Xenoturbella
Xenoturbella (лат.) — род морских червеобразных животных, выделяемый в монотипические семейство Xenoturbellidae и подтип ксенотурбеллид[1] (Xenoturbellida) из типа Xenacoelomorpha. Включает 6 видов, которые встречаются в водах Атлантического и Тихого океанов. Для Xenoturbella характерно чрезвычайно простое строение: у них нет централизованной нервной системы, ануса, почек и кровеносной системы. Эволюционное происхождение рода Xenoturbella неясно, и на этот счёт существует несколько гипотез.
| Xenoturbella | ||||||||
|---|---|---|---|---|---|---|---|---|
 | ||||||||
| Научная классификация | ||||||||
|
Домен: Царство: Подцарство: Без ранга: Тип: Подтип: Ксенотурбеллиды (Xenoturbellida Bourlat et al., 2006) Семейство: Xenoturbellidae Westblad, 1949 Род: Xenoturbella |
||||||||
| Международное научное название | ||||||||
| Xenoturbella Westblad, 1949 | ||||||||
| Виды | ||||||||
|
||||||||
| ||||||||
История изучения
Первые экземпляры Xenoturbella были обнаружены в шведских водах Северного моря в 1915 году шведским биологом Сикстеном Боком (1884—1946), но описаны только в 1949 году Эйнаром Вестбладом под названием Xenoturbella bocki и отнесены к Acoelomorpha (тогда в составе плоских червей)[2].
Старые сборы этих двух исследователей, как оказалось, включали второй вид меньшего размера, который был описан только в 1999 году под именем Xenoturbella westbladi шведским зоологом Олле Исраэльссоном[3].
Описание
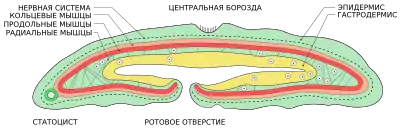
Xenoturbella — очень просто устроенные двусторонне-симметричные животные длиной 1–3 см (или даже 10 см, в зависимости от вида), обитающие в морском бентосе. Цвет от коричневатого до бледно-жёлтого, причём передняя часть тела беловатая, на многих экземплярах имеются чёрные пятна. Примерно посередине тело перехвачено бороздой, кроме того, имеются боковые борозды. В теле имеется два слоя эпителиальной ткани: состоящий из ресничных клеток внешний эпидермис и внутренний лишённый ресничек гастродермис, выстилающий простую мешковидную кишку. Реснички эпидермиса используются для передвижения. Ротовое отверстие располагается на брюшной стороне, немного впереди от центральной борозды. Анального отверстия нет[3], и непереваренные останки пищи выбрасываются наружу через рот[4]. Под базальной мембраной, подстилающей наружный эпителий, залегают два слоя мышц: снаружи слой кольцевых мышц, внутри слой продольных мышц[5].
Нервная система представлена сетью нейронов под эпидермисом, без каких-либо скоплений, формирующих ганглии и нервные стволы[6], хотя рядом с боковыми бороздами и вблизи статоциста нервная сеть слегка уплотняется. Боковые борозды могут выполнять сенсорную роль, но доподлинно это неизвестно[5]. Синапсы плохо развиты, зато распространена паракринная передача сигнала[7].
У Xenoturbella также отсутствуют дыхательная, кровеносная и выделительная системы. Помимо кишки, в теле нет каких-либо оформленных органов, за исключением статоциста, образованного ресничными клетками[3]. Морфологически статоцист Xenoturbella не имеет ничего общего со статоцистами других морских беспозвоночных животных. Неизвестно, точно ли он выполняет, как у других организмов, роль балансира; высказывается предположение, что он имеет эндокринные функции[5].
Размножение и развитие
Детали гаметогенеза и годичного цикла Xenoturbella неизвестны, хотя в оригинальном исследовании 1949 года сообщается о том, что популяция из Gullmarsfjord предпочтительнее размножается зимой. Недавно этот факт был подтверждён, но по-прежнему неизвестно, что служит стимулом для размножения в холодных тёмных водах[5].
Оформленные гонады отсутствуют, гаметы образуются и располагаются в различных местах внутри тела. Взрослые особи, продуцирующие сперму, встречаются редко, в отличие от яиц и эмбрионов[8]. Тем не менее, установлено, что сперматозоиды Xenoturbella схожи со сперматозоидами других примитивных Bilateria и состоят из слитной головки, не подразделённой на отделы, и жгутика с типичным для эукариот строением 9х2 + 2. Такой тип сперматозоидов встречается у разнообразных беспозвоночных, имеющих наружное оплодотворение. Яйца достигают 0,2 мм в диаметре, непрозрачные, бледно-оранжевого цвета. Личинки, вылупившиеся из яиц, покрыты ресничками, свободно плавают и стремятся держаться у поверхности воды. Они не имеют рта и кишки и живут только за счёт желтка[8], однако имеют нервные клетки, залегающие в базальной части эпителия, и мышечные клетки[9], ещё не организованные в продольные и кольцевые мышцы, как у взрослых. Личинки имеют мутный желтоватый цвет, что соответствует их лецитотрофности (питанию желтком). Таким образом, у Xenoturbella личинки лецитотрофные [10], и стадия питающейся личинки отсутствует. Неизвестно, способна ли Xenoturbella к бесполому размножению посредством фрагментации или почкования[5]. В развитии Xenoturbella имеется много общих черт с Acoelomorpha[11].
Распространение и экология
Ксенотурбеллиды населяют воды Атлантического и Тихого океанов. Два вида (Xenoturbella bocki и X. westbladi) найдены в Северном море у берегов Норвегии, Швеции и Шотландии и в Адриатическом море у берегов Хорватии. Остальные виды обнаружены у тихоокеанского побережья Мексики (в Калифорнийском заливе) и США (в заливе Монтерей)[3][12][13].
Об экологии Xenoturbella известно мало. Считается, что эти черви живут на поверхности донного ила или зарываются в него не слишком глубоко. Анализ с помощью изотопов азота показал, что Xenoturbella имеет высокий трофический уровень и, скорее всего, питается другими животными. Хотя рядом с этими червями обитает много других беспозвоночных животных, внутри них была обнаружена ДНК только двустворчатых моллюсков, поэтому, возможно, ксенотурбеллиды питаются исключительно двустворчатыми. Поскольку они не имеют специализированных органов для вскрытия раковин моллюсков, предполагается, что Xenoturbella питается ранними стадиями развития двустворчатых (яйцами, спермой, личинками и ювенильными особями), а также мёртвыми моллюсками, слизью моллюсков и их экскрементами. Эта гипотеза подтверждается находкой трохофороподобной личинки двустворчатого моллюска внутри взрослой ксенотурбеллиды. В то же время в экспериментальных условиях эти черви могли голодать месяцами, отказываясь от предложенной пищи, входящей в приведенный выше перечень. Возможно, они питаются очень редко и могут выживать в условиях голодания до года, а может быть, они поглощают растворённые органические вещества через эпидермис[5].
У Xenoturbella были идентифицированы два типа эндосимбиотических бактерий: в кишечнике многочисленны похожие на хламидий бактерии, а также гамма-протеобактерия Candidatus Endoxenoturbella lovénii[14]. Они также были найдены в скоплениях сперматид, что означает, что эндосимбиотические бактери у ксенотурбеллид могут передаваться вертикально, то есть от родителей к потомкам. Высказываются предположения, что бактерии обеспечивают способность к голоданию у ксенотурбеллид, а также могут быть ответственны за утрату ими ряда органов в ходе эволюции[5].
Филогения и эволюция
|
Систематическое положение этой группы было загадочным с самого её открытия. Сначала найденные животные идентифицировались как свободноживущие плоские черви. Позже их сближали с моллюсками[15], но оказалось, что ДНК Xenoturbella была загрязнена ДНК моллюсков, которыми Xenoturbella, по-видимому, питаются[8][16]. В 2003 году анализ ДНК Xenoturbella показал положение этого таксона как одного из наиболее примитивных представителей вторичноротых животных. В 2006 году предлагалось также выделить их в отдельный тип Xenoturbellida, сестринский к кладе Ambulacraria (включающей два типа: иглокожие + полухордовые)[17][18][19].
Впрочем, по своим морфологическим характеристикам (таким, как строение ресничного эпителия) род Xenoturbella очень близок к Acoelomorpha — другой малоизученной группе червеобразных организмов. Молекулярные данные указывают на то, что Xenoturbella и Acoelomorpha образуют монофилетическую группу, поэтому в настоящее время их объединяют в тип Xenacoelomorpha[11][20] (к нему также относят группу Nemertodermatida[21]). Несмотря на подтверждённую монофилию Xenacoelomorpha, их точное систематическое положение — в качестве базальной группы двусторонне-симметричных животных или же базальной группы вторичноротых — не определено до сих пор. Анализ транскриптома Xenoturbella profunda показал, что Xenoturbella — сестринская группа Acoelomorpha в составе Xenacoelomorpha, а Xenacoelomorpha, в свою очередь, — сестринская группа Nephrozoa, в которую включаются первичноротые и вторичноротые животные; следовательно, по данным этого анализа Xenacoelomorpha является базальной группой двусторонне-симметричных животных (Bilateria)[13].
Эволюционная история ксенотурбеллид может быть реконструирована по-разному в зависимости от того, каково их настоящее происхождение. Если они являются ранней ветвью Bilateria, то, скорее всего, упрощённый план тела без централизованной нервной системы и ануса был унаследован ими от последнего общего предка двусторонне-симметричных животных. Более того, возможно, не питающиеся однообразно покрытые ресничками личинки губок, стрекающих и Xenoturbella являются симплезоморфией Metazoa. Если же Xenoturbella и Acoelomorpha образуют один тип Xenacoelomorpha, сестринский по отношению к Ambulacraria в составе вторичноротых, то у Xenacoelomorpha вторично утрачены такие органы, как анус, метанефридии и жабры, которые, как предполагается, были у общего предка вторичноротых животных. Однако причины вторичного упрощения у Xenoturbella неясны, поскольку эти черви не являются сидячими, паразитическими или микроскопическими организмами[5].
Классификация
Xenoturbella по данным Rouse et al. (2016)[13]
| Xenoturbella |
| ||||||||||||||||||||||||||||||
Род Xenoturbella содержит 6 видов[12]:
- Xenoturbella bocki Westblad, 1949
- Xenoturbella churro Rouse et al., 2016
- Xenoturbella hollandorum Rouse et al., 2016
- Xenoturbella japonica Nakano, 2017[22]
- Xenoturbella monstrosa Rouse et al., 2016
- Xenoturbella profunda Rouse et al., 2016
- Xenoturbella westbladi Israelsson, 1999
На основе анализа ДНК в 2016 году былb исследованы филогенетические отношения ранее известных видов и выявлено, что таксоны X. monstrosa вместе с X. profunda и X. churro формируют общую кладу глубоководных видов (они достигают в длину 10 см и более и живут на глубинах до 1700 до 3700 м), а виды X. bocki и X. hollandorum (имеют длину до 4 см, обитают на глубинах до 650 м) составляют кладу мелководных видов[13]. В таблице ниже приводится краткое описание известных видов ксенотурбеллид.
| Вид | География | Описание[13] |
|---|---|---|
| Xenoturbella monstrosa | Побережье Калифорнии и Мексики | Тело длиной около 20 см, пурпурного или бледно-розового цвета. На дорсальной стороне тела имеется две продольные борозды. Рот овальной формы. Эпидермальная нервная сеть покрывает 2/3 вентральной поверхности. Гаметы залегают дорсально и вентрально |
| Xenoturbella churro | Побережье Калифорнии и Мексики | Тело 10 см длиной, оранжевое или розовое. На дорсальной стороне тела четыре глубокие продольные борозды. Передний конец тела округлый, задний заострённо-суживающийся. Рот овальный. Эпидермальная нервная сеть покрывает 1/3 вентральной поверхности. Гаметы залегают дорсально и вентрально |
| Xenoturbella profunda | Побережье Мексики | Тело до 15 см длиной, бледно-розового цвета, имеется эпидермис, кольцевые и продольные мышцы, паренхима и гастродермис. На дорсальной стороне две глубокие продольные борозды. Передний конец тела округлый, задний суживающийся. Рот овальный. Эпидермальная нервная сеть покрывает 1/3 вентральной поверхности. Гаметы залегают дорсально и вентрально. Ооциты достигают 450 мкм в диаметре, сперма в сферических головках |
| Xenoturbella hollandorum | Побережье Калифорнии | Тело короткое (2,5 см), ярко-розовое. На дорсальной стороне имеется пара продольных борозд. Рот ромбовидный. |
| Xenoturbella bocki | Западное побережье Швеции[3] | Медленно ползающее животное. Тело до 4 см длиной, бело-жёлтое с тёмными пятнышками, каждое из которых образовано скоплениями пигментных гранул прямо у поверхности. В передней части тела имеются две продольные сенсорные бороздки. Ооциты до 140 мкм в диаметре, с многочисленными микроядрами, покрыты толстой (12 мкм) оболочкой[3] |
| Xenoturbella westbladi | Побережье Швеции[3] | Быстро скользящее животное, тело до 12 мм длиной, розовое, с малочисленными тёмными пятнышками. Сперматозоиды соединены друг с другом, яйца мелкие (около 55 мкм в диаметре), с тонкой оболочкой. В период размножения у самок в гастродермисе формируются ресничные каналы, разделяющиеся после копуляции. Оплодотворение внутреннее, яйца развиваются в паренхиме[3] |
Примечания
- Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М. : Сов. энциклопедия, 1986. — С. 300. — 831 с. — 100 000 экз.
- Westblad E. Xenoturbella bocki n.g., n.sp., a peculiar, primitive turbellarian type // Arkiv för Zoologi. — Stockholm, 1949. — Vol. 1. — P. 3—29.
- Israelsson O. New light on the enigmatic Xenoturbella (phylum uncertain): ontogeny and phylogeny // Proceedings of the Royal Society B: Biological Sciences. — 1999. — 22 апреля (т. 266, № 1421). — С. 835—841. — ISSN 0962-8452. — doi:10.1098/rspb.1999.0713.
- McCafferty G. Deep-sea ‘purple sock’ provides clues to early life. // CNN (4 февраля 2016). Дата обращения: 6 мая 2016. Архивировано 7 мая 2016 года.
- Nakano H. What is Xenoturbella? (англ.) // Zoological letters. — 2015. — Vol. 1. — P. 22. — doi:10.1186/s40851-015-0018-z. — PMID 26605067.
- Perea-Atienza E., Gavilán B., Chiodin M., Abril J. F., Hoff K. J., Poustka A. J., Martinez P. The nervous system of Xenacoelomorpha: a genomic perspective // The Journal of Experimental Biology. — 2015. — Vol. 218, Pt. 4. — P. 618—628. — doi:10.1242/jeb.110379. — PMID 25696825.
- Raikova Olga I., Reuter M., Jondelius Ulf, Gustafsson Margaretha K. S. An immunocytochemical and ultrastructural study of the nervous and muscular systems of Xenoturbella westbladi (Bilateria inc. sed.) // Zoomorphology. — 2000. — 23 октября (т. 120, № 2). — С. 107—118. — ISSN 0720-213X. — doi:10.1007/s004350000028.
- Israelsson O., Budd G. E. Eggs and embryos in Xenoturbella (phylum uncertain) are not ingested prey // Development Genes and Evolution. — 2005. — Vol. 215, no. 7. — P. 358—363. — doi:10.1007/s00427-005-0485-x. — PMID 15818482.
- Evolutionary Developmental Biology of Invertebrates. Vol. 1. Introduction, Non-Bilateria, Acoelomorpha, Xenoturbellida, Chaetognatha / Ed. by A. Wanninger. — Wien: Springer Verlag, 2015. — P. 204. — vii + 251 p. — ISBN 978-3-7091-1861-0.
- Development of Xenoturbellida
- Nakano H., Lundin K., Bourlat S. J., Telford M. J., Funch P., Nyengaard J. R., Obst M., Thorndyke M. C. Xenoturbella bocki exhibits direct development with similarities to Acoelomorpha // Nature Communications. — 2013. — Vol. 4. — P. 1537. — doi:10.1038/ncomms2556. — PMID 23443565.
- Род Xenoturbella (англ.) в Мировом реестре морских видов (World Register of Marine Species). (Дата обращения: 24 июля 2016).
- Rouse G. W., Wilson N. G., Carvajal J. I., Vrijenhoek R. C. New deep-sea species of Xenoturbella and the position of Xenacoelomorpha // Nature. — 2016. — Vol. 530, no. 7588. — P. 94—97. — doi:10.1038/nature16545. — PMID 26842060.
- Kjeldsen K. U., Obst M., Nakano H., Funch P., Schramm A. Two types of endosymbiotic bacteria in the enigmatic marine worm Xenoturbella bocki. (англ.) // Applied and environmental microbiology. — 2010. — Vol. 76, no. 8. — P. 2657—2662. — doi:10.1128/AEM.01092-09. — PMID 20139320.
- Norén M., Jondelius U. Xenoturbella’s molluscan relatives // Nature. — 1997. — Vol. 390, no. 6655. — P. 31—32. — ISSN 0028-0836. — doi:10.1038/36242.
- Bourlat S. J., Nielsen C., Lockyer A. E., Littlewood D. T. J., Telford M. J. Xenoturbella is a deuterostome that eats molluscs // Nature. — 2003. — Vol. 424, no. 6951. — P. 925—928. — doi:10.1038/nature01851. — PMID 12931184.
- Bourlat S. J., Juliusdottir Thorhildur, Lowe C. J., Freeman R., Aronowicz J., Kirschner M., Lander E. S., Thorndyke M., Nakano H., Kohn A. B., Heyland A., Moroz L. L., Copley R. R., Telford M. J. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida // Nature. — 2006. — Vol. 444, no. 7115. — P. 85—88. — ISSN 0028-0836. — doi:10.1038/nature05241.
- Perseke M., Hankeln T., Weich B., Fritzsch G., Stadler P. F., Israelsson O., Bernhard D., Schlegel M. The mitochondrial DNA of Xenoturbella bocki: genomic architecture and phylogenetic analysis // Theory in Biosciences = Theorie in den Biowissenschaften. — 2007. — Vol. 126, no. 1. — P. 35—42. — doi:10.1007/s12064-007-0007-7. — PMID 18087755.
- Bourlat S. J., Rota-Stabelli O., Lanfear R., Telford M. J. The mitochondrial genome structure of Xenoturbella bocki (phylum Xenoturbellida) is ancestral within the deuterostomes // BMC Evolutionary Biology. — 2009. — Vol. 9. — P. 107. — doi:10.1186/1471-2148-9-107. — PMID 19450249.
- Hejnol A., Obst M., Stamatakis A., Ott M., Rouse G. W., Edgecombe G. D., Martinez P., Baguñà J., Bailly X., Jondelius U., Wiens M., Müller W. E., Seaver E., Wheeler W. C., Martindale M. Q., Giribet G., Dunn C. W. Assessing the root of bilaterian animals with scalable phylogenomic methods // Proceedings of the Royal Society. Ser. B. Biological Sciences. — 2009. — Vol. 276, no. 1677. — P. 4261—4270. — doi:10.1098/rspb.2009.0896. — PMID 19759036.
- Cannon J. T., Vellutini B. C., Smith J 3rd., Ronquist F., Jondelius U., Hejnol A. Xenacoelomorpha is the sister group to Nephrozoa. (англ.) // Nature. — 2016. — 4 February (vol. 530, no. 7588). — P. 89—93. — doi:10.1038/nature16520. — PMID 26842059.
- Nakano, Hiroaki; Miyazawa, Hideyuki; Maeno, Akiteru; Shiroishi, Toshihiko; Kakui, Keiichi; Koyanagi, Ryo; Kanda, Miyuki; Satoh, Noriyuki; Omori, Akihito; Kohtsuka, Hisanori. A new species of Xenoturbella from the western Pacific Ocean and the evolution of Xenoturbella (англ.) // BioMed Central : journal. — 2017. — Vol. 17, no. 1. — P. 245. — doi:10.1186/s12862-017-1080-2. — PMID 29249199.
Ссылки
- A PCR Survey of Xenoturbella bocki Hox Genes. (англ.)
- Movie of adult Xenoturbella bocki. (англ.)
- Ксенотурбеллиды оказались близки к предкам двустороннесимметричных животных.