтрованную информацию в соответствующую первичную область сенсорной коры больших полушарий (зрительную, слуховую и т. д.), а также в соответствующие вторичные сенсорно-ассоциативные области коры. Там происходит окончательная обработка и осознание поступившей информации[9].
Роль таламуса в обработке сенсорной, моторной, висцеросенсорной и соматосенсорной информации
Различные ядра и области таламуса выполняют различные специфические функции. В частности, это относится ко многим системам органов чувств, за исключением обонятельной системы, таким, как слуховая, зрительная, соматосенсорная, висцеросенсорная системы, система ощущения вкуса. Для каждой из этих систем существуют свои специализированные ядра таламуса, выполняющие функцию центральной релейной или ретрансляторной станции именно для этой системы. Изолированные локальные поражения этих ядер таламуса вызывают специфические нейросенсорные нарушения или дефициты в системе восприятия информации от соответствующих органов чувств[9].
Так, например, для зрительной системы, вся входящая информация от сетчатки глаз передаётся через верхние холмики четверохолмия в латеральное коленчатое тело, а уже оно, в свою очередь, направляет эту информацию, после её первичной обработки, в зрительную кору в затылочных долях коры больших полушарий мозга. Аналогично, медиальное коленчатое тело является центральным реле или ретрансляторной станцией для всей звуковой (слуховой) информации. Это ядро передаёт всю поступающую от нижних холмиков четверохолмия слуховую и звуковую информацию, после её первичной обработки, в первичную слуховую кору. В свою очередь, вентральное заднее ядро таламуса является центральным реле для всей соматосенсорной, тактильной, проприоцептивной и ноцицептивной (болевой) информации, поступающей от спинного мозга, и направляет её в первичную соматосенсорную кору. Парвоцеллюлярная часть вентрального постеромедиального ядра является аналогичным центральным реле для всей вкусовой информации[9].
Роль таламуса в регуляции уровня сознания, цикла сон-бодрствование, концентрации внимания
Таламус играет важную роль в регуляции уровня сознания, общего уровня возбуждения ЦНС, в регуляции концентрации внимания, смены состояний сна и бодрствования[29]. Ядра таламуса имеют множество сильных двусторонних взаимных связей с корой больших полушарий головного мозга. Эти связи образуют кругообразно замкнутые таламо-корково-таламические и корково-таламо-корковые цепи, которые, как считается, связаны с регулированием уровня сознания, уровня возбуждения ЦНС, концентрации внимания, смены состояний сна и бодрствования. Повреждение таламуса может привести к летаргическому сну или перманентной (постоянной) коме или, наоборот, к упорной бессоннице[9].
Роль таламуса в работе двигательной системы и систем языка и речи
Помимо передачи сенсорной, соматосенсорной, висцеросенсорной и двигательной информации в кору больших полушарий, таламус играет важную роль в интеграции работы и поддержании функционирования двигательной системы и системы языка и речи. Большая часть нейросхем таламуса, задействованных в регуляции работы этих сложных систем, вовлекает не одно, а несколько таламических ядер или групп ядер[9].
Таламус участвует во множестве нейронных информационных цепей, необходимых для управления двигательной подсистемой, и выполняет роль ключевого подкоркового центра регуляции движений, являясь подкорковым двигательным центром «высшего порядка» по отношению к мозжечку и базальным ядрам[30]. Благодаря исследованиям анатомии головного мозга приматов[31], удалось выяснить характер многочисленных взаимосвязей ядер таламуса с мозжечком, базальными ядрами и двигательной корой. Это позволило выдвинуть предположение, что таламус выполняет функцию ключевого центра связи и передачи двигательной информации по специализированным каналам от базальных ядер и мозжечка к двигательной коре[32][33]. На обезьянах (макаках) было показано, что ядра таламуса участвуют в осуществлении антисаккадных движений глаз[34][35][36].
Роль таламуса в регуляции функций нижележащих и более передне расположенных структур системы базальных ядер, в частности, нигростриарной и стрио-паллидарной систем, участвующих в двигательных актах, хотя и общепризнана, но всё ещё относительно плохо изучена. Роль таламуса в регуляции вестибулярной функции (функции поддержания равновесия тела) и функций ориентировки четверохолмия часто недооценивается или игнорируется, и также пока ещё плохо изучена[9].
Роль таламуса в работе систем памяти и эмоций (гиппокампа и лимбики)
Передние ядра таламуса тесно функционально связаны с гиппокампом и структурами лимбической системы[37], и часто рассматриваются как составная часть расширенной системы гиппокампа и лимбики[38]. В этой системе гиппокамп участвует в реализации функций памяти: запоминании, хранении и последующем воспроизведении информации о том или ином событии, в том числе о его привязке к пространству, времени, и о связанных с ним запахах, звуках, зрительных образах, вкусовых и прочих сенсорных ощущениях, а также о сопровождавших событие эмоциях и о его эмоциональной оценке постфактум. Лимбическая система (в частности, миндалина (амигдала)) предоставляет гиппокампу для последующего запоминания формируемую ею эмоциональную оценку события как положительного или отрицательного, как во время самого события, так и после. Передние ядра таламуса предоставляют гиппокампу для последующего запоминания интегрированную, мультимодальную сенсорную и двигательную информацию о событии (о сопровождавших это событие зрительных образах, звуках, вкусовых, соматосенсорных и прочих ощущениях, в привязке ко времени и месту в пространстве). Таким образом, взаимодействие этих трёх структур — передних ядер таламуса, эмоциональных центров лимбической системы и системы памяти в гиппокампе — играет решающую роль в формировании целостного воспоминания о картине того или иного события. Сюда относятся и все сопутствующие событию пространственно-временные атрибуты (метки), звуки, образы и другие сенсорные ощущения, а также его эмоциональная оценка. Тесные функциональные связи передних ядер таламуса с гиппокампом и структурами лимбической системы играют ключевую роль в формировании как человеческой эпизодической памяти, так и памяти событий у грызунов и других млекопитающих[39][40].
Существует гипотеза о том, что связи определённых областей таламуса с определёнными областями мезио-темпоральной части (средней части височной доли) коры больших полушарий играют важную роль в дифференциации функционирования памяти пассивных воспоминаний и памяти узнавания знакомых мест, предметов и др., как у человека, так и у других млекопитающих[10].
Эволюция таламуса в ходе филогенеза
В таламусе выделяют эволюционно более древнюю часть — так называемый палеоталамус[41], и эволюционно более молодую часть — неоталамус[42].
При сохранении общего эволюционно-консервативного плана строения таламуса, конкретные детали строения, степень сложности, общее количество ядер и функциональность таламуса очень сильно различаются у разных видов хордовых, стоящих на разных ступенях эволюционной лестницы.
Таламус анамниотических хордовых
Таламус у круглоротых рыб достаточно хорошо различим, в нём заметны дорсальная и вентральная части («дорсальный таламус», или собственно таламус, и «вентральный таламус», или субталамус). Однако в обеих частях таламуса у круглоротых всего по нескольку ядер. У костистых рыб таламус организован уже более сложно, в нём имеется несколько хорошо дифференцированных ядерных групп, он имеет больше связей с другими областями мозга рыбы. Большинство ядер таламуса костистых рыб занимаются обработкой соматосенсорной и зрительной информации, проекции их диффузны, слабо топографически локализованы. У земноводных таламус, особенно его эволюционно более молодая дорсальная часть («собственно таламус») уже значительно больше по размерам, чем у рыб. Клеточный состав разных групп ядер таламуса земноводных сильнее различается, чем у рыб, но менее дифференцирован, чем у пресмыкающихся. Большая часть нейронов таламуса земноводных занимается обработкой зрительной информации, меньшая — соматической, слуховой или вестибулярной информации[4].
Таламус низших амниот (пресмыкающихся и птиц)
В таламусе пресмыкающихся можно найти чётко гомологичные ядрам таламуса млекопитающих группы ядер, с типичным именно для этих групп ядер гистологическим строением и типичным паттерном связей с другими частями мозга. У наиболее изученных модельных видов пресмыкающихся в дорсальном таламусе можно выделить 9 ядер, в вентральном таламусе (субталамусе) — 7 ядер. Входящие и исходящие связи таламуса с другими частями мозга у пресмыкающихся организованы значительно сложнее, чем у рыб и у земноводных. Боковые ядра таламуса пресмыкающихся принимают входящую информацию от ретиноталамического тракта (то есть от волокон сетчатки). Центромедиальные ядра таламуса пресмыкающихся занимаются обработкой и интеграцией зрительной и соматосенсорной информации, а вентрокаудальные — обработкой и интеграцией зрительной и слуховой информации. То есть те и другие занимаются мультисенсорной интеграцией и ассоциацией, что не свойственно таламусу рыб. Таламус птиц устроен не намного сложнее таламуса пресмыкающихся. Строение таламуса птиц, количество и функциональное подразделение его ядер у них в целом сходно с таковыми у пресмыкающихся[4].
Таламус высших амниот (млекопитающих и человека)
Наибольшего развития таламус достигает у млекопитающих. У хищников он более развит, чем у грызунов или травоядных животных, а наибольшего развития достигает у высших приматов и особенно у человека. Именно у млекопитающих дорсальный таламус («собственно таламус») стал основной релейно-ретрансляторной станцией, основным центром связи нижележащих отделов мозга с новой корой, звеном, через который проходит и в котором ассоциируется, фильтруется и обрабатывается вся сенсорная, висцеросенсорная, соматосенсорная и двигательная информация, за исключением информации от органов обоняния. Именно у млекопитающих сформировалось множество замкнутых по принципу кольца двусторонних связей таламуса с новой корой, то есть таламо-корково-таламических и корково-таламо-корковых систем. Наиболее интенсивно в ходе эволюции у млекопитающих развивались так называемые ассоциативные ядра высшего порядка (или ассоциативные ядра верхнего уровня), занимающие дорсальную часть таламуса. В эти ядра поступает меньшее количество входящих нервных волокон, чем в ассоциативные ядра низшего порядка, но они теснее связаны с ассоциативными областями коры. Именно ядра верхнего уровня, участвующие в образовании таламо-корковых ассоциативных систем, достигают наибольшего развития у высших приматов и человека. С работой этих ядер, наряду с работой новой коры, связывают возникновение у приматов зачатков разума и самосознания[4].
Гомология таламуса хордовых и LAL членистоногих
Как уже упоминалось, в центральном нервном узле или головном мозге членистоногих обнаружена структура, гомологичная таламусу в головном мозге хордовых, как с точки зрения сходства процессов эмбрионального развития и паттернов экспрессии генов, так и с точки зрения сходства анатомического расположения в головном мозге, и с точки зрения сходства выполняемых физиологических функций (сбор информации и ретрансляция различных сенсорных путей в более передне расположенные части головного мозга или центрального нервного узла) — так называемые «боковые вспомогательные дольки» (англ. lateral accessory lobes, LAL)[5][6].
Обнаружение этой гомологии таламуса хордовых и LAL членистоногих оставляет учёным две возможности для её объяснения. Первая гипотеза состоит в том, что хотя бы зачаточные, примитивные структуры мозга, подобные таламусу у хордовых и LAL членистоногих, уже имелись у гипотетического последнего общего предка хордовых и членистоногих — у так называемой «урбилатерии». И были унаследованы этими двумя ветвями эволюционного древа от этого гипотетического последнего общего предка. И далее развивались уже независимо. В этом случае можно вести речь об истинной гомологии этих структур. Согласно этой теории, момент возникновения зачаточного таламуса у общего предка хордовых и членистоногих датируется 550—600 млн лет назад, когда этот гипотетический последний общий предок, предположительно, жил[5][6].
Вторая гипотеза состоит в том, что у последнего общего предка хордовых и членистоногих — урбилатерии — этих структур мозга, даже в самом примитивном виде, вообще ещё не было. Согласно этой гипотезе, таламус хордовых и LAL членистоногих возникли у обеих ветвей эволюционного древа параллельно и совершенно независимо, а не были унаследованы от последнего общего предка. Это может объясняться, например, параллельной или конвергентной эволюцией. Согласно этому предположению, сходные условия жизни ранних морских хордовых и ранних морских членистоногих (ракообразных) и сходное давление эволюционного отбора на тех и на других привело к независимому появлению в мозге тех и других сходных структур, решающих сходные задачи — у хордовых — таламуса, а у членистоногих — LAL. В этом случае речь идёт не об истинной гомологии, а о гомоплазии. Эволюционный возраст таламуса, согласно этой теории, несколько меньше и соответствует моменту появления собственно хордовых[6].
Среди учёных имеются сторонники как той, так и другой точки зрения на эволюционную историю таламуса хордовых и LAL членистоногих и на причины их сходства. Однако, даже если хордовые и членистоногие развили сходные структуры мозга — у первых таламус, а у вторых LAL — полностью независимо, то их последний общий предок уже должен был обладать так называемым «эмбриональным потенциалом» для того, чтобы его потомки, образовавшие разные ветви эволюционного древа, могли независимо развить подобные структуры. Эмбриональный потенциал — это наличие в геноме живого существа генов, которые изначально могли выполнять какие-то другие функции, но позже, в процессе эволюции, были востребованы («рекрутированы») потомками этого живого существа для образования во время эмбриогенеза каких-то новых анатомических структур, для усложнения строения тела. Кроме того, эмбриональный потенциал также подразумевает гибкость уже имеющейся у этого живого существа-предка программы эмбриогенеза, её совместимость с таким расширением у потомков, без поломки других этапов эмбриогенеза. В данном случае речь идёт о том, что у гипотетического последнего общего предка хордовых и членистоногих — урбилатерии — уже должны были иметься гены, ныне управляющие сегментацией мозга и эмбриональным развитием зачатков таламуса у хордовых и зачатков LAL у членистоногих. Те самые гены, которые учёные идентифицировали как гомологичные у хордовых и у членистоногих и на основании обнаружения которых предположили гомологичность таламуса хордовых и LAL членистоногих и общую их эволюционную историю. Молекулярные часы по этим генам, опять-таки, приводят к оценке времени возникновения у последнего общего предка хордовых и членистоногих эмбрионального потенциала для последующего независимого развития таламуса у хордовых и LAL у членистоногих в 550—600 млн лет[5][6].
Эмбриональное развитие таламуса

Зародышевый таламический комплекс состоит из субталамуса (вентрального таламуса), среднедиэнцефалического организатора (который позже, в процессе эмбрионального развития таламуса, образует так называемый ограниченный внутриталамический пояс), и собственно таламуса (дорсального таламуса)[43][44]. Процесс эмбрионального развития таламуса подразделяется на три основных этапа: формирование первичных доменов таламуса, образование среднедиэнцефалического организатора, и последующее созревание таламуса с формированием его ядерной и зональной организации[45].
Таламус является самой крупной мозговой структурой, происходящей из зародышевого промежуточного мозга, расположенной между нижележащими структурами среднего мозга и вышележащими структурами большого мозга, в частности, корой больших полушарий мозга[45].
Раннее развитие мозга
У зародыша человека уже на стадии Карнеги 9, то есть ещё до завершения нейруляции и формирования первичной нервной трубки, ещё на стадии загибания внутрь концов первичной нервной пластинки, в ней становятся различимы отдельные нейромеры, в том числе самый ростральный (самый передне расположенный) прозомер P, зачаток будущего переднего мозга. Позднее этот зачаток становится передним первичным мозговым пузырём. Затем этот первичный мозговой пузырь делится на два вторичных мозговых пузыря — конечный мозг и промежуточный мозг. Ещё чуть позже в развивающемся промежуточном мозге зародыша образуются два вторичных прозомера — D1 и D2[46][47]. Из прозомера D2, собственно, и развиваются в дальнейшем таламус, а также эпиталамус и субталамус, в то время как из прозомера D1 развивается в дальнейшем гипоталамус[48].
Данные, полученные в результате изучения процессов эмбрионального развития головного мозга у различных модельных позвоночных организмов, позволяют выдвинуть гипотезу о том, что для правильного развития эмбрионального таламического комплекса решающее значение имеет взаимодействие между двумя семействами факторов транскрипции, Fez-подобными белками FEZ1 и Fez2, и Otx-подобными белками Otx1 и OTX2. Fez-подобные факторы транскрипции FEZF1 и FEZF2 селективно экспрессируются в процессе эмбрионального развития головного мозга клетками именно в области субталамуса, и функциональные эксперименты с выключением соответствующего гена показывают, что экспрессия Fez-подобных белков FEZF1 и FEZF2 необходима для правильного развития субталамуса[49][50]. Позади развивающегося под влиянием Fez-подобных белков FEZF1 и FEZF2 субталамуса, области экспрессии белков Otx1 и OTX2 примыкают и упираются в область экспрессии Fez-подобных белков FEZF1 и FEZF2 (то есть в будущий субталамус). Эти два белка, Otx1 и OTX2, необходимы для правильного развития таламуса[51][52].
Формирование первичных доменов таламуса
В процессе раннего эмбрионального развития таламуса формируются два его первичных домена, каудальный домен (так называемый домен TH-C) и ростральный домен (так называемый домен TH-R). Каудальный домен эмбрионального таламуса служит источником клеток-предшественников для развития всех глутаматергических нейронов таламуса взрослых особей хордовых животных, в то время как ростральный домен эмбрионального таламуса служит источником клеток-предшественников для развития всех ГАМКергических нейронов таламуса взрослых особей хордовых животных[53].
Формирование среднедиэнцефалического организатора
В области стыка между доменами экспрессии семейств факторов транскрипции Fez-подобных белков FEZF1 и FEZF2 с одной стороны, и Otx1 и OTX2 с другой стороны, то есть на границе между будущим субталамусом, и будущим таламусом, в эмбриональном таламическом комплексе формируется так называемая среднедиэнцефалическая организующая структура, или среднедиэнцефалический организатор. Среднедиэнцефалический организатор является главным организатором всего последующего процесса эмбрионального развития таламуса и субталамуса, рассылающим необходимые для правильной дифференцировки клеток ядер таламуса и субталамуса межклеточные сигналы. Отсутствие среднедиэнцефалического организатора приводит к отсутствию таламуса и нередко также субталамуса в развивающемся головном мозге эмбриона. Сам среднедиэнцефалический организатор созревает в процессе эмбрионального развития таламического комплекса в направлении от более вентральных его частей, созревающих раньше, к более дорсальным, которые созревают позже. Белки, принадлежащие к семействам SHH и Wnt, являются главными регуляторными и дифференцировочными сигналами, испускаемыми среднедиэнцефалическим организатором[45].
Кроме своей функции организатора, управляющего всем процессом дальнейшего эмбрионального развития таламуса и субталамуса, среднедиэнцефалический организатор впоследствии созревает в особую гистологическую структуру внутри таламуса, так называемую ограниченную внутриталамическую зону[45].
Созревание и зональная организация таламуса
_section_description.JPG.webp)
Сразу после своего формирования среднедиэнцефалический организатор начинает выполнять роль главного организатора всего дальнейшего процесса эмбрионального развития таламуса и субталамуса. Эту роль он выполняет, выделяя такие сигнальные молекулы, как SHH и Wnt[54]. У мышей и других млекопитающих не удалось напрямую выяснить функциональную роль сигнальных молекул белка SHH, выделяемых среднедиэнцефалическим организатором, в управлении процессом дальнейшего эмбрионального развития таламуса и субталамуса. Причиной является то, что искусственно внесённая генетическая мутация, которая приводит к отсутствию функционального белка SHH, приводит к полному отсутствию у развивающегося зародыша зачатков не только таламического комплекса, но и всего промежуточного мозга[55].
Тем не менее, исследования на развивающихся эмбрионах курицы показали, что экспрессия среднедиэнцефалическим организатором сигнального белка SHH является одновременно и необходимым, и достаточным условием для последующей индукции экспрессии генов, управляющих дифференцировкой клеток таламуса и субталамуса, и, соответственно, для их правильного развития[56]. Исследования на другом модельном организме, рыбках данио-рерио, показали, что экспрессия двух генов семейства SHH, так называемых SHH-a и SHH-b (ранее также известного как twhh), определяет границы зоны среднедиэнцефалического организатора, и что сигнальные молекулы SHH необходимы и достаточны для начальной индукции молекулярной дифференцировки клеток будущего таламуса и субталамуса, но не обязательны для их дальнейшего поддержания и созревания. Кроме того, исследования на рыбках данио-рерио показали, что сигнальные молекулы SHH, исходящие из среднедиэнцефалического организатора, необходимы и достаточны для индукции дальнейшей дифференцировки и созревания как таламуса, так и субталамуса. В то же время сигналы SHH, исходящие из более вентральных по отношению к развивающимся таламусу и субталамусу областей мозга, большого значения для развития этих структур не имеют, и отсутствие вентрально исходящих сигналов SHH не приводит к нарушению развития таламуса и/или субталамуса, в отличие от сигналов SHH, исходящих от среднедиэнцефалического организатора[57].
Воздействие градиента экспрессии белка SHH, продуцируемого среднедиэнцефалическим организатором, приводит к дифференцировке нейронов будущего таламуса и субталамуса. Градиент экспрессии белка SHH, продуцируемого среднедиэнцефалическим организатором, вызывает формирование волны градиента экспрессии белка пронейрального гена нейрогенина-1, распространяющейся в направлении от задней части к передней, в основном (каудальном) домене таламического зачатка, и одновременно — формирование волны градиента экспрессии белка ASCL1 (ранее известного как Mash1) в оставшейся узкой полоске рострально расположенных клеток таламического зачатка, непосредственно прилегающей к среднедиэнцефалическому организатору (то есть в ростральном домене таламического зачатка), и в субталамусе[58][59].
Формирование этих специфических зональных градиентов экспрессии тех или иных пронейральных белков приводит к дальнейшей дифференцировке глутаматергических «релейно-ретрансляторных» нейронов из расположенных в каудальном домене таламического зачатка клеток-предшественников, содержащих белок нейрогенин-1, и к дифференцировке ГАМКергических ингибирующих нейронов из расположенных в ростральном домене таламического зачатка, непосредственно прилегающем к среднедиэнцефалическому организатору, и в субталамусе клеток-предшественников, содержащих белок ASCL1. У рыбок данио-рерио выбор одного из этих двух альтернативных путей дифференцировки для каждой конкретной клетки-предшественника в той или иной зоне зачаточного таламического комплекса управляется динамической экспрессией белка Her6, являющегося гомологом белка HES1 человека. Экспрессия этого фактора транскрипции, относящегося к семейству «волосообразных» белков bHLH, приводит к подавлению экспрессии гена нейрогенина-1, однако необходима для поддержания и усиления экспрессии белка ASCL1. В процессе дальнейшего эмбрионального развития таламического зачатка экспрессия белка Her6 и, соответственно, связанное с ним подавление экспрессии белка нейрогенина-1 и усиление экспрессии белка ASCL1 постепенно исчезает в каудальном домене таламического зачатка, в то время как в субталамусе и в узкой полоске рострально расположенных таламических клеток, примыкающей к среднедиэнцефалическому организатору, экспрессия белка Her6 и, соответственно, подавление экспрессии белка нейрогенина-1 и усиление экспрессии ASCL1 нарастает. Это делает каудально-ростральный градиент экспрессии нейрогенина-1/ASCL1 более выраженным, границы доменов более чёткими, и способствует завершению созревания и дифференцировки клеток таламуса и субталамуса. Исследования на развивающихся эмбрионах курицы и мыши показали, что блокада сигнального пути белка SHH в этот период эмбрионального развития приводит к полному отсутствию рострального домена таламического зачатка и к значительному уменьшению размеров каудального домена таламического зачатка. Ростральный домен таламического зачатка даёт начало ГАМКергическим ингибирующим нейронам таламуса, расположенным в основном в ретикулярном ядре таламуса взрослых животных, в то время как каудальный домен таламического зачатка даёт начало глутаматергическим «релейно-ретрансляторным» нейронам, составляющим основную часть клеток таламуса. В дальнейшем происходит дифференцировка этих нейронов с образованием отдельных таламических ядер и групп ядер[45].
Было показано, что у человека часто встречающаяся генетическая вариация в области промотора гена белка транспортера серотонина (SERT), а именно, обладание длинной (SERT-long) или короткой (SERT-short) аллелями этого гена (гена 5-HTTLPR), влияет как на эмбриональное, так и на последующее (постэмбриональное) развитие и созревание определённых областей таламуса и на их конечный размер у взрослых. Люди, у которых имеется две «коротких» аллели гена 5-HTTLPR (SERT-ss), имеют больше нейронов в ядрах подушки таламуса и более крупный размер этих ядер, а также, возможно, больше нейронов и более крупный размер лимбических ядер таламуса (ядер, поддерживающих связь с эмоциональными центрами лимбической системы), по сравнению с гетерозиготами по этому гену или обладателями двух «длинных» аллелей гена 5-HTTLPR. Увеличение размеров этих структур таламуса у таких людей предполагается в качестве части анатомического объяснения того, почему люди, у которых имеется две «коротких» аллели гена 5-HTTLPR, в большей мере, чем люди, гетерозиготные по этому гену или обладающие двумя «длинными» аллелями гена 5-HTTLPR, предрасположены к таким психическим расстройствам, как большое депрессивное расстройство, посттравматическое стрессовое расстройство (ПТСР), а также к суицидальным тенденциям и попыткам[60].
Заболевания таламуса
Повреждение некоторых ядер таламуса, например, возникающее в результате церебрососудистой катастрофы (ишемического или геморрагического инсульта), или в результате метастазирования злокачественной опухоли в таламус, может привести к развитию так называемого «синдрома Дежерина-Русси», или синдрома таламической боли, синдрома таламической каузалгии — синдрома, характеризующегося чрезвычайно интенсивными, нелокализованными или плохо локализованными, ощущениями жжения или жгучей боли (откуда, собственно, и название «таламическая каузалгия», от «каустик» — жгучий) в одной или обеих половинах тела, а также перепадами настроения или депрессией[61]. Синдром этот назван в честь французских неврологов Дежерина и Русси, впервые описавших его в 1906 году под названием «таламический синдром» (фр. le syndrome talamique)[61][62].
Однако таламическим синдромом в настоящее время нередко называют не только синдром таламической боли, как в исходном определении Дежерина и Русси, но любые клинические синдромы, связанные с поражением таламуса. Так, в частности, одно- или, чаще, двусторонняя ишемия области, кровоснабжаемой парамедианной артерией, может вызвать серьёзные проблемы с регуляцией двигательных, глазодвигательных и речевых функций, вплоть до развития атаксии или акинетического мутизма (немоты и неподвижности), или паралича взора[63]. Таламокортикальная дизритмия может вызывать нарушения цикла сна и бодрствования, а также разнообразные другие нарушения, в зависимости от характера затронутых дизритмией таламокортикальных цепей[64]. Окклюзия артерии Першерона может привести к двустороннему инфаркту таламуса[65].
Корсаковский синдром (синдром ретроградной амнезии) связывают с повреждением или нарушением функции таламо-гиппокампальных связей, в частности, сосцевидно-таламического пути, сосцевидных телец или самого таламуса, и его даже иногда называют «таламической амнезией» или «таламической деменцией»[10].
Фатальная семейная бессонница — это редкое наследственное прионное заболевание, при котором происходят постепенно нарастающие дегенеративные изменения в нейронах таламуса. В результате этого пациент постепенно утрачивает способность уснуть, и в конечном итоге переходит в состояние полной бессонницы, что неизбежно приводит к смерти[66]. Напротив, повреждение таламуса в результате травмы, нейроинфекции, ишемического или геморрагического инсульта, или его поражение опухолью, может привести к летаргическому сну или коме[9].
Синдром дефицита внимания с гиперактивностью связывают с естественной, физиологической возрастной функциональной незрелостью таламуса у детей в целом, и с патологическим запаздыванием созревания таламуса у детей с СДВГ по сравнению с контрольной группой здоровых детей, в особенности[67]. При синдроме Клейне — Левина отмечается гиперактивность определённых ядер таламуса на фМРТ. Этот факт предложен к использованию для облегчения диагностики этого редкого синдрома, и как один из способов приблизиться к пониманию нейробиологии данного синдрома[68]. При нарколепсии, которую связывают с функциональной недостаточностью орексинергической системы мозга (например, вследствие аутоиммунного разрушения орексинергических нейронов гипоталамуса) наблюдается дизрегуляция активности эмоциональных центров лимбической системы и системы вознаграждения, в частности передних ядер таламуса, миндалины (амигдалы), прилежащего ядра, гиппокампа, вентральной области покрышки и некоторых других областей среднего мозга, а также префронтальной и височной коры[69][70].
Хирургические вмешательства на таламусе

При некоторых заболеваниях ЦНС, в частности, при болезни Паркинсона, эссенциальном треморе, синдроме таламической боли, иногда прибегают к хирургическим вмешательствам на таламусе, а именно — к таламотомии или к имплантации таламического стимулятора[71][72][73]. Однако, поскольку эти вмешательства являются инвазивными и необратимыми (особенно в случае таламотомии), то они обычно резервируются для использования у пациентов с тяжёлыми, инвалидизирующими формами этих заболеваний, оказавшихся резистентными к стандартному лечению[64]. Таламический стимулятор может также быть эффективен для устранения или уменьшения резистентных нейропатических болей, в частности, болей, не связанных с таламическим синдромом. Это было впервые показано ещё в 1977 году[74]. Эффективен таламический стимулятор и для лечения резистентной эпилепсии[75], резистентных форм синдрома Туретта[76].
Среди возможных побочных эффектов таламотомии или имплантации таламического стимулятора отмечаются, в частности, афазия и другие нарушения речи[72], депрессия и когнитивные нарушения[71], инфицирование во время операции, кровотечение, кровоизлияние в мозг, ишемический инсульт[64].
Примечания
- Thalamus // Foundational Model of Anatomy
- S. Sherman. Thalamus : [англ.] // Scholarpedia. — 2006. — Т. 1, № 9. — С. 1583. — ISSN 1941-6016. — doi:10.4249/scholarpedia.1583. — . — OCLC 4663345276.
- S. Murray Sherman, Ray W. Guillery. Exploring the Thalamus : [англ.]. — 1-е изд. — Academic Press, 2000. — 312 с. — ISBN 978-0123054609. — . — OCLC 494512886.
- Edward G. Jones. The Thalamus : [англ.] : in 2 vol.. — редакция от 1985 года. — Нью-Йорк : Springer, 2012. — 915 с. — ISBN 978-1-4615-1749-8. — doi:10.1007/978-1-4615-1749-8. — . — OCLC 970814982.
- Nicholas J. Strausfeld, Frank Hirth. Deep Homology of Arthropod Central Complex and Vertebrate Basal Ganglia : [англ.] // Science. — 2013. — Т. 340, № 6129 (12 April). — С. 157—161. — doi:10.1126/science.1231828. — PMID 23580521.
- Farries M. A. How ‘Basal' Are the Basal Ganglia? : [англ.] // Brain, Behavior and Evolution. — 2013. — Т. 82, № 4. — С. 211—214. — ISSN 1421-9743. — doi:10.1159/000356101. — PMID 24335184.
- María-Trinidad Herrero, Carlos Barcia, Juana Navarro. Functional anatomy of thalamus and basal ganglia : [англ.] // Child’s Nervous System. — 2002. — Т. 18, № 8 (August). — С. 386—404. — ISSN 0256-7040. — doi:10.1007/s00381-002-0604-1. — OCLC 4644394304. — PMID 12192499.
- Gerard Percheron. Chapter 20. Thalamus // The Human Nervous System : [англ.] / под ред. George Paxinos, Juergen Mai. — 2-е изд. — Academic Press, 2004. — С. 592—675. — 1366 с. — ISBN 978-0125476263. — doi:10.1016/B978-012547626-3/50021-1. — . — OCLC 4934574442.
- Darlene Susan Melchitzky, David A. Lewis. 1.2 Functional Neuroanatomy // Kaplan and Sadock's Comprehensive Textbook of Psychiatry : [англ.] : in 2 vol. / под ред. Benjamin J. Sadock, Virginia A. Sadock, Pedro Ruiz. — 10-е изд. — Lippincott Williams & Wilkins, 2017. — Thalamus. — С. 158—170. — 12754 (эл.), 4997 (бум.) с. — ISBN 978-1451100471. — . — OCLC 949866139.
- Giovanni Augusto Carlesimo, Maria Giovanna Lombardi, Carlo Caltagirone. Vascular thalamic amnesia: A reappraisal : [англ.] // Neuropsychologia. — 2011. — Т. 49, № 5 (April). — С. 777—789. — ISSN 0028-3932. — doi:10.1016/j.neuropsychologia.2011.01.026. — OCLC 4803930095. — PMID 21255590.
- Вашингтонский университет (Сиэтл). mamillothalamic tract (англ.). BrainInfo. Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Вашингтонский университет (Сиэтл). thalamocortical radiations (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017. Архивировано 16 сентября 2017 года.
- Вашингтонский университет (Сиэтл). thalamocortical systems (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Вашингтонский университет (Сиэтл). thalamoparietal fibers (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Вашингтонский университет (Сиэтл). spinothalamic tract (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Вашингтонский университет (Сиэтл). lateral spinothalamic tract (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Вашингтонский университет (Сиэтл). anterior spinothalamic tract (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Alan M. Rapoport. Analgesic Rebound Headache : [англ.] // Headache: The Journal of Head and Face Pain. — 1988. — Т. 28, № 10 (November). — С. 662—665. — ISSN 1526-4610. — doi:10.1111/j.1526-4610.1988.hed2810662.x. — OCLC 4658898144. — PMID 3068202.
- Golda Anne Kevetter, William D. Willis. Collateralization in the spinothalamic tract: New methodology to support or deny phylogenetic theories : [англ.] // Brain Research Reviews. — 1984. — Т. 7, № 1 (March). — С. 1—14. — ISSN 0006-8993. — doi:10.1016/0165-0173(84)90026-2. — OCLC 4643875849. — PMID 6370375.
- Вашингтонский университет (Сиэтл). thalamostriate fibers (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Вашингтонский университет (Сиэтл). central tegmental tract (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Вашингтонский университет (Сиэтл). Tractus cerebello-thalamo-cerebralis (англ.). BrainInfo. штат Вашингтон: washington.edu (27 сентября 2017). Дата обращения: 27 сентября 2017.
- Benno Schlesinger. IV. The Parenchymal Blood Vessels of the Upper Brainstem // The Upper Brainstem in the Human : Its Nuclear Configuration and Vascular Supply : [англ.]. — Springer Berlin Heidelberg, 1976. — С. 175—238. — 275 с. — ISBN 978-3-642-66257-7. — doi:10.1007/978-3-642-66255-3. — . — OCLC 858929573.
- Yuranga Weerakkody, Jeremy Jones et al. Thalamus (англ.). Radiopaedia.org (27 сентября 2017). Дата обращения: 27 сентября 2017. Архивировано 27 мая 2017 года.
- Gerard Percheron. The arterial supply of the thalamus // Stereotaxy of the Human Brain : Anatomical, Physiological and Clinical Applications : [англ.] / под ред. Georges Schaltenbrand, A. Earl Walker. — 2-е изд. — Штутгарт : Thieme Publishing Group, 1982. — С. 218—232. — 714 с. — ISBN 978-3135832029. — OCLC 8908048.
- Шилкин В.В., Филимонов В.И. Анатомия по Пирогову. : Атлас анатомии человека. : [] : в 3 т.. — Москва : ГЭОТАР-Медиа, 2011. — Т. 2. Голова и шея., Оболочки головного мозга. Вены головного мозга.. — С. 347, 351. — 724 с. — ББК Е860я61 Р457.844я61 Р457.845я61. — УДК 611(084.4). — ISBN 978-5-9704-1967-0.
- Abigail W. Leonard. Your Brain Boots Up Like a Computer (англ.). livescience.com (17 августа 2006). Дата обращения: 27 сентября 2017. Архивировано 12 июля 2017 года.
- Michael S. Gazzaniga, Richard B. Ivry, George R. Mangun. Cognitive Neuroscience: The Biology of the Mind : [англ.]. — 4-е изд. — Нью-Йорк : W. W. Norton & Company, 2013. — 752 с. — ISBN 978-0393913484. — . — OCLC 908087478.
- Mircea Steriade, Rodolfo R. Llinas. The functional states of the thalamus and the associated neuronal interplay : [англ.] // Physiological Reviews. — 1988. — Т. 68, № 3 (July). — С. 649—742. — ISSN 1522-1210. — OCLC 114111014. — PMID 2839857.
- E. V. Evarts, W. T. Thach. Motor Mechanisms of the CNS: Cerebrocerebellar Interrelations : [англ.] // Annual Review of Physiology. — 1969. — Т. 31 (March). — С. 451—498. — doi:10.1146/annurev.ph.31.030169.002315. — OCLC 4761161312. — PMID 4885774.
- Paul J. Orioli, Peter L. Strick. Cerebellar connections with the motor cortex and the arcuate premotor area: An analysis employing retrograde transneuronal transport of WGA-HRP : [англ.] // The Journal of Comparative Neurology. — 1989. — Т. 288, № 4 (22 October). — С. 612—626. — ISSN 1096-9861. — doi:10.1002/cne.902880408. — OCLC 118356328. — PMID 2478593.
- C. Asanuma, W. T. Thach, E. G. Jones. Cytoarchitectonic delineation of the ventral lateral thalamic region in the monkey : [англ.] // Brain Research Reviews. — 1983. — Т. 5(286), № 3 (May). — С. 219—235. — ISSN 0006-8993. — doi:10.1016/0165-0173(83)90014-0. — OCLC 4929455787. — PMID 6850357.
- Kiyoshi Kurata. Activity Properties and Location of Neurons in the Motor Thalamus That Project to the Cortical Motor Areas in Monkeys : [англ.] // Journal of Neurophysiology. — 2005. — Т. 94, № 1 (1 July). — С. 550—566. — ISSN 1522-1598. — doi:10.1152/jn.01034.2004. — OCLC 110603384. — PMID 15703228.
- Jun Kunimatsu, Masaki Tanaka. Roles of the Primate Motor Thalamus in the Generation of Antisaccades : [англ.] // Journal of Neuroscience. — 2010. — Т. 30, № 14 (1 April). — С. 5108—5117. — ISSN 0270-6474. — doi:10.1523/JNEUROSCI.0406-10.2010. — OCLC 605093762. — PMID 20371831.
- Burkhart Fischer, Stefan Everling. The Antisaccade: A Review of Basic Research and Clinical Studies (англ.). Optomotorik.de. Фрайбург: Университет Фрайбурга. Дата обращения: 28 сентября 2017. Архивировано 16 сентября 2017 года.
- Stefan Everling, Burkhart Fischer. The antisaccade: a review of basic research and clinical studies : [англ.] // Neuropsychologia. — 1998. — Т. 36, № 9 (1 September). — ISSN 0028-3932. — doi:10.1016/S0028-3932(98)00020-7. — OCLC 4924629675. — PMID 9740362.
- Thor Stein, Chad Moritz, Michelle Quigley, Dietmar Cordes, Victor Haughton, Elizabeth Meyerand. Functional Connectivity in the Thalamus and Hippocampus Studied with Functional MR Imaging : [англ.] // American Journal of Neuroradiology. — 2000. — Т. 21, № 8 (September). — С. 1397—1401. — ISSN 0195-6108. — OCLC 199701670. — PMID 11003270.
- John P. Aggleton, Malcolm W. Brown. Episodic memory, amnesia, and the hippocampal–anterior thalamic axis : [англ.] // Behavioral and Brain Sciences. — 1999. — Т. 22, № 3 (June). — С. 425—444; дискуссия на стр. 444-489. — ISSN 1469-1825. — doi:10.1017/S0140525X99002034. — OCLC 4669514763. — PMID 11301518.
- John P. Aggleton, Shane M. O’Mara, Seralynne D. Vann, Nick F. Wright, Marian Tsanov, Jonathan T. Erichsen. Hippocampal–anterior thalamic pathways for memory: uncovering a network of direct and indirect actions : [англ.] // European Journal of Neuroscience. — 2010. — Т. 31, № 12 (14 June). — С. 2292–2307. — ISSN 1460-9568. — doi:10.1111/j.1460-9568.2010.07251.x. — OCLC 5151632719. — PMID 20550571. — PMC 2936113.
- Neil Burgess, Eleanor A Maguire, John O'Keefe. The Human Hippocampus and Spatial and Episodic Memory : [англ.] // Neuron. — 2002. — Т. 35, № 4 (15 August). — С. 625—641. — ISSN 0896-6273. — doi:10.1016/S0896-6273(02)00830-9. — OCLC 198675262. — PMID 12194864.
- The Medical Dictionary. paleothalamus (англ.). The Medical Dictionary. FarLex Inc. Дата обращения: 28 сентября 2017.
- Merriam Webster Inc. neothalamus (англ.). Merriam Webster Medical Dictionary. Merriam Webster Inc.. Дата обращения: 28 сентября 2017. Архивировано 18 сентября 2017 года.
- Hartwig Kuhlenbeck. The ontogenetic development of the diencephalic centers in a bird's brain (chick) and comparison with the reptilian and mammalian diencephalon : [англ.] // The Journal of Comparative Neurology. — 1937. — Т. 66, № 1 (February). — С. 23–75. — ISSN 1096-9861. — doi:10.1002/cne.900660103. — OCLC 4641762835.
- Kenji Shimamura, Dennis J. Hartigan, Salvador Martinez, Luis Puelles, John L. R. Rubenstein. Longitudinal organization of the anterior neural plate and neural tube : [англ.] // Development. — 1995. — Т. 121, № 12 (December). — С. 3923—3933. — ISSN 1477-9129. — OCLC 192459955. — PMID 8575293.
- Steffen Scholpp, Andrew Lumsden. Building a bridal chamber: development of the thalamus : [англ.] // Trends in Neurosciences. — 2010. — Т. 33, № 8 (August). — С. 373–380. — ISSN 0166-2236. — doi:10.1016/j.tins.2010.05.003. — OCLC 654635968. — PMID 20541814. — PMC 2954313.
- Müller Fabiola, O'Rahilly Ronan. The timing and sequence of appearance of neuromeres and their derivatives in staged human embryos : [англ.] // Acta Anatomica. — 1997. — Т. 158, № 2. — С. 83—99. — ISSN 1422-6421. — doi:10.1159/000147917. — OCLC 86493197. — PMID 9311417.
- O'Rahilly Ronan, Müller Fabiola. The longitudinal growth of the neuromeres and the resulting brain in the human embryo : [англ.] // Cells Tissues Organs. — 2013. — Т. 197, № 3 (February). — С. 178—195. — ISSN 1422-6421. — doi:10.1159/000343170. — OCLC 5817230667. — PMID 23183269.
- Mallika Chatterjee, Qiuxia Guo, James Y.H. Li. Gbx2 is essential for maintaining thalamic neuron identity and repressing habenular characters in the developing thalamus : [англ.] // Developmental Biology. — 2015. — Т. 407, № 1 (1 November). — С. 26—39. — ISSN 0012-1606. — doi:10.1016/j.ydbio.2015.08.010. — OCLC 5913930043. — PMID 26297811. — PMC 4641819.
- Tsutomu Hirata, Masato Nakazawa, Osamu Muraoka, Rika Nakayama, Yoko Suda, Masahiko Hibi. Zinc-finger genes Fez and Fez-like function in the establishment of diencephalon subdivisions : [англ.] // Development. — 2006. — Т. 133, № 20 (October). — С. 3993—4004. — ISSN 1477-9129. — doi:10.1242/dev.02585. — OCLC 202024440. — PMID 16971467.
- Jae-Yeon Jeong, Zev Einhorn, Priya Mathur, Lishan Chen, Susie Lee, Koichi Kawakami, Su Guo. Patterning the zebrafish diencephalon by the conserved zinc-finger protein Fezl : [англ.] // Development. — 2007. — Т. 134, № 1 (January). — С. 127—136. — ISSN 1477-9129. — doi:10.1242/dev.02705. — OCLC 4636344085. — PMID 17164418.
- Dario Acampora, Virginia Avantaggiato, Francesca Tuorto, Antonio Simeone. Genetic control of brain morphogenesis through Otx gene dosage requirement : [англ.] // Development. — 1997. — Т. 124, № 18 (September). — С. 3639—3650. — ISSN 1477-9129. — OCLC 200505171. — PMID 9342056.
- Steffen Scholpp, Isabelle Foucher, Nicole Staudt, Daniela Peukert, Andrew Lumsden, Corinne Houart. Otx1l, Otx2 and Irx1b establish and position the ZLI in the diencephalon : [англ.] // Development. — 2007. — Т. 134, № 17 (September). — С. 3167—3176. — ISSN 1477-9129. — doi:10.1242/dev.001461. — OCLC 211790140. — PMID 17670791.
- Hobeom Song, Bumwhee Lee, Dohoon Pyun, Jordi Guimera, Youngsook Son, Jaeseung Yoon, Kwanghee Baek, Wolfgang Wurst, Yongsu Jeong. Ascl1 and Helt act combinatorially to specify thalamic neuronal identity by repressing Dlxs activation : [англ.] // Developmental Biology. — 2015. — Т. 398, № 2 (15 February). — С. 280–291. — ISSN 0012-1606. — doi:10.1016/j.ydbio.2014.12.003. — OCLC 5712498415. — PMID 25512300.
- Luis Puelles, John L.R. Rubenstein. Forebrain gene expression domains and the evolving prosomeric model : [англ.] // Trends in Neurosciences. — 2003. — Т. 26, № 9 (September). — С. 469–476. — ISSN 0166-2236. — doi:10.1016/S0166-2236(03)00234-0. — OCLC 112198916. — PMID 12948657.
- Makoto Ishibashi, Andrew P. McMahon. A sonic hedgehog-dependent signaling relay regulates growth of diencephalic and mesencephalic primordia in the early mouse embryo : [англ.] // Development. — 2002. — Т. 129, № 20 (October). — С. 4807—4819. — ISSN 1477-9129. — OCLC 200691112. — PMID 12361972.
- Clemens Kiecker, Andrew Lumsden. Hedgehog signaling from the ZLI regulates diencephalic regional identity : [англ.] // Nature Neuroscience. — 2004. — Т. 7, № 11 (November). — С. 1242—1249. — doi:10.1038/nn1338. — OCLC 201081969. — PMID 15494730.
- Steffen Scholpp, Olivia Wolf, Michael Brand, Andrew Lumsden. Hedgehog signalling from the zona limitans intrathalamica orchestrates patterning of the zebrafish diencephalon : [англ.] // Development. — 2006. — Т. 133, № 5 (March). — С. 855—864. — ISSN 1477-9129. — doi:10.1242/dev.02248. — OCLC 4636339052. — PMID 16452095.
- Steffen Scholpp, Alessio Delogu, Jonathan Gilthorpe, Daniela Peukert, Simone Schindler, Andrew Lumsden. Her6 regulates the neurogenetic gradient and neuronal identity in the thalamus : [англ.] // Proceedings of the National Academy of Sciences of the United States of America. — 2009. — Т. 106, № 47 (24 November). — С. 19895—19900. — ISSN 1091-6490. — doi:10.1073/pnas.0910894106. — OCLC 488933764. — PMID 19903880. — PMC 2775703.
- Tou Yia Vue, Krista Bluske, Amin Alishahi, Lin Lin Yang, Naoko Koyano-Nakagawa, Bennett Novitch, Yasushi Nakagawa. Sonic Hedgehog Signaling Controls Thalamic Progenitor Identity and Nuclei Specification in Mice : [англ.] // Journal of Neuroscience. — 2009. — Т. 29, № 14 (1 April). — С. 4484—4497. — ISSN 1529-2401. — doi:10.1523/JNEUROSCI.0656-09.2009. — OCLC 4633866223. — PMID 19357274. — PMC 2718849.
- Keith A. Young, Leigh A. Holcomb, Willy L. Bonkale, Paul B. Hicks, Umar Yazdani, Dwight C. German. 5HTTLPR Polymorphism and Enlargement of the Pulvinar: Unlocking the Backdoor to the Limbic System : [англ.] // Biological Psychiatry. — 2007. — Т. 61, № 6 (15 March). — С. 813–818. — ISSN 0006-3223. — doi:10.1016/j.biopsych.2006.08.047. — OCLC 4922785860. — PMID 17083920.
- Rajal A. Patel, James P. Chandler, Sarika Jain, Mahesh Gopalakrishnan, Sean Sachdev. Dejerine-Roussy syndrome from thalamic metastasis treated with stereotactic radiosurgery : [англ.] // Journal of Clinical Neuroscience. — 2017. — Т. 44 (October). — С. 227–228. — doi:10.1016/j.jocn.2017.06.025. — OCLC 7065358380. — PMID 28684151.
- J. Dejerine, G. Roussy. Le syndrome thalamique : [фр.] // Revue Neurologique. — 1906. — Т. 14. — С. 521—532. — OCLC 755636738.
- Tülay Kamaşak, Sevim Sahin, İlker Eyüboğlu, Gökce Pinar Reis, Ali Cansu. Bilateral Paramedian Thalamic Syndrome After Infection : [англ.] // Pediatric Neurology. — 2015. — Т. 52, № 2 (February). — С. 235–238. — doi:10.1016/j.pediatrneurol.2014.09.012. — OCLC 5776963712. — PMID 25693586.
- Rodolfo R. Llinás, Urs Ribary, Daniel Jeanmonod, Eugene Kronberg, Partha P. Mitra. Thalamocortical dysrhythmia: A neurological and neuropsychiatric syndrome characterized by magnetoencephalography : [англ.] // Proceedings of the National Academy of Sciences of United States of America. — 1999. — Т. 96, № 26 (21 December). — С. 15222–15227. — ISSN 1091-6490. — doi:10.1073/pnas.96.26.15222. — PMID 10611366. — PMC 24801.
- Axel Sandvig, Sandra Lundberg, Jiri Neuwirth. Artery of Percheron infarction: a case report : [англ.] // Journal of Medical Case Reports. — 2017. — Т. 11, № 1 (12 August). — С. 221. — ISSN 1752-1947. — doi:10.1186/s13256-017-1375-3. — OCLC 7103544786. — PMID 28800746. — PMC 5554405.
- Franc Llorens, Juan-José Zarranz, Andre Fischer, Inga Zerr, Isidro Ferrer. Fatal Familial Insomnia: Clinical Aspects and Molecular Alterations : [англ.] // Current Neurology and Neuroscience Reports. — 2017. — Т. 17, № 4 (April). — С. 30. — ISSN 1534-6293. — doi:10.1007/s11910-017-0743-0. — OCLC 6994559043. — PMID 28324299.
- Вячеслав Дубынин. Таламус и гипоталамус. PostNauka.ru (16 февраля 2017). Дата обращения: 28 октября 2017. Архивировано 28 октября 2017 года.
- Maria Engström, Thomas Karlsson, Anne-Marie Landtblom. Thalamic activation in the Kleine-Levin syndrome : [англ.] // Sleep. — 2014. — Т. 37, № 2 (1 February). — С. 379—386. — ISSN 1550-9109. — doi:10.5665/sleep.3420. — PMID 24497666. — PMC 3900625.
- Maria Engström, Tove Hallböök, Attila Szakacs, Thomas Karlsson, Anne-Marie Landtblom. Functional Magnetic Resonance Imaging in Narcolepsy and the Kleine–Levin Syndrome : [англ.] // Frontiers in Neurology. — 2014. — Т. 5 (25 June). — С. 105. — ISSN 1664-2295. — doi:10.3389/fneur.2014.00105. — PMID 25009530. — PMC 4069720.
- Dang-Vu TT. Neuroimaging findings in narcolepsy with cataplexy : [англ.] // Current Neurology and Neuroscience Reports. — 2013. — Т. 13, № 5 (May). — С. 349—351. — ISSN 1534-6293. — doi:10.1007/s11910-013-0349-0. — PMID 23526549.
- Julie A.Fields, Alexander I.Tröster. Cognitive Outcomes after Deep Brain Stimulation for Parkinson's Disease: A Review of Initial Studies and Recommendations for Future Research : [англ.] // Brain and Cognition. — 2000. — Т. 42, № 2 (March). — С. 268—293. — ISSN 0278-2626. — doi:10.1006/brcg.1999.1104. — OCLC 359047453. — PMID 10744924.
- Bruce B.B., Foote K.D., Rosenbek J., Sapienza C., Romrell J., Crucian G., Okun M.S. Aphasia and Thalamotomy: Important Issues : [англ.] // Stereotactic and Functional Neurosurgery. — 2004. — Т. 82, № 4 (December). — С. 186—190. — ISSN 1423-0372. — doi:10.1159/000082207. — OCLC 202401175. — PMID 15557767.
- Justin S. Cetas, Targol Saedi, Kim J. Burchiel. Destructive procedures for the treatment of nonmalignant pain: a structured literature review : [англ.] // Journal of Neurosurgery. — 2008. — Т. 109, № 3 (September). — С. 389—404. — ISSN 1933-0693. — doi:10.3171/JNS/2008/109/9/0389. — OCLC 4665611587. — PMID 18759567.
- Mundinger F. Treatment of chronic pain with intracerebral stimulators : [нем.] // Dtsch Med Wochenschr. — 1977. — Т. 102, № 47 (25 November). — С. 1724—1729. — doi:10.1055/s-0028-1105565. — PMID 303562.
- Robert S. Fisher, Sumio Uematsu, Gregory L. Krauss, Barbara J. Cysyk, Robert McPherson, Ronald P. Lesser, Barry Gordon, Pamela Schwerdt, Mark Rise. Placebo-Controlled Pilot Study of Centromedian Thalamic Stimulation in Treatment of Intractable Seizures : [англ.] // Epilepsia. — 1992. — Т. 33, № 5 (September). — С. 841—851. — ISSN 1528-1167. — doi:10.1111/j.1528-1157.1992.tb02192.x. — PMID 1396427.
- Paola Testini, Cong Z. Zhao, Matt Stead, Penelope S. Duffy, Bryan T. Klassen, Kendall H. Lee. Centromedian-Parafascicular Complex Deep Brain Stimulation for Tourette Syndrome: A Retrospective Study : [англ.] // Mayo Clinic Proceedings. — 2016. — Т. 91, № 2 (February). — С. 218–225. — ISSN 0025-6196. — doi:10.1016/j.mayocp.2015.11.016. — PMID 26848003. — PMC 4765735.
Ссылки
- Анатомия головного мозга. Таламус
- Интегральная медицина XXI ВЕКА: теория и практика. Таламус
- Физиология человека под ред. В. М. Покровского, Г. Ф. Коротько. Таламус
| В библиографических каталогах |
|---|