Хордовые
Хо́рдовые (лат. Chordata) — тип вторичноротых животных, для которых характерно наличие энтодермального осевого скелета в виде хорды, которая у высших форм заменяется позвоночником. По степени развития нервной системы тип хордовых занимает высшее место среди всех животных. В мире известно более 60 000 видов хордовых.
| Хордовые | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд: тигр и асцидия Polycarpa aurata 2-й ряд: голубая сойка и Enchelycore anatina | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Хордовые |
||||||||||||
| Международное научное название | ||||||||||||
| Chordata Haeckel, 1874[1][2] | ||||||||||||
| ||||||||||||
По определению Cameron, 2020, хордовые — крупнейшая кроновая группа, включающая морскую миногу (Petromyzon marinus), но не пурпурного стронгилоцентротуса (Strongylocentrotus purpuratus)[3]. Понятие хордовые объединяет позвоночных и некоторых беспозвоночных, имеющих, по крайней мере в некоторый период их жизненного цикла, хорду, полый спинной нервный тяж, жаберные щели, эндостиль и хвост, располагающийся позади смещённого на вентральную сторону тела анального отверстия. Тип хордовых делится на три подтипа: бесчерепных, оболочников, позвоночных — единственный подтип, имеющий череп. Ранее в качестве четвёртого подтипа рассматривались полухордовые, которые теперь вынесены в классификации в отдельную группу.
Особенности строения
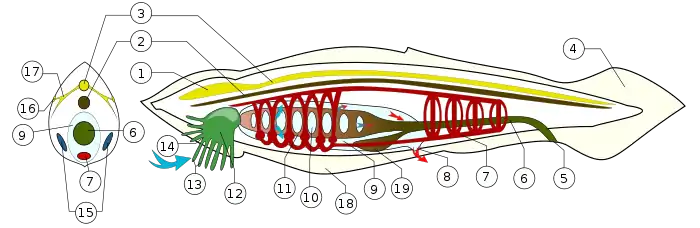
Хордовые — тип животных, характеризующихся билатеральной симметрией и наличием, по крайней мере, на определённых стадиях развития следующих признаков:
- Хорда, представляющая собой эластичный стержень, образующийся путём выпячивания спинной стороны кишечной трубки. У большинства позвоночных хорда в ходе эмбрионального развития полностью или частично замещается хрящевой и костной тканью, образующей позвоночник.
- Нервная трубка, расположенная дорсально. У позвоночных развивается в спинной мозг и головной мозг.
- Жаберные щели — парные отверстия в глотке. У низших хордовых участвуют в фильтрации воды для питания. У наземных позвоночных жаберные щели закладываются в раннем эмбриогенезе в виде жаберных мешочков.
- Мышечный хвост — постанальный отдел тела, расположенный каудальнее смещённого на брюшную сторону тела ануса (в него заходят хорда и нервная трубка, но не заходит кишечник).
- Эндостиль — желобок на вентральной стороне глотки. У низших хордовых-фильтраторов в нём производится слизь, помогающая собирать частицы пищи и доставлять их в пищевод. Также в нём накапливается иод и, вероятно, он является предшественником щитовидной железы позвоночных. Как таковой, эндостиль у позвоночных есть только у пескоройки.
Происхождение
В настоящее время достигнуто согласие, что хордовые — монофилетическая группа (являются потомками одного общего предка, который сам был хордовым), а ближайшими родственниками позвоночных (лат. Vertebrata) являются оболочники (лат. Tunicata)[4][5]. Поскольку окаменелости ранних хордовых плохо сохранились, выяснить родственные связи представителей типа можно только с помощью молекулярной филогенетики.
Билатеральные животные делятся на две большие группы — первичноротые и вторичноротые. Хордовые относятся ко вторичноротым. Весьма вероятно, что ископаемое кимберелла, жившая 555 млн лет назад, принадлежала к первичноротым[6]. Жившая 549—543 млн лет назад в эдиакарии эрниетта была уже явно вторичноротым животным[7]. Таким образом, первичноротые и вторичноротые должны были разделиться до времени существования этих животных, то есть до начала кембрийского периода[8].
Первые известные ископаемые двух близких к хордовым групп — иглокожих и полухордовых — обнаруживаются с раннего и среднего кембрия соответственно[9][10]. Кроме того, известно ископаемое чэнцзянской биоты Yunnanozoon, принадлежность которого к полухордовым или хордовым не определена[11][12]. Другое ископаемое, относящееся к той же биоте, Haikouella lanceolata явно является хордовым и, возможно, позвоночным. У него обнаружены признаки сердца, артерий, нервной трубки и мозга, хвоста, жаберных лепестков, возможно, глаз, но в то же время вокруг ротового отверстия есть щупальца[12]. Haikouichthys и Myllokunmingia, также из чэнцзянской фауны, и пикайя из сланцев Бёрджес относятся к примитивным позвоночным[13][14]. Ископаемые ранних хордовых весьма редки, поскольку у них не было твёрдых частей тела.
Исследования родственных отношений хордовых начались с 90-х годов XIX века. Они основывались на анатомических, эмбриологических и палеонтологических данных и приводили к разным филогенетическим деревьям. Некоторое время ближайшими современными родственниками хордовых считали полухордовых[15], но позже ими оказалась группа, состоящая из полухордовых и иглокожих (Ambulacraria)[16].
Время разделения эволюционных линий хордовых и Ambulacraria на основании метода молекулярных часов было оценено в 896 млн лет назад[17].
Ниже приведена кладограмма, основанная на сравнении последовательностей митохондриальных ДНК[18] (положение Xenoturbellida по результатам некоторых других молекулярных исследований значительно отличается)[19]:
| Deuterostomia |
| ||||||||||||||||||||||||||||||||||||
Классификация
Обычно выделяют три подтипа хордовых (иногда четыре). Высшим подтипом являются позвоночные, к которым принадлежит порядка 95 % всех видов хордовых. Из низших хордовых выделяют бесчерепных и оболочников.
Ниже перечислены три общепризнанных подтипа хордовых вместе с входящими в них классами и подклассами[20].
Тип Хордовые (Chordata)
- Подтип Бесчерепные (Acraniata) или головохордовые (Cephalochordata)
- Класс Ланцетники (Leptocardii)
- Подтип Оболочники (Tunicata) или личиночнохордовые (Urochordata)
- Класс Аппендикулярии (Appendicularia)
- Класс Асцидии (Ascidiacea)
- Класс Сальпы (Thaliacea): сальпиды, огнетелки и бочёночники
- Подтип Позвоночные (Vertebrata)
- † Класс Конодонты (Conodonta)
- Надкласс Бесчелюстные (Agnatha) — парафилетическая группа
- † Класс Анаспиды, или беспанцирные, или бесщитковые (Anaspida)
- † Класс Непарноноздрёвые, или цефаласпидоморфы (Cephalaspidomorphi)
- † Класс Парноноздрёвые, или птероспидоморфы (Pteraspidomorpha)
- † Класс Телодонты (Thelodonti)
- Класс Миксины (Myxini)
- Класс Миноги (Petromyzontida)
- Инфратип Челюстноротые (Gnathostomata)
- † Класс Плакодермы или пластинокожие рыбы (Placodermi)
- † Класс Акантоды или колючкозубые (Acanthodii)
- Класс Хрящевые рыбы (Chondrichthyes)
- Подкласс Пластиножаберные (Elasmobranchii): акулы и скаты
- Подкласс Цельноголовые (Holocephali)
- Группа Костные рыбы (Osteichthyes)
- Класс Лучепёрые рыбы (Actinopterygii)
- Класс Лопастеперые рыбы, или мясистолопастные (Sarcopterygii) → Надкласс Четвероногие (Tetrapoda)
- Надкласс Четвероногие (Tetrapoda)
- Класс Земноводные, или амфибии (Amphibia) / Батрахоморфы (Batrachomorpha)
- Класс Завропсиды (Sauropsida), традиционно — Пресмыкающиеся, или рептилии (Reptilia)
- † Подкласс Парарептилии (Parareptilia)
- Подкласс Диапсиды (Diapsida) → Класс Птицы (Aves)
- Класс Синапсиды (Synapsida) → Класс Млекопитающие (Mammalia)
Завропсиды (пресмыкающиеся и птицы) и синапсиды (млекопитающие и вымершие родственники) относятся к кладе амниот, остальные классы позвоночных — к парафилетической группе анамний.
Иногда отряды огнетелок, сальп и бочёночников повышают до классов в подтипе Tunicata. Также существует довольно много альтернативных классификаций позвоночных. Их наличие связано, в частности, с тем, что многие традиционно выделяемые группы позвоночных парафилетичны. Например, парафилетичны лопастепёрые (чтобы эта группа стала монофилетичной, в неё надо включить наземных позвоночных) и рептилии (в них с позиций кладистической систематики следовало бы включить класс птиц).
Очень часто используется несистемная группа беспозвоночные, которая включает два подтипа хордовых (головохордовые и оболочники) и все остальные типы животных. Её использование подчёркивает важное значение, которое имеют позвоночные в животном мире и жизни человека.
Хронограмма
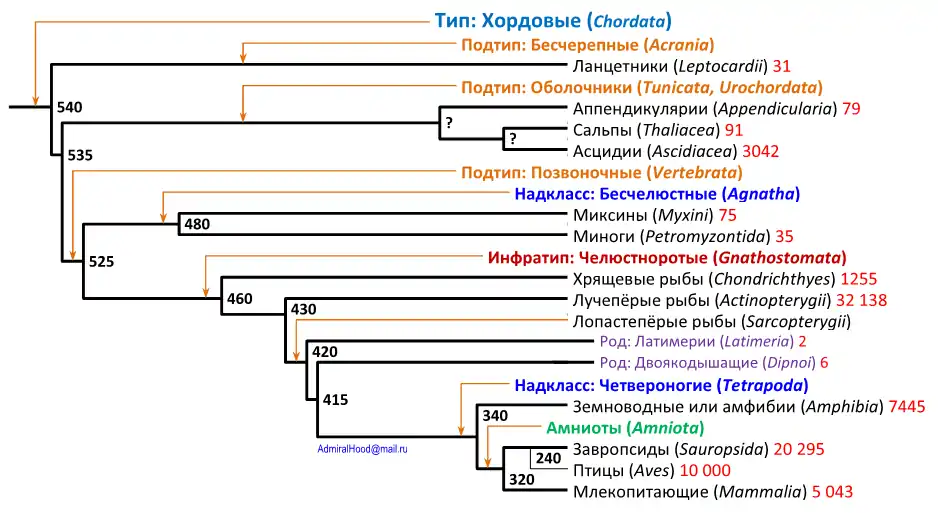
На рисунке показана хронограмма современных классов хордовых.
При кладистическом подходе сопоставление иерархии таксономических групп с ветвями филогенетического дерева сталкивается с определёнными трудностями. Например, при этом подходе класс лопастепёрых рыб содержит 8 видов собственно рыб, а также всех наземных хордовых (четвероногих), поскольку они произошли от лопастепёрых рыб. Между тем, традиционно четвероногие делятся на классы земноводных, пресмыкающихся, птиц и млекопитающих, то есть в кладистической системе несколько традиционных классов оказываются подчинёнными другому классу.
Не меньшие проблемы возникают в надклассе четвероногих. Невозможно выделить отдельную ветвь для рептилий, так как они не являются монофилетической группой (произошедшие от рептилий птицы традиционно выделяются в отдельный класс). В результате выделен класс завропсид, включающий в себя пресмыкающихся и птиц, а выделение птиц из завропсид показано тонкой линией. При этом нужно учитывать, что обозначенное на дереве число видов птиц входит в число видов завропсид.
Подтипы
Головохордовые

Головохо́рдовые (лат. Cephalochordata) или бесчерепны́е (лат. Acrania)[21] — небольшие морские рыбообразные животные со всеми признаками, свойственными хордовым. Бесчерепные — подтип низших хордовых животных, в отличие от других хордовых (оболочников и позвоночных), сохраняющий основные признаки типа (хорда, нервная трубка и жаберные щели) в течение всей жизни. Головной мозг отсутствует, органы чувств примитивны[22]. Ведут придонный образ жизни, по характеру питания — роющие фильтраторы[23][24]. Всего к бесчерепным относятся около 30 видов, составляющих один класс — ланцетники.
Оболочники

Оболо́чники (лат. Tunicata, Urochordata) — подтип хордовых животных. Включают 5 классов — асцидий, аппендикулярий, сальп, огнетелок и бочёночников. По другой классификации последние 3 класса считаются отрядами класса Thaliacea[25]. Известно более 1 000 видов. Большинство из них имеют хорду только на личиночной стадии. Они распространены по всему миру и населяют морское дно.
Три крупных класса оболочников:
- Асцидии — низшие мягкотелые хордовые-фильтраторы, во взрослом состоянии ведущие сидячий образ жизни[26];
- аппендикулярии сохраняют личиночные черты, такие как хвост, на протяжении всей жизни. По этой причине долгое время рассматривались как личинки асцидий и сальп[27]. Из-за наличия длинных хвостов личинки оболочников называют лат. urochordata[26];
- третья группа оболочников — свободноплавающие сальпы, питаются планктоном. В их жизненном цикле известно два поколения — одиночное гермафродитное и почкующееся колониальное бесполое. У личинок этих животных есть все основные признаки хордовых, в том числе хорда и хвост. Они также оснащены рудиментарным мозгом и датчиками освещённости и положения (крена)[26].
Позвоночные

Позвоно́чные (лат. Vertebrata) — высший подтип хордовых животных. Доминирующая (наряду с насекомыми) на земле и в воздушной среде группа животных. Отличаются от других хордовых наличием обособленного черепа и развитием головного мозга и органов чувств. Хорда у большинства представителей высших хордовых замещается на позвоночник[28], защищающий спинной мозг и состоящий, как правило, из хрящевой и костной ткани. Эндостиль, как таковой, присутствует только у личинок миног[29]. По сравнению с низшими хордовыми — бесчерепными и оболочниками — они характеризуются значительно более высоким уровнем организации, что наглядно выражено как в их строении, так и в физиологических отправлениях. Среди позвоночных нет видов, ведущих сидячий (прикреплённый) образ жизни. Они перемещаются в широких пределах, активно разыскивая и захватывая пищу, находя для размножения особей другого пола, спасаясь от преследования врагов.
У миног есть недоразвитый череп и рудиментарные позвонки — следовательно, они могут рассматриваться как позвоночные[30]. Миксиновые, имеющие жаберный скелет, состоящий из небольшого числа хрящевых пластинок, рудиментарные позвонки, не всегда рассматриваются как истинные позвоночные[31], но молекулярная филогенетика показала, что это сестринская группа миног, образующая вместе с ними группу круглоротых[32].
Ближайшие родственники
Полухордовые

Полухо́рдовые (лат. Hemichordata) — тип морских донных беспозвоночных из группы вторичноротых. Мягкотелые червеобразные животные. В мире известно 130 видов[33], в России — 4 вида[34]. Первоначально полухордовые считались подтипом или близкими родственниками хордовых, с которыми их объединяет наличие жаберных щелей, спинного нервного ствола и хордоподобного органа (стомохорда), представляющего собой вырост кишечника, поддерживающий хоботок. Однако стомохорд не гомологичен хорде. Кроме того, в отличие от хордовых, у полухордовых тело чётко делится на 3 отдела: хоботок (у кишечнодышащих) или головной щит (у перистожаберных), воротник и туловище. Есть и ещё ряд отличий от хордовых: строение сердца, нервной системы и другие особенности внутренней организации, например, помимо спинного, у полухордовых есть и брюшной нервный тяж. Выделяют два класса: кишечнодышащие (лат. Enteropneusta) — 108 современных видов — и перистожаберные (лат. Pterobranchia) — 22 современных вида[33]. К последним принадлежат граптолиты, широко распространённые в палеозое и ранее считавшиеся отдельным классом[33].
Сходство личинок полухордовых (торнарии) с личинками-бипиннариями иглокожих позволило считать полухордовых промежуточным звеном между иглокожими и хордовыми.
Иглокожие

Иглоко́жие (лат. Echinodermata) — тип исключительно морских донных животных, большей частью свободноживущих, реже сидячих, встречающихся на любых глубинах Мирового океана. Насчитывается около 7000 современных видов (в России — 400)[35]. Наряду с хордовыми, иглокожие относятся к ветви вторичноротых животных (Deuterostomia). Современными представителями типа являются морские звёзды, морские ежи, офиуры (змеехвостки), голотурии (морские огурцы) и морские лилии. В состав этого типа входят также приблизительно 13 000 вымерших видов, которые процветали в морях, начиная с раннего кембрия.
Иглокожие отличаются от хордовых тремя признаками:
- вместо двусторонней (билатеральной) они обладают радиальной симметрией;
- схема строения их тела напоминает кольцо (в виде колеса с трубами-ногами);
- их тела поддерживает скелет из кальцита, материала, не встречающегося у хордовых.
Несмотря на то, что тела иглокожих хорошо защищены от воздействия окружающей среды кальцинированной оболочкой (скелетом), снаружи они покрыты тонкой кожей. Ноги иглокожих окружены мышцами и выполняют функцию дыхания и питания — действуют как насосы, обеспечивая циркуляцию воды рядом с кровеносными сосудами, осуществляя газообмен и фильтрацию частиц пищи из воды. Видоизменённые ноги-перья несут и оборонительную функцию. Некоторые иглокожие, например морские лилии, живут оседло (прикреплены к скалам), и немногие из них способны очень медленно перемещаться. Другие иглокожие мобильны, например, морские звёзды, морские ежи и морские огурцы[36].
Примечания
- Haeckel E. Anthropogenie oder Entwicklungsgeschichte des Menschen (нем.). — Leipzig: Engelmann, 1874.
- Nielsen C. The authorship of higher chordate taxa (англ.) // Zoologica Scripta : journal. — 2012. — Vol. 41, iss. 4. — P. 435—436. — ISSN 1463-6409. — doi:10.1111/j.1463-6409.2012.00536.x. — .
- Chordata E. Haeckel 1874 [C. B. Cameron], converted clade name // Phylonyms: A Companion to the PhyloCode (англ.) / Kevin de Queiroz, Philip D. Cantino, Jacques A. Gauthier. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — P. 661. — 1352 p. — ISBN 978-1-138-33293-5.
- Марков А. В. Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных. Элементы.ру (23 июня 2008). Архивировано 4 августа 2019 года.
- Ястребов С. А. Общий предок вторичноротых мог быть похож на хордовое. Элементы.ру (18 марта 2015). Архивировано 4 декабря 2018 года.
- Fedonkin, M. A.; Simonetta, A; Ivantsov, A. Y. New data on Kimberella, the Vendian mollusc-like organism (White sea region, Russia): palaeoecological and evolutionary implications. — In: The Rise and Fall of the Ediacaran Biota : [англ.] / Vickers-Rich, Patricia; Komarower, Patricia // Geological Society. — London, 2007. — Vol. 286. — P. 157—179. — (Special publications). — ISBN 9781862392335. — doi:10.1144/SP286.12.
- Dzik, J. (June 1999). «Organic membranous skeleton of the Precambrian metazoans from Namibia». Geology 27 (6): 519—522. doi:10.1130/0091-7613(1999)027<0519:OMSOTP>2.3.CO;2
- Erwin, Douglas H.; Eric H. Davidson (July 1, 2002). «The last common bilaterian ancestor». Development 129 (13): 3021-3032
- Bengtson, S. (2004). Early skeletal fossils. In Lipps, J.H., and Waggoner, B.M.. «Neoproterozoic-Cambrian Biological Revolutions». Paleontological Society Papers 10: 67-78. doi:10.1017/S1089332600002345.
- Bengtson, S., and Urbanek, A. (1986). «Rhabdotubus, a Middle Cambrian rhabdopleurid hemichordate». Lethaia 19 (4): 293—308. doi:10.1111/j.1502-3931.1986.tb00743.x
- Shu, D., Zhang, X. and Chen, L. (April 1996). «Reinterpretation of Yunnanozoon as the earliest known hemichordate». Nature 380 (6573): 428—430. doi:10.1038/380428a0.
- Chen, J-Y., Hang, D-Y., and Li, C.W. (December 1999). «An early Cambrian craniate-like chordate». Nature 402 (6761): 518—522. doi:10.1038/990080.
- Shu, D-G., Conway Morris, S., and Zhang, X-L. (November 1999). «Lower Cambrian vertebrates from south China». Nature 402 (6757). doi:10.1038/46965.
- Shu, D-G., Conway Morris, S., and Zhang, X-L. (November 1996). «A Pikaia-like chordate from the Lower Cambrian of China». Nature 384 (6605). doi:10.1038/384157a0.
- Ruppert, E. (2005). «Key characters uniting hemichordates and chordates: homologies or homoplasies?». Canadian Journal of Zoology 83: 8-23. doi:10.1139/z04-158
- Edgecombe G. D., Giribet G., Dunn C. W. et al. Higher-level metazoan relationships: recent progress and remaining questions (англ.) // Organisms Diversity & Evolution. — 2011. — Vol. 11, no. 2. — P. 151—172. — doi:10.1007/s13127-011-0044-4.
- Blair, J.E., and S. Blair Hedges, S.B. (2005). «Molecular Phylogeny and Divergence Times of Deuterostome Animals». Molecular Biology and Evolution 22 (11): 2275—2284. doi:10.1093/molbev/msi225.
- Perseke M, Hankeln T, Weich B, Fritzsch G, Stadler PF, Israelsson O, Bernhard D, Schlegel M. The mitochondrial DNA of Xenoturbella bocki: genomic architecture and phylogenetic analysis // Theory in Biosciences. — 2007. — Vol. 126, № 1. — P. 35-42. — doi:10.1007/s12064-007-0007-7. — PMID 18087755.
- Marlétaz F., Peijnenburg K. T. C. A., Goto T., Satoh N., Rokhsar D. S. A new spiralian phylogeny places the enigmatic arrow worms among gnathiferans (англ.) // Current Biology. — Cell Press, 2019. — Vol. 29, no. 2. — doi:10.1016/j.cub.2018.11.042. — PMID 30639106.
- Benton M. J. Vertebrate Paleontology. — Blackwell Science Ltd, 2005. — 472 с. — P. 389—403. — ISBN 978-0-632-05637-8.
- Бесчерепные — статья из Большой советской энциклопедии.
- Benton, M.J. Vertebrate Palaeontology: Biology and Evolution (англ.). — Blackwell Publishing, 2000. — P. 6. — ISBN 0632056142.
- Gee, H. Evolutionary biology: The amphioxus unleashed (англ.) // Nature. — 2008. — June (vol. 453, no. 7198). — P. 999—1000. — doi:10.1038/453999a. — . — PMID 18563145.
- Branchiostoma. Lander University. Дата обращения: 23 сентября 2008.
- Thaliacea в European Register of Marine Species
- Benton, M.J. Vertebrate Palaeontology: Biology and Evolution (англ.). — Blackwell Publishing, 2000. — P. 5. — ISBN 0632056142.
- Appendicularia (PDF) (недоступная ссылка). Australian Government Department of the Environment, Water, Heritage and the Arts. Дата обращения: 28 октября 2008. Архивировано 20 марта 2011 года.
- Morphology of the Vertebrates. University of California Museum of Paleontology. Дата обращения: 23 сентября 2008. Архивировано 27 мая 2012 года.
- Глотка — статья из Большой советской энциклопедии.
- Introduction to the Petromyzontiformes (недоступная ссылка). University of California Museum of Paleontology. Дата обращения: 28 октября 2008. Архивировано 24 января 2018 года.
- Introduction to the Myxini. University of California Museum of Paleontology. Дата обращения: 28 октября 2008. Архивировано 27 мая 2012 года.
- Shigehiro Kuraku, S., Hoshiyama, D., Katoh, K., Suga, H., and Miyata, T. Monophyly of Lampreys and Hagfishes Supported by Nuclear DNA-Coded Genes (англ.) // Journal of Molecular Evolution. — 1999. — December (vol. 49, no. 6). — P. 729—735. — doi:10.1007/PL00006595. — PMID 10594174.
- Tassia M. G., Cannon J. T., Konikoff C. E. et al. The global diversity of Hemichordata (англ.) // PLOS One. — Public Library of Science, 2016. — Vol. 11, no. 10. — doi:10.1371/journal.pone.0162564. — PMID 27701429.
- Данные о числе видов для России и всего мира на сайте Зоологического института РАН
- ZOOINT Part21
- Cowen, R. History of Life (неопр.). — 3rd. — Blackwell Science, 2000. — С. 412. — ISBN 063204444-6.
Литература
- Н. Н. Гуртовой, Б. С. Матвеев, Ф. Я. Дзержинский. Практическая зоотомия позвоночных (низшие хордовые, бесчелюстные, рыбы). — М.: Высшая школа, 1976. — С. 351.
- Хордовые // Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М. : Сов. энциклопедия, 1986. — С. 692. — 831 с. — 100 000 экз.
| Таксономия | |
|---|---|
| В библиографических каталогах |
