Гребневики
Гребневики́ (лат. Ctenophora, от греч. κτείς — гребень, расческа и греч. φέρω — ношу) — широко распространённый тип морских животных. Отличительная особенность гребневиков — «гребни» из ресничек, используемые при плавании. Гребневики — самые большие среди передвигающихся при помощи ресничек животных: их размеры колеблются от нескольких миллиметров до полутора метров. Как и у стрекающих, их тела состоят из желеобразной массы, выстланной одним слоем клеток снаружи и ещё одним — изнутри; при этом эпителиальные слои имеют толщину в две клетки, а не одну, как у стрекающих. Ещё одно сходство этих двух типов животных — наличие децентрализованной нервной системы. Ранее обе группы объединяли в составе типа кишечнополостных (в ранге классов)[1], но впоследствии они были повышены в ранге до самостоятельных типов[2].
| Гребневики | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 «Ctenophorae», иллюстрация из работы Э. Геккеля «Красота форм в природе» (1904) | ||||||||||||
| Научная классификация | ||||||||||||
| Международное научное название | ||||||||||||
| Ctenophora Eschscholtz, 1829 | ||||||||||||
| Классы | ||||||||||||
|
||||||||||||
| ||||||||||||
Почти все гребневики — хищники. Их рацион включает разнообразных водных животных от микроскопических личинок и коловраток до небольших взрослых ракообразных. Исключением являются молодые представители двух видов, которые приспособленны к паразитированию на сальпидах. В благоприятных условиях гребневик может съедать в день в десять раз больше собственной массы. Официально известны лишь 100—150 видов, ещё около 25 не описаны и не названы. Наиболее распространённые представители гребневиков — цидипповые, обладатели яйцевидного тела и вытягивающихся щупалец, окруженных тентиллами с клейкими клетками — коллобластами. Внешний вид и образ жизни гребневиков разнообразны: этот тип включает и плоских глубоководных платиктенид, у которых взрослые особи лишены «гребней», и мелководных Beroe, лишенных щупалец и питающихся другими гребневиками. Подобный диморфизм позволяет эффективно сосуществовать сразу нескольким видам, так как вследствие разных типов охоты их экологические ниши не перекрываются.
Большинство видов — гермафродиты, притом встречаются как одновременные (одновременно производящие яйца и сперму), так и последовательные (производящие разные гаметы в разное время) в своем гермафродитизме формы. Оплодотворение, как правило, наружное, хотя для платиктенид характерно внутреннее оплодотворение и живорождение. Личинки, как правило, являются планктонными формами, похожими внешне на маленьких цидипповых, но в процессе онтогенеза их строение меняется, пока они не становятся похожими на взрослые формы. По крайней мере у нескольких видов личинки способны к неотении. Это, в сочетании с гермафродитизмом, позволяет популяциям быстро расти в числе.
Несмотря на мягкое тело, наиболее древние гребневики сохранились в палеонтологической летописи; древнейшие отпечатки относятся к раннему кембрию (около 525 миллионов лет назад). Положение гребневиков на филогенетическом древе долгое время было предметом споров; по последним данным, базирующимся на результатах молекулярной филогенетики, гребневики являются сестринским таксоном ко всем остальным животным. Принято считать, что последний общий предок всех современных гребневиков был похож на цидипповых и что все современные формы возникли уже после мел-палеогенового вымирания, то есть сравнительно недавно.
Анатомия и физиология
Внешний вид
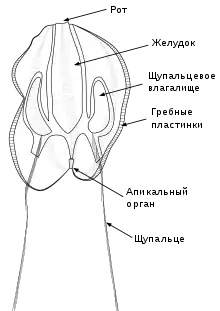
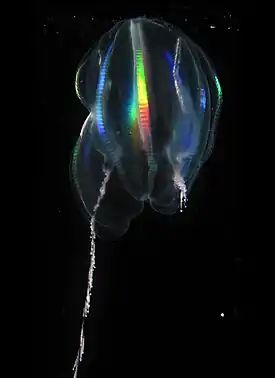
Размеры тела гребневиков варьируют от нескольких миллиметров до 30 см и более. Так, лентовидный венерин пояс (Cestum veneris) достигает в длину 1,5 м[3]. Общий план строения тела гребневиков можно представить следующим образом. Тело гребневика имеет два противолежащих полюса: оральный, на котором располагается рот, и аборальный, на котором находится особый аборальный орган, похожий на нервный ганглий. Органы в теле гребневика залегают радиально относительно орально-аборальной оси. От одного полюса к другому на поверхности гребневика тянется 8 продольных рядов довольно длинных ресничек (до 2 мм), которые позволяют гребневикам перемещаться со скоростью до 50 мм/с. Реснички располагаются компактными пластинчатыми группами — гребными пластинками, или ктенами, каждая из которых включает несколько тысяч ресничек. 8 рядов ктен начинаются от аборального полюса и тянутся к оральному, не доходя до ротового отверстия. Ктены совершают гребные движения, эффективный удар которых направлен в сторону аборального полюса, поэтому гребневики плывут ртом вперёд. В работающих гребных пластинках происходит дифракция света, из-за чего возникает эффект радуги. Реснички могут формировать некоторые другие специализированные органы, например, пейсмейкеры, задающие частоту биения пластинок, или своеобразные «зубы» (макроцилии) у Beroe[4].
На аборальной половине тела находятся два длинных щупальца, по одному с каждой стороны. Они берут начало в глубоких карманах, выстланных ресничным эпителием — щупальцевых влагалищах. Каждое щупальце снабжено боковым рядом тонких нитевидных выростов — тентилл. Щупальца способны сильно вытягиваться и сокращаться[4].
Симметрия тела гребневиков весьма своеобразна и является двухлучевой, то есть через тело гребневика можно провести две воображаемые взаимно перпендикулярные плоскости, каждая из которых разделит его на две идентичные половины (две плоскости симметрии). Первая такая плоскость — щупальцевая (тентакулярная), проходит вдоль орально-аборальной оси через оба щупальцевых влагалища. Вторая — глоточная (фарингеальная) плоскость, проходит вдоль орально-аборальной оси перпендикулярно к щупальцевой[4].
От описанного выше типового плана строения тела гребневиков у некоторых представителей имеются существенные отличия. Так, представители отряда Platyctenida имеют уплощённое тело, похожее на плоских червей или голожаберных моллюсков, лишённое гребных рядов[5]. У форм из отряда Beroida тело лишено щупалец[6].
Внутреннее строение

Значительная часть объёма тела гребневиков занята мезоглеей, несущей опорную функцию. Мезоглея представляет собой упругий гель, на 99 % состоящий из воды[7] и содержащий разнообразные клетки и коллаген. Рот ведёт в пищеварительную систему, называемую целентероном. Она начинается большой, уплощённой в одной плоскости глоткой (фаринксом), которая открывается в короткий канал, дающий начало дополнительным каналам, которые поставляют питательные вещества к гребным рядам, щупальцевым влагалищам и другим внутренним органам[4]. Мышечные и нервные клетки располагаются и в мезоглее, и в эпидермисе[8].
Стенка тела

Стенка тела гребневиков, а также стенка глотки и щупальцевых влагалищ образована эпидермисом, который подстилается мезоглеей и покрыт защитным слоем слизи. Эпидермис гребневиков выглядит как двухслойный эпителий с базальной пластинкой. Наружный слой образован ресничными клетками, причём их апикальный конец несёт не одну, а много ресничек; интерстициальными (промежуточными) клетками, функционирующими как стволовые; секреторными (железистыми) клетками, выделяющими слизь; чувствительными клетками нескольких типов и особыми клетками, присущими только гребневикам — коллоцитами, или коллобластами. По всей поверхности тела распределены только чувствительные и железистые клетки, другие клетки приурочены только к определёнными органам (гребными пластинкам, щупальцам, щупальцевым влагалищам). Внутренний слой эпителия образован миоэпителиальными клетками и нервными клетками, формирующими нервную сеть (нервный плексус). В мезоглее находятся разнообразные клетки: миоциты, нервные клетки, пигментные клетки и амебоидные клетки, которые формируют внеклеточный матрикс — основу мезоглеи[8].
В эпидермисе тентилл и щупалец имеются многочисленные специализированные клетки — коллоциты, которые выделяют клейкое вещество и приклеиваются к добыче. Коллоциты, как и стрекающие клетки, формируются в эпидермисе в ходе дифференцировки интерстициальных клеток. Зрелый коллоцит состоит из вздутой апикальной части (шапочки), которая несёт многочисленные везикулы с клейким веществом, и более узкой «ножки», которая заякоривает коллоцит в эпидермисе, мышечном слое или мезоглее. Вокруг «ножки» спирально обвивается специальная нить, по-видимому, гасящая механическую нагрузку на коллоцит. Вероятно, каждый коллоцит используется только один раз и после этого заменяется новым[9].
Гребневики не имеют собственных стрекательных клеток, однако Haeckelia rubra, имеющая щупальца без тентилл и коллобластов, может вставлять в свои щупальца стрекательные клетки съеденных гидрозоев[7].
Мышечная система
Мышечные элементы располагаются в эпидермисе и мезоглее. В эпидермисе мышцы могут быть представлены продольными и/или кольцевыми пучками в зависимости от видовой принадлежности гребневика. При хорошем развитии обоих видов пучков формируется мышечная сеть. Мышечные элементы мезоглеи представлены продольными, поперечными или радиальными миоцитами. За счёт работы мышц эпидермиса и мезоглеи у разных гребневиков могут осуществляться открывание и закрывание рта, изменение объёма глотки, втягивание аборального органа и гребных пластинок[10].
У подавляющего большинства гребневиков имеются только гладкие мышечные элементы. Поперечнополосатые мышцы обнаружены только у гребневика Euplokamis dunlapae и участвуют в раскручивании тентилл, необходимых для ловли добычи. Характерные для стрекающих эпителиально-мускульные клетки у гребневиков отсутствуют. Интересно, что миоциты, располагающиеся в мезоглее, окружены базальной пластинкой, что может говорить в пользу их эпителиального происхождения[11].
Гладкие мышцы гребневиков отличаются от гладких мышц двусторонне-симметричных животных. У гребневиков актиновые микрофиламенты крепятся только к мембране мышечной клетки, а у билатерий, например, плоских червей и моллюсков, они могут прикрепляться к некоторым белкам (например, α-актинину) прямо в цитоплазме[11].
Нервная система
Нервная система гребневиков представлена эпидермальной нервной сетью, образующей скопления нейронов в области сложных структур и органов (гребных рядов, глотки, щупалец, аборального органа). Нейроны также имеются в мезоглее, где иннервируют мышечные элементы. Главным нервным центром служит аборальный орган, хотя вокруг рта залегает нервное кольцо. Аборальный (апикальный) орган, по сути, представляет собой мозг простейшего строения, являющийся комбинацией рецепторных и эффекторных элементов. Аборальный орган имеет статоцист, расположенный на самом конце аборального канала пищеварительной системы. Его прозрачный купол сформирован длинными, неподвижными, изогнутыми ресничками. Под куполом находится шар, сложенный клетками с преломляющими свет гранулами. Снизу от статолита отходят 4 ряда пучка подвижных ресничек (балансира), расположенных на равном расстоянии друг от друга. Каждый балансир контролирует активность ресничек в двух смежных гребных рядах, то есть ресничек одного квадранта. От балансиров по направлению к оральному полюсу отходит по одной ресничной бороздке, каждая из которых далее делится на две ветви, соединяющиеся с двумя соседними гребными рядами. Частота биения ресничек квадранта определяется частотой биения соответствующего балансира. На дне статоциста имеется небольшое углубление, выстланное сенсорным эпителием. За пределами статоциста он представлен двумя узкими ресничными полосками — полярными полями, которые вытянуты в глоточной плоскости и расходятся от статоциста в противоположные стороны. Ресничные клетки полярных полей создают ток воды по направлению к статоцисту, другие клетки, скорее всего, выполняют сенсорные функции. У некоторых гребневиков, например, Beroe, по краю полярных полей в один ряд располагаются пальцевидные ветвящиеся папиллы. Аборальный орган, скорее всего, функционирует также как фото-, баро- и хеморецепторный орган[12].
В настоящее время многие учёные склоняются к версии о независимом происхождении нервной системы гребневиков, таким образом, нервная система (нейроны, синапсы, синаптическая передача) гребневиков и всех остальных животных может рассматриваться как результат конвергентной эволюции. В работе нервной системы многих гребневиков не задействовано большинство канонических низкомолекулярных нейромедиаторов других животных, а именно ацетилхолин, серотонин, дофамин, норадреналин, октопамин, гистамин, глицин). Впрочем, у некоторых гребневиков выявлены ацетилхолин и адреналин, причём у них эти нейромедиаторы задействованы в биолюминесценции. Передача сигнала в нейронах гребневиков осуществляется в основном посредством секреторных пептидов; это подтверждается наличием у них множества рецепторов, связанных с G-белками (GPCR). Описанные нейропептиды гребневиков не обнаруживают никакой гомологии с нейропептидами Metazoa. Нейропептиды гребневиков активируют ионные каналы, имеющие ворота, например, ионные каналы семейства DEG/ENaC. Многие гены, контролирующие развитие и судьбу нейронов у двусторонне-симметричных животных, у гребневиков отсутствуют. Они лишены ортологов генов, кодирующих многие важные белки синаптической передачи Metazoa, однако у них обнаружены специфические пресинаптические белки. У гребневиков имеются разнообразные иннексины, образующие щелевые контакты. Существует гипотеза, что гребневики утратили эти гены и низкомолекулярные нейромедиаторы и затем приобрели собственные механизмы передачи сигнала, однако она плохо согласуется с активным образом жизни гребневиков (активным животным было бы незачем утрачивать весьма эффективную систему передачи сигнала между нейронами, присущую другим животным)[13][14].
Вместе с тем секвенирование геномов гребневиков Mnemiopsis leidyi и Pleurobrachia bachei показало наличие у них многочисленных ионотропных глутаматных рецепторов, которые значительно отличаются от хорошо изученных ионотропных глутаматных рецепторов позвоночных (AMPA-, NMDA- и каинатных). Некоторые из них содержат солевой мостик между остатками аргинина и глутамата и способны связываться с глицином, что приводит к их активации. Выдвинуто предположение, что глутаматные рецепторы гребневиков и NMDA-рецепторы имеют общее происхождение, поэтому, возможно, у гребневиков глутамат и глицин могут функционировать как нейромедиаторы[15].
Пищеварительная система
Пищеварительная система гребевиков (целентерон) представлена несколькими отделами и каналами, по которым питательные вещества разносятся по телу. В типичном случае рот ведёт в сжатую с боков глотку, от которой начинается короткий аборальный канал. Начальный отдел аборального канала иногда называют желудком. Непосредственно под аборальным органом аборальный канал разделяется на 4 коротких анальных канала, охватывающих аборальный орган. Два анальных канала, расположенных напротив друг друга, открываются наружу маленькими анальными порами. К остальным органам отходят каналы, берущие начало от желудка. Среди них два глоточных канала, идущих вдоль плоских поверхностей глотки; два щупальцевых канала, снабжающих два щупальцевых влагалища; восемь меридиональных каналов, тянущихся под каждым из восьми гребных рядов; каждая гребная пластинка снабжена латеральным каналом. Меридиональные каналы могут заканчиваться слепо, а могут впадать в околоротовой кольцевой канал. У разных видов гребневиков расположение каналов может отличаться от описанной выше общей схемы[16].
Целентерон выстлан особым эпителием (гастродермисом). Ресничные клетки встречаются по всему гастродермису, однако они наиболее многочисленны в месте соединения глотки с аборальным каналом. Железистые клетки распространены по всему гастродермису. В области рта, помимо ресничных клеток, имеются хорошо развитые мышечные волокна[17].
Все каналы пищеварительной системы (за исключением аборального) находятся близко к поверхности тела или снабжаемому органу. Со стороны органа гастродермис образован высокими вакуолизированными питающими клетками, которые перемежаются с половыми клетками и фотоцитами, обеспечивающими биолюминесценцию. На противоположной стороне канала гастродермис образован плоским ресничным эпителием, пронизанным порами, причём каждая пора окружена двумя ресничными венчиками, каждый из которых образован шестью клетками. Реснички нижнего венчика обращены в просвет канала, а верхнего — в мезоглею. Это образование получило название ресничной розетки[17].
При проглатывании гребневиком добычи происходит её переваривание: мышцы глотки сокращаются, в её просвет выделяются пищеварительные ферменты. В глотке пища превращается в мелкодисперсную взвесь, которая легко переносится в желудок протоком жидкости, создаваемым ресничным продольными гребнями глотки. Неперевариваемые фрагменты пищи (например, экзоскелеты ракообразных) выбрасываются через рот, только некоторые выводятся через анальные поры. Из желудка пища благодаря работе ресничек распределяется по всем каналам. Пищевые частицы перемещаются в каналах в противоположных направлениях до тех пор, пока не будут захвачены питающими клетками, в которых происходит внутриклеточное переваривание. В них же, скорее всего, запасаются питательные вещества[18].
Выделение и осморегуляция
О механизмах выделения продуктов обмена у гребневиков известно мало. Аммиак, образующийся при деградации белков, по-видимому, удаляется через поверхность тела за счёт диффузии. Другие продукты обмена, скорее всего, выделяются из тела с жидкостью, заполняющей каналы целентерона, через рот или анальные поры[18].
Сохранение плавучести планктонными гребневиками при изменении плотности воды, скорее всего, объясняется их способностью к пассивному осмосу. Возможно, некоторую роль в этом играют и ресничные розетки, регулируя количество воды в мезоглее, которая выполняет гидростатическую функцию. Например, если морской гребневик с плотностью жидкости тела, равной плотности морской воды, попадёт в солоноватую воду эстуариев, то ресничные розетки будут активно гнать воду в мезоглею, чтобы уменьшить её плотность и увеличить объём. Если же адаптированный к солоноватой воде гребневик окажется в морской воде, то вода будет откачиваться из мезоглеи ресничными розетками в каналы пищеварительной системы и выводиться через рот[19].
Окраска и биолюминесценция
 Неклассифицированный вид Tortugas red: видны щупальца и тентиллы |
Гребневики, живущие у поверхности воды, обычно прозрачны и бесцветны. Глубоководные виды, напротив, имеют яркую пигментацию (например, ещё не классифицированный вид «красная Тортуга» (англ. Tortugas red)[20]). Платиктениды часто прикрепляются к другим бентосным организмам и имеют схожую с ними окраску. Целентерон представителей глубоководного рода Bathocyroe имеет яркий красный цвет: это нивелирует люминесценцию поглощённых гребневиком копепод[21].
Ресничные гребни планктонных форм создают подобия радуги, вызванные не люминесценцией, а преломлением света о вибрирующие реснички[22][23]. Тем не менее, почти все гребневики способны к биолюминесценции (исключения — род Pleurobrachia и некоторые другие), но излучаемый таким образом свет имеет синий либо зелёный оттенок и виден только в темноте[24].
Некоторые виды, например, Bathyctena chuni, Euplokamis stationis и Eurhamphaea vexilligera, при раздражении выделяют секрет (чернила), люминесцирующий при той же длине световой волны, что и их собственные тела. Молодые особи светятся ярче для своих размеров тела, чем взрослые, у которых люминесцентные клетки распределены равномерно по всему телу. Детальные исследования не смогли ни определить назначение биолюминесцентных систем у гребневиков, ни найти корреляцию между цветом испускаемого свечения и экологическими факторами (например, конкретной зоной обитания)[25].
В случае гребневиков биолюминесценция обусловлена наличием специальных клеток — фотоцитов, содержащих кальций-активируемые белки — фотопротеины. Примером фотопротеинов гребневиков может служить мнемиопсин — белок гребневика Mnemiopsis, испускающий синий свет из хромофора, который прочно, но нековалентно связан с центральным гидрофобным кором из 21 аминокислотного остатка[26]. У гребневиков был описан белок, близкий к зелёному флуоресцентному белку (GFP)[27]. Фотоциты нередко заключены внутри меридиональных каналов прямо под ресничными гребнями. В геноме Mnemiopsis leidyi находится десять генов фотопротеинов; все они экспрессируются в растущих фотоцитах совместно с генами опсина, что позволяет предполагать связь между биолюминесценцией и фототаксисом[28]. Гребневики биолюминесцируют по ночам, испуская вспышки света[3].
Биология
Движение
Многие гребневики (Mnemiopsis, Pleurobranchia и др.) плавают в основном за счёт гребных пластинок. Гребневики — самые крупные одиночные животные, использующие ресничный способ локомоции[3]. Впрочем, специализированные гребневики (Cestida, Platyctenida, некоторые Lobata) используют другие способы локомоции: плавают в толще воды или ползают за счёт волнообразных движений тела[29].
Механизм движения за счёт гребных пластинок таков. Работающие ктены совершают гребки от орального полюса к аборальному, однако при возобновлении движения после остановки, ктены начинают работать в обратном порядке: первыми активируются аборальные ктены, и от них зона работающих пластинок, как волна, распространяется к оральному полюсу. Активность ктен регулируется балансиром, причём генерируемый им импульс передаётся ктенам не по нервным тяжам, а вдоль ресничных бороздок. Считается, что у Cydippida в роли механического стимула работы ктен выступают возмущения в водной среде, вызываемые работающими ктенами. Проведение импульса между гребными пластинками осуществляется через межпластиночные ресничные бороздки[29].
Частота колебаний каждого из четырёх балансиров регулируется статолитом. Так, когда гребневик занимает вертикальное положение, то давление на статолиты одинаковое, а значит, и частота биения ктен одинакова. При смещении гребневика относительно вертикального положения давление на балансиры изменяется, поэтому частота биения гребных пластинок в разных рядах тоже различается[30].
Для Mnemiopsis leidyi показаны суточные вертикальные миграции[31].
Питание
Гребневики — активные хищники, питаются в основном планктонными ракообразными, медузами и другими гребневиками. Для ловли рачков используются щупальца и тентиллы, а у гребневиков, поедающих медуз и других гребневиков, щупальца, как правило, лишены тентилл, нередко они ловят добычу прямо ртом (например, лишённый щупалец Beroe). Для Pleurobranchia характерен способ питания, известный как захват с поворотом. Этот гребневик плавает ртом вперёд и тянет за собой щупальца, как невод. Если одного из щупалец коснётся маленький рачок, то он захватывается липкими тентиллами с коллоцитами. Далее щупальце сокращается, подтягивая добычу ко рту. Ктены соответствующей пары гребных рядов начинают работать в обратном направлении, в результате чего тело разворачивается так, чтобы рот оказался расположенным вплотную к добыче[18].
Размножение и развитие
Взрослым гребневикам свойственна регенерация[32], причём участки тела со статоцистами регенерируют быстрее, чем участки без них[33]. Однако только платиктениды способны к полноценному вегетативному размножению путём отделения небольших фрагментов по краям уплощённого тела. В отделившихся фрагментах начинается дифференцировка, и они становятся полноценными взрослыми животными[19].
Половые железы, или гонады, расположены на стенках меридиональных каналов целентерона. Гребневики — гермафродиты. Исключение составляют два вида рода Ocyropsis, представители которых однополы и не меняют пол в течение жизни. Яичники и семенники расположены в каждом из меридиональных каналов вдоль их стенок, обращённых к поверхности тела; таким образом, они тесно связаны с питающими клетками. Внутри яичника каждая яйцеклетка окружена клетками-«няньками», питающими её. Гаметы выметываются в ответ на световой стимул. Сначала гаметы поступают из гонады в короткие каналы, ведущие к гонопорам. Гонопоры образуют продольные ряды, причём каждая гонопора располагается между двумя соседними гребными пластинками. Нередко гаметы выводятся наружу через разрыв стенки[33]. У некоторых представителей рода Mnemiopsis зафиксировано самооплодотворение, для большинства же гребневиков характерно перекрёстное оплодотворение[34].
Для некоторых гребневиков (в основном видов отряда Lobata) характерна диссогония — размножение на двух стадиях жизни. Первоначально половые железы развиваются у цидиппидной личинки, которая образует более мелкие яйца, чем взрослые особи. Однако развиваются эти яйца нормально, из них выходит новое поколение. Затем половые железы личинки резорбируются и вновь образуются у взрослого гребневика. Созревание половых клеток продолжается до конца жизни или пока не начнётся дефицит пищи: в последнем случае гребневик сначала перестаёт вырабатывать гаметы, а затем уменьшается в размере. При восстановлении оптимальных условий размер и продуктивность приходят в норму. Это особенность позволяет гребневикам быстро восстанавливать численность популяций[22].
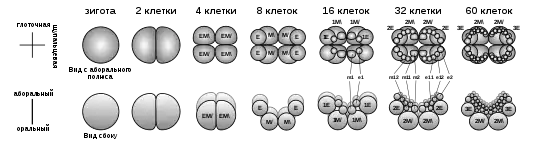
Яйца оплодотворяются в морской воде. В зиготах желток сконцентрирован в центре, а по периферии клетки находится цитоплазма с меньшим количеством желтка и ядро. Зигота претерпевает детерминированное дробление, то есть судьба каждого бластомера определяется ещё на самых ранних этапах дробления. Точка, в которой начинает формироваться первая борозда, соответствует оральному полюсу. Две первые взаимно перпендикулярные борозды дробления соответствуют будущим глоточной и щупальцевой плоскостям. Такое двулучевое дробление уникально для животного мира. В результате дальнейшего деления на оральном полюсе оказываются несколько макромеров, а на аборальном полюсе — многочисленные мелкие микромеры. Гаструляция идёт по типу эпиболии: аборальные микромеры перемещаются в сторону орального полюса, где погружаются в эмбрион путём инвагинации. Из них впоследствии развивается эпидермис, глотка, щупальцевые влагалища и гонады, то есть они имеют эктодермальное происхождение. Из макромеров отделяются микромеры орального полюса, которые представляют собой мезодерму. Они попадают в бластоцель через первичную кишку и дают начало миоцитам и другим клеткам мезодермы. После отделения оральных микромеров макромеры формируют энтодермальные каналы целентерона[35][33]. Главную роль в развитии и поддержании интерстициальных клеток играют гены семейства Sox[36].

Развитие гребневиков прямое, однако имеется планктонная ювенильная стадия, называемая цидиппидной личинкой. У Platyctenida эмбрионы вынашиваются в особых камерах внутри тела до стадии цидиппидной личинки[5]. Цидиппидная личинка похожа на миниатюрную Pleurobrachia с 8 гребными рядами и парой щупалец. На ранних стадиях развития гребневики имеют 4 гастральных канала. По мере развития цидиппидная личинка приобретает особенности организации, присущие половозрелым особям данной группы гребневиков. Так, у червеобразных бентосных Platyctenida личинка занимает иную экологическую нишу, чем взрослая ообь, и при превращении цидиппидных личинок во взрослых особей происходит настоящий метаморфоз[37]. Некоторые гребневики демонстрируют заботу о потомстве, например, у Tjalfiella яйца развиваются в выводковых сумках[33].
Развитие нервной и мышечной системы гребневиков имеет ряд уникальных особенностей. Так, у гребневика Pleurobrachia bachei мышечные элементы начинают развиваться на 1 день развития, а нервная система — лишь два дня спустя, непосредственно перед вылуплением полностью сформированной цидиппидной личинки. Первые нейроны возникают на аборальном полюсе на 3 день развития. Любопытно, что нейрогенез начинается после того, как у зародышей начинают проявляться различные поведенческие паттерны. Таким образом, для осуществления поведения зародышей, связанного с биением гребных пластинок и работой мышечных элементов, нет нужды в сформированных нейронах и их отростках[38].
Экология
Распространение
Гребневики широко распространены по всему миру и найдены во всех частях Мирового океана[22]. Тем не менее, наиболее изученными остаются прибрежные и мелководные планктонные формы (роды Pleurobrachia, Beroe и Mnemiopsis)[21][39]. Некоторые виды образуют массовые скопления в пелагиали и забивают рыбацкие сети, мешая прибрежному рыболовству (например, Pleurobranchia pileus в Северном море)[40].
Трофические связи

Почти все известные гребневики — хищники: известен лишь один паразитоидный род Lampeia, чьи личинки паразитируют на сальпидах, которыми питаются взрослые особи[21]. При обильном питании особь может каждый день съедать массу пищи, в 10 раз большую собственной массы[41]. В то время как представители рода Beroe питаются преимущественно другими гребневиками, большая часть нектоновых форм питается зоопланктоном разных размеров, начиная с личинок моллюсков и заканчивая небольшими ракообразными наподобие копепод, амфипод и криля[22]. Из-за разнообразия пищевых стратегий гребневиков нередко сравнивают с пауками. Эта дивергенция по методам охоты и удержания пищи и обусловливает разнообразие форм при сравнительно небольшом числе представителей[21]. Это также исключает перекрывание популяций нескольких видов по их экологическим нишам, например, виды рода Pleurobrachia, специализирующиеся на крупных копеподах, и рода Bolinopsis, питающиеся более мелкими коловратками и личинками ракообразных[42], могут достигать большой популяционной плотности на одной территории.
Гребневиков принято считать «тупиками» пищевых цепочек: низкое содержание питательных веществ делает их недостаточно привлекательными пищевыми объектами. Кроме того, остатки гребневиков в кишечнике хищника тяжело распознать, хотя «гребни» могут иногда сохраняться достаточно долго. Так, исследования кеты (Oncorhynchus keta) показывают, что рыбы переваривают гребневиков в 20 раз быстрее креветок той же массы и что в достаточном количестве гребневики могут послужить основой для питания. Гребневиками могут питаться мальки речных угрей[43]. Помимо рыб, гребневиками питаются медузы, морские черепахи и даже другие гребневики (род Beroe). Нередко они становятся жертвами организмов, питающихся медузами, ввиду общих циклов колебания численности популяций. На гребневиках также паразитируют личинки некоторых актиний и плоских червей[44].
Экологическая роль

Гребневики могут выступать в роли стабилизаторов морских экосистем, регулируя численность зоопланктона[45].
Наиболее известной экологической катастрофой, связанной с гребневиками, стала инвазия вида Mnemiopsis leidyi, завезённого кораблями вместе с балластными водами в экосистемы Чёрного и Азовского морей в 1980-х годах. Толерантный к малым перепадам солёности и температуры воды[46], этот вид нанёс большой урон, пожирая как зоопланктон, так и икру местных рыб[45]. Результатом, помимо прочего, стала эвтрофикация указанных водоёмов[47]. Положение ухудшалось отсутствием хищника, способного контролировать численность гребневика[46]. Уменьшению популяции M. leidyi способствовала интродукция североамериканского хищного гребневика Beroe ovata[48] и локальное похолодание 1991–93 гг.[46][47], однако восстановлению до первоначального уровня популяция зоопланктонных организмов в указанных морях, скорее всего, не подлежит[49].
В конце 1990-х M. leidyi появился в Каспийском море. Появление Beroe ovata ожидаемо снизило темп размножения гребневика. Примерно в то же время Mnemiopsis появился на востоке Средиземногого моря и сейчас угрожает Балтийскому и Северному морям[22].
Классификация
Точное количество ныне существующих видов гребневиков неизвестно; нередко представители одного и того же вида классифицируются как представители нескольких новых. Клаудия Миллс (англ. Claudia Mills) утверждает, что существует от 100 до 150 изученных видов, а также известны, но полностью не изучены представители ещё около 25 глубоководных видов[20].
Согласно современным представлениям, тип гребневиков подразделяется на следующие классы и отряды:
- Класс Tentaculata Eschscholtz, 1825 — гребневики с двумя щупальцами и сравнительно узкой глоткой[33].
- Подкласс Cyclocoela Ospovat, 1985
- Отряд Cambojiida Ospovat, 1985
- Отряд Cestidae Gegenbaur, 1856 — тропические и субтропические гребневики с прозрачным, бесцветным лентовидным или ремневидным телом[50].
- Отряд Cryptolobiferida Ospovat, 1985
- Отряд Ganeshida Moser, 1908 — два тропических вида, похожих на ювенильных особей Lobata[51].
- Отряд Lobata Eschscholtz, 1825 — планктонные формы с двумя мощными ротовыми лопастями, сильно редуцированными щупальцами и четырьмя аурикулями — выростами, на которые заходят оральные концы гребных рядов. Вместо типичных лопастевидных ктен — узкие треугольные группы ресничек[52].
- Отряд Thalassocalycida Madin & Harbison, 1978 — единственный вид Thalassocalyce inconstans, похожий на очень крупную гидроидную медузу[6].
- Подкласс Typhlocoela Ospovat, 1985
- Отряд Cydippida Gegenbaur, 1856 — планктонные сферические или овальные гребневики с хорошо развитыми щупальцами и щупальцевыми влагалищами. Между гребными пластинками нет ресничных бороздок[5].
- Отряд Platyctenida Bourne, 1900 — уплощённые бентосные формы, внешне похожие на плоских червей и голожаберных моллюсков. Лишены гребных рядов, но щупальца развиты хорошо и служат для захвата добычи. Семенники представлены отдельными мешочками, расположенными на поверхности каналов целентерона. Эмбрионы вынашиваются в особых каналах внутри тела[5].
- Подкласс Cyclocoela Ospovat, 1985
- Класс Nuda Chun, 1879

 Bathocyroe fosteri (отряд Lobata)
Bathocyroe fosteri (отряд Lobata) Pleurobrachia bachei (отряд Cydippida)
Pleurobrachia bachei (отряд Cydippida)_on_Echniaster_luzonicus_(Seastar).jpg.webp) Coeloplana astericola (отряд Platyctenida) на поверхности морской звезды
Coeloplana astericola (отряд Platyctenida) на поверхности морской звезды Beroe cucumis (отряд Beroida)
Beroe cucumis (отряд Beroida)
| |||||||||||||||||||||||||||||||||||||
Так как личинки всех гребневиков, кроме представителей класса бесщупальцевых, схожи между собой и напоминают цидипповых, принято считать, что последний общий предок современных форм также был похож на них, то есть имел овальное тело и одну пару втягиваемых щупалец. В 1985 Ричард Харбисон (англ. Richard Harbison) на основании исключительно морфологического анализа заключил, что отряд Cydippida не является монофилетической группой, так как разные семейства внутри отряда имеют большее сходства с представителями других отрядов, чем друг с другом. Харбисон также предположил, что общий предок современных гребневиков мог выглядеть и как бероиды[55]. Проведённый в 2001 году молекулярный анализ 26 видов, включая четыре на тот момент недавно открытых, подтвердил предположения относительно нарушенной монофилии цидипповых, однако заключил, что последний общий предок гребневиков всё же выглядел, как цидиппида. Кроме того, анализ выявил, что различия между геномами этих особей настолько незначительны, что родственные связи между отрядами Lobata, Cestida и Thalassocalycida до сих пор не удается определить. Из этого следует, что общий предок в эволюционных масштабах появился сравнительно недавно и, возможно, единственный из всех древних гребневиков пережил мел-палеогеновое вымирание. Включение в исследование анализ представителей других типов выявило, что, возможно, стрекающие куда ближе связаны с Bilateria, чем любой из этих таксонов с гребневиками[54].
Эволюция и филогения
Ископаемые формы


Гребневики не имеют твёрдых покровов, поэтому практически не оставляют ископаемых останков. Окаменелости, интерпретируемые как отпечатки тел гребневиков, как правило, остаются только в лагерштеттах, где фоссилизации подвергаются даже мягкие ткани (например, сланцы Бёрджес в Канаде). До середины 1990-х было найдено только два отпечатка; оба принадлежали представителям коронной группы из отложений раннего девона. Ещё три предполагаемых отпечатка гребневиков, найденные в сланцах Бёрджес и аналогичных по возрасту формациях Канады, датируются средним кембрием (примерно 505 миллионов лет назад). Все три представителя были лишены щупалец, но имели аномальное число ресничных гребней (от 24 до 80 против 8 у большинства современных представителей). У данных организмов также были подобия внутренних органов, отличные от любых структур современных гребневиков. Для одного ископаемого, найденного в 1996 году, характерен крупный рот, окруженный, по-видимому, крупной складкой, возможно, мускульной природы[56]. Обнаружение аналогичных фоссилий в Китае в 1997 году свидетельствует о том, что данные организмы были широко распространены в кембрийских морях, однако сильно отличались от современных гребневиков — так, например, у одного из найденных представителей ряды ресничек располагались на специфических выдающихся лопастях[57]. Эдиакарская Eoandromeda octobrachiata, жившая 580 миллионов лет назад, также могла быть древним гребневиком[58].
Вайяобразный организм Stromatoveris из юньаньских отложений (Китай), живший примерно 515 млн. лет назад и напоминающий вендобионтов, вполне мог быть биологическим прародителем гребневиков. По мнению ряда ученых (Shu, Conway Morris, et al.), отпечатки Stromatoveris несут следы фильтрационного аппарата, состоящего из циллий. Вероятно, гребневики произошли от сидячих организмов, впоследствии развивших ресничный аппарат в средство перемещения в толще воды[59].
Эволюционные связи с другими животными
Связи гребневиков с другими представителями царства важны для понимания ранней эволюции животных и появления многоклеточности. По различным версиям, приходятся сестринской группой Bilateria[60][61], стрекающим[62][63][64][65]или Bilateria, стрекающим и пластинчатым одновременно[66][67][68] либо сразу всем остальным типам животных[69][70]. Исследования гребневиков на наличие либо отсутствие конкретных групп генов (например, гомеобокса, а также микроРНК[14]) и молекулярных механизмов (таких, как сигнальный путь Wnt, ядерные рецепторы и натриевые каналы) говорят в пользу последних двух теорий[71][72][73][74]. Показано, что у Pleurobranchia отсутствуют ключевые элементы запуска врождённого иммунного ответа: рецепторы опознавания паттерна (Toll-подобные, Nod-подобные, RIG-подобные, домен Ig-Toll-IL-1R), иммунные медиаторы и транскрипционные факторы MyD88 and RHD, которые присутствуют у билатерий, книдарий и, в изменённом виде, у губок[75]. Последние результаты сравнения секвенированного генома гребневика Mnemiopsis leidyi с геномами других животных подтверждают сестринские отношения гребневиков и всех прочих Metazoa[76][77]. Из этого следует, что нервные и мышечные клетки либо исчезали в эволюционной истории остальных типов (например, у губок), либо появились у гребневиков совершенно независимо[76]. Впрочем, некоторые исследования опровергают сестринское положение гребневиков по отношению к другим животным[78].
Примечания
- Гребневики // Большая советская энциклопедия : в 66 т. (65 т. и 1 доп.) / гл. ред. О. Ю. Шмидт. — М. : Советская энциклопедия, 1926—1947.
- Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- Рупперт, Фокс, Барнс, 2008, с. 324.
- Рупперт, Фокс, Барнс, 2008, с. 325.
- Рупперт, Фокс, Барнс, 2008, с. 343.
- Рупперт, Фокс, Барнс, 2008, с. 346.
- Вестхайде, Ригер, 2008, с. 194.
- Рупперт, Фокс, Барнс, 2008, с. 329.
- Рупперт, Фокс, Барнс, 2008, с. 330.
- Рупперт, Фокс, Барнс, 2008, с. 330, 332.
- Рупперт, Фокс, Барнс, 2008, с. 332.
- Рупперт, Фокс, Барнс, 2008, с. 332—333.
- Moroz L. L. The genealogy of genealogy of neurons. (англ.) // Communicative & integrative biology. — 2014. — Vol. 7, no. 6. — P. e993269. — doi:10.4161/19420889.2014.993269. — PMID 26478767.
- Jékely G., Paps J., Nielsen C. The phylogenetic position of ctenophores and the origin(s) of nervous systems. (англ.) // EvoDevo. — 2015. — Vol. 6. — P. 1. — doi:10.1186/2041-9139-6-1. — PMID 25905000.
- Alberstein R., Grey R., Zimmet A., Simmons D. K., Mayer M. L. Glycine activated ion channel subunits encoded by ctenophore glutamate receptor genes. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2015. — Vol. 112, no. 44. — P. 6048—6057. — doi:10.1073/pnas.1513771112. — PMID 26460032.
- Рупперт, Фокс, Барнс, 2008, с. 335—336.
- Рупперт, Фокс, Барнс, 2008, с. 336.
- Рупперт, Фокс, Барнс, 2008, с. 338.
- Рупперт, Фокс, Барнс, 2008, с. 339.
- Mills, C.E. Phylum Ctenophora: list of all valid scientific names (май 2007). Дата обращения: 10 февраля 2009. Архивировано 16 августа 2013 года.
- Haddock S. H. Comparative feeding behavior of planktonic ctenophores. (англ.) // Integrative and comparative biology. — 2007. — Vol. 47, no. 6. — P. 847—853. — doi:10.1093/icb/icm088. — PMID 21669763.
- Mills, C.E. Ctenophores – some notes from an expert. Дата обращения: 5 февраля 2009. Архивировано 16 августа 2013 года.
- Welch V., Vigneron J. P., Lousse V., Parker A. Optical properties of the iridescent organ of the comb-jellyfish Beroë cucumis (Ctenophora). (англ.) // Physical review. E, Statistical, nonlinear, and soft matter physics. — 2006. — Vol. 73, no. 4 Pt 1. — P. 041916. — doi:10.1103/PhysRevE.73.041916. — PMID 16711845.
- Haddock S. H. D., Case, J. F. Not All Ctenophores Are Bioluminescent: Pleurobrachia (фр.) // Biological Bulletin : magazine. — 1995. — Vol. 189, no 3. — P. 356—362. — doi:10.2307/1542153. — . Архивировано 20 февраля 2009 года.
- Haddock, S. H. D., and Case, J. F. Bioluminescence spectra of shallow and deep-sea gelatinous zooplankton: ctenophores, medusae and siphonophores (англ.) // Marine Biology : journal. — 1999. — April (vol. 133). — P. 571—582. — doi:10.1007/s002270050497. Архивировано 26 октября 2005 года. Архивированная копия (недоступная ссылка). Дата обращения: 13 августа 2013. Архивировано 16 мая 2008 года.
- Mahdavi A., Sajedi R. H., Hosseinkhani S., Taghdir M. Hyperactive Arg39Lys mutated mnemiopsin: implication of positively charged residue in chromophore binding cavity. (англ.) // Photochemical & photobiological sciences : Official journal of the European Photochemistry Association and the European Society for Photobiology. — 2015. — Vol. 14, no. 4. — P. 792—800. — doi:10.1039/c4pp00191e. — PMID 25635518.
- Haddock S. H., Mastroianni N., Christianson L. M. A photoactivatable green-fluorescent protein from the phylum Ctenophora. (англ.) // Proceedings. Biological sciences / The Royal Society. — 2010. — Vol. 277, no. 1685. — P. 1155—1160. — doi:10.1098/rspb.2009.1774. — PMID 20018790.
- Schnitzler C. E., Pang K., Powers M. L., Reitzel A. M., Ryan J. F., Simmons D., Tada T., Park M., Gupta J., Brooks S. Y., Blakesley R. W., Yokoyama S., Haddock S. H., Martindale M. Q., Baxevanis A. D. Genomic organization, evolution, and expression of photoprotein and opsin genes in Mnemiopsis leidyi: a new view of ctenophore photocytes. (англ.) // BMC biology. — 2012. — Vol. 10. — P. 107. — doi:10.1186/1741-7007-10-107. — PMID 23259493.
- Рупперт, Фокс, Барнс, 2008, с. 333.
- Рупперт, Фокс, Барнс, 2008, с. 333—334.
- Haraldsson M., Båmstedt U., Tiselius P., Titelman J., Aksnes D. L. Evidence of diel vertical migration in Mnemiopsis leidyi. (англ.) // Public Library of Science ONE. — 2014. — Vol. 9, no. 1. — P. e86595. — doi:10.1371/journal.pone.0086595. — PMID 24466162.
- Martindale M. Q. The ontogeny and maintenance of adult symmetry properties in the ctenophore, Mnemiopsis mccradyi. (англ.) // Developmental biology. — 1986. — Vol. 118, no. 2. — P. 556—576. — PMID 2878844.
- Вестхайде, Ригер, 2008, с. 195.
- Рупперт, Фокс, Барнс, 2008, с. 339—340.
- Рупперт, Фокс, Барнс, 2008, с. 340—341.
- Schnitzler C. E., Simmons D. K., Pang K., Martindale M. Q., Baxevanis A. D. Expression of multiple Sox genes through embryonic development in the ctenophore Mnemiopsis leidyi is spatially restricted to zones of cell proliferation. (англ.) // EvoDevo. — 2014. — Vol. 5. — P. 15. — doi:10.1186/2041-9139-5-15. — PMID 24834317.
- Рупперт, Фокс, Барнс, 2008, с. 341.
- Norekian T. P., Moroz L. L. Development of neuromuscular organization in the ctenophore Pleurobrachia bachei. (англ.) // The Journal of comparative neurology. — 2016. — Vol. 524, no. 1. — P. 136—151. — doi:10.1002/cne.23830. — PMID 26105692.
- Haddock, S. H. D. A golden age of gelata: past and future research on planktonic ctenophores and cnidarians (англ.) // Hydrobiologia : journal. — 2004. — Vol. 530/531. — P. 549—556. — doi:10.1007/s10750-004-2653-9. Архивировано 26 октября 2005 года. Архивированная копия (недоступная ссылка). Дата обращения: 2 августа 2013. Архивировано 26 октября 2005 года.
- Вестхайде, Ригер, 2008, с. 192.
- Reeve M. R. and Walter M. A. Laboratory studies of ingestion and food utilization in lobate and tentaculate ctenophores (англ.) // Limnology and Oceanography : journal. — 1978. — Vol. 23, no. 4. — P. 740—751. — doi:10.4319/lo.1978.23.4.0740. Архивировано 16 июля 2011 года. Архивированная копия (недоступная ссылка). Дата обращения: 2 августа 2013. Архивировано 16 июля 2011 года.
- Costello, J. H. and Coverdale, R. Planktonic Feeding and Evolutionary Significance of the Lobate Body Plan within the Ctenophora (англ.) // Biological Bulletin : journal. — 1998. — Vol. 195, no. 2. — P. 247—248. — doi:10.2307/1542863. — . Архивировано 16 июня 2011 года.
- Bouilliart M., Tomkiewicz J., Lauesen P., De Kegel B., Adriaens D. Musculoskeletal anatomy and feeding performance of pre-feeding engyodontic larvae of the European eel (Anguilla anguilla). (англ.) // Journal of anatomy. — 2015. — Vol. 227, no. 3. — P. 325—340. — doi:10.1111/joa.12335. — PMID 26278932.
- Arai, M. N. Predation on pelagic coelenterates: a review (неопр.) // Journal of the Marine Biological Association of the United Kingdom. — 2005. — Т. 85. — С. 523—536. — doi:10.1017/S0025315405011458. (недоступная ссылка)
- Chandy S. T., Greene, C. H. Estimating the predatory impact of gelatinous zooplankton (англ.) // Limnology and Oceanography : journal. — 1995. — Vol. 40, no. 5. — P. 947—955. — doi:10.4319/lo.1995.40.5.0947. Архивировано 16 июля 2011 года. Архивированная копия (недоступная ссылка). Дата обращения: 2 августа 2013. Архивировано 16 июля 2011 года.
- Purcell, J. E., Shiganova, T. A., Decker, M. B., and Houde, E. D. The ctenophore Mnemiopsis in native and exotic habitats: U.S. estuaries versus the Black Sea basin (англ.) // Hydrobiologia : journal. — 2001. — Vol. 451. — P. 145—176. — doi:10.1023/A:1011826618539. Архивировано 6 марта 2009 года.
- Oguz, T., Fach, B., and Salihoglu, B. Invasion dynamics of the alien ctenophore Mnemiopsis leidyi and its impact on anchovy collapse in the Black Sea (англ.) // Journal of Plankton Research : journal. — 2008. — Vol. 30, no. 12. — P. 1385—1397. — doi:10.1093/plankt/fbn094.
- Bayha, K. M.; Harbison, R., McDonald, J. H., Gaffney, P. M. Preliminary investigation on the molecular systematics of the invasive ctenophore Beroe ovata (англ.) // Aquatic Invasions in the Black, Caspian, and Mediterranean Seas : journal. — 2004. — P. 167—175.
- Kamburska, L. Effects of Beroe cf ovata on gelatinous and other zooplankton along the Bulgarian Black Sea Coast // Aquatic Invasions in the Black, Caspian, and Mediterranean Seas (англ.) / Dumont, H., Shiganova, T. A., and Niermann, U.. — Springer Netherlands, 2006. — P. 137—154. — ISBN 978-1-4020-1866-4. — doi:10.1007/1-4020-2152-6_5.
- Рупперт, Фокс, Барнс, 2008, с. 345.
- Рупперт, Фокс, Барнс, 2008, с. 344—345.
- Рупперт, Фокс, Барнс, 2008, с. 343—344.
- Тип Ctenophora (англ.) в Мировом реестре морских видов (World Register of Marine Species).
- Podar M., Haddock S. H., Sogin M. L., Harbison G. R. A molecular phylogenetic framework for the phylum Ctenophora using 18S rRNA genes. (англ.) // Molecular phylogenetics and evolution. — 2001. — Vol. 21, no. 2. — P. 218—230. — doi:10.1006/mpev.2001.1036. — PMID 11697917.
- Harbison G. R. On the classification and evolution of the Ctenophora // The Origins and Relationships of Lower Invertebrates (англ.) / Conway Morris S., George J. D., Gibson R., and Platt H. M.. — Oxford University Press, 1985. — P. 78—100. — ISBN 0-19-857181-X.
- Conway Morris S., Collins, D. H. Middle Cambrian Ctenophores from the Stephen Formation, British Columbia, Canada (англ.) // Philosophical Transactions of the Royal Society B : journal. — London: The Royal Society, 1996. — March (vol. 351, no. 1337). — P. 279—308. — doi:10.1098/rstb.1996.0024. — .
- Conway-Morris S. The Cambrian "explosion" of metazoans and molecular biology: would Darwin be satisfied? (англ.) // The International journal of developmental biology. — 2003. — Vol. 47, no. 7—8. — P. 505—515. — PMID 14756326.
- Tang F., Bengtson S., Wang Y., Wang X. L., Yin C. Y. Eoandromeda and the origin of Ctenophora. (англ.) // Evolution & development. — 2011. — Vol. 13, no. 5. — P. 408—414. — doi:10.1111/j.1525-142X.2011.00499.x. — PMID 23016902.
- Shu D. G., Morris S. C., Han J., Li Y., Zhang X. L., Hua H., Zhang Z. F., Liu J. N., Guo J. F., Yao Y., Yasui K. Lower Cambrian vendobionts from China and early diploblast evolution. (англ.) // Science (New York, N.Y.). — 2006. — Vol. 312, no. 5774. — P. 731—734. — doi:10.1126/science.1124565. — PMID 16675697.
- Simon Conway Morris, Alberto M. Simonetta. The Early Evolution of Metazoa and the Significance of Problematic Taxa (англ.). — Cambridge University Press, 1991. — P. 308. — ISBN 0-521-11158-7.
- NIELSEN C. Cladistic analyses of the animal kingdom // Biological Journal of the Linnean Society. — 1996. — Апрель (т. 57, № 4). — С. 385—410. — ISSN 0024-4066. — doi:10.1006/bijl.1996.0023.
- Rudolf Leuckart. Ueber die Morphologie und die Verwandtschaftsverhältnisse der wirbellosen thiere. Ein Beitrag zur Charakteristik und Classification der thierischen Formen (нем.). — 1923. — ISBN 1-245-56027-1.
- Ernst Heinrich Philipp August Haeckel. Systematische Phylogenie Der Wirbellosen Thiere, Invertebrata, Part 2: Des Entwurfs Einer Systematischen Stammesgeschichte (нем.). — 1896. — ISBN 1-120-86850-5.
- Libbie Henrietta Hyman. The Invertebrates: Volume I, Protozoa Through Ctenophra (англ.). — McGraw-Hill Education, 1940. — ISBN 0-07-031660-0.
- Philippe H., Derelle R., Lopez P., Pick K., Borchiellini C., Boury-Esnault N., Vacelet J., Renard E., Houliston E., Quéinnec E., Da Silva C., Wincker P., Le Guyader H., Leys S., Jackson D. J., Schreiber F., Erpenbeck D., Morgenstern B., Wörheide G., Manuel M. Phylogenomics revives traditional views on deep animal relationships. (англ.) // Current biology : CB. — 2009. — Vol. 19, no. 8. — P. 706—712. — doi:10.1016/j.cub.2009.02.052. — PMID 19345102.
- Wallberg Andreas, Thollesson Mikael, Farris James S., Jondelius Ulf. The phylogenetic position of the comb jellies (Ctenophora) and the importance of taxonomic sampling // Cladistics. — 2004. — Декабрь (т. 20, № 6). — С. 558—578. — ISSN 0748-3007. — doi:10.1111/j.1096-0031.2004.00041.x.
- Collins A. G. Phylogeny of Medusozoa and the evolution of cnidarian life cycles // Journal of Evolutionary Biology. — 2002. — 30 апреля (т. 15, № 3). — С. 418—432. — ISSN 1010-061X. — doi:10.1046/j.1420-9101.2002.00403.x.
- Pick K. S., Philippe H., Schreiber F., Erpenbeck D., Jackson D. J., Wrede P., Wiens M., Alié A., Morgenstern B., Manuel M., Wörheide G. Improved phylogenomic taxon sampling noticeably affects nonbilaterian relationships. (англ.) // Molecular biology and evolution. — 2010. — Vol. 27, no. 9. — P. 1983—1987. — doi:10.1093/molbev/msq089. — PMID 20378579.
- Dunn C. W., Hejnol A., Matus D. Q., Pang K., Browne W. E., Smith S. A., Seaver E., Rouse G. W., Obst M., Edgecombe G. D., Sørensen M. V., Haddock S. H., Schmidt-Rhaesa A., Okusu A., Kristensen R. M., Wheeler W. C., Martindale M. Q., Giribet G. Broad phylogenomic sampling improves resolution of the animal tree of life // Nature. — 2008. — Vol. 452, no. 7188. — P. 745—749. — doi:10.1038/nature06614. — PMID 18322464.
- Hejnol A., Obst M., Stamatakis A., Ott M., Rouse G. W., Edgecombe G. D., Martinez P., Baguñà J., Bailly X., Jondelius U., Wiens M., Müller W. E., Seaver E., Wheeler W. C., Martindale M. Q., Giribet G., Dunn C. W. Assessing the root of bilaterian animals with scalable phylogenomic methods // Proceedings of the Royal Society. Ser. B. Biological Sciences. — 2009. — Vol. 276, no. 1677. — P. 4261—4270. — doi:10.1098/rspb.2009.0896. — PMID 19759036.
- Ryan J. F., Pang K., Mullikin J. C., Martindale M. Q., Baxevanis A. D. The homeodomain complement of the ctenophore Mnemiopsis leidyi suggests that Ctenophora and Porifera diverged prior to the ParaHoxozoa // EvoDevo. — 2010. — Vol. 1, no. 1. — P. 9. — doi:10.1186/2041-9139-1-9. — PMID 20920347.
- Reitzel A. M., Pang K., Ryan J. F., Mullikin J. C., Martindale M. Q., Baxevanis A. D., Tarrant A. M. Nuclear receptors from the ctenophore Mnemiopsis leidyi lack a zinc-finger DNA-binding domain: lineage-specific loss or ancestral condition in the emergence of the nuclear receptor superfamily? // EvoDevo. — 2011. — Vol. 2, no. 1. — P. 3. — doi:10.1186/2041-9139-2-3. — PMID 21291545.
- Pang K., Ryan J. F., Mullikin J. C., Baxevanis A. D., Martindale M. Q. Genomic insights into Wnt signaling in an early diverging metazoan, the ctenophore Mnemiopsis leidyi // EvoDevo. — 2010. — Vol. 1, no. 1. — P. 10. — doi:10.1186/2041-9139-1-10. — PMID 20920349.
- Liebeskind B. J., Hillis D. M., Zakon H. H. Evolution of sodium channels predates the origin of nervous systems in animals // Proc. Nat. Acad. Sci. USA. — 2011. — Vol. 108, no. 22. — P. 9154—9159. — doi:10.1073/pnas.1106363108. — PMID 21576472.
- Moroz L. L., Kocot K. M., Citarella M. R., Dosung S., Norekian T. P., Povolotskaya I. S., Grigorenko A. P., Dailey C., Berezikov E., Buckley K. M., Ptitsyn A., Reshetov D., Mukherjee K., Moroz T. P., Bobkova Y., Yu F., Kapitonov V. V., Jurka J., Bobkov Y. V., Swore J. J., Girardo D. O., Fodor A., Gusev F., Sanford R., Bruders R., Kittler E., Mills C. E., Rast J. P., Derelle R., Solovyev V. V., Kondrashov F. A., Swalla B. J., Sweedler J. V., Rogaev E. I., Halanych K. M., Kohn A. B. The ctenophore genome and the evolutionary origins of neural systems. (англ.) // Nature. — 2014. — Vol. 510, no. 7503. — P. 109—114. — doi:10.1038/nature13400. — PMID 24847885.
- Ryan J. F., Pang K., Schnitzler C. E., Nguyen A. D., Moreland R. T., Simmons D. K., Koch B. J., Francis W. R., Havlak P., Smith S. A., Putnam N. H., Haddock S. H., Dunn C. W., Wolfsberg T. G., Mullikin J. C., Martindale M. Q., Baxevanis A. D. The genome of the ctenophore Mnemiopsis leidyi and its implications for cell type evolution. (англ.) // Science (New York, N.Y.). — 2013. — Vol. 342, no. 6164. — P. 1242592. — doi:10.1126/science.1242592. — PMID 24337300.
- Borowiec M. L., Lee E. K., Chiu J. C., Plachetzki D. C. Extracting phylogenetic signal and accounting for bias in whole-genome data sets supports the Ctenophora as sister to remaining Metazoa. (англ.) // BMC genomics. — 2015. — Vol. 16. — P. 987. — doi:10.1186/s12864-015-2146-4. — PMID 26596625.
- Pisani D., Pett W., Dohrmann M., Feuda R., Rota-Stabelli O., Philippe H., Lartillot N., Wörheide G. Genomic data do not support comb jellies as the sister group to all other animals. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2015. — Vol. 112, no. 50. — P. 15402—15407. — doi:10.1073/pnas.1518127112. — PMID 26621703.
Литература
- Зоология беспозвоночных : в 2 т. / под ред. В. Вестхайде и Р. Ригера. — М. : Товарищество научных изданий КМК, 2008. — Т. 1: от простейших до моллюсков и артропод. — 512, [8] с. — ISBN 978-5-87317-491-1.
- Рупперт, Э. Э. Зоология беспозвоночных: Функциональные и эволюционные аспекты : в 4 т. / Э. Э. Рупперт, Р. С. Фокс, Р. Д. Барнс ; под ред. А. А. Добровольского и А. И. Грановича. — М. : Издательский центр «Академия», 2008. — Т. 1. — 496 с. — ISBN 978-5-7695-3493-5.
- Стрекающие, гребневики // Иллюстрированные определители свободноживущих беспозвоночных евразийских морей и прилегающих глубоководных частей Арктики. — М.-СПб., 2012. — Т. 3.
Ссылки
| Таксономия | |
|---|---|
| В библиографических каталогах |
| Класс Щупальцевые (Tentaculata) |
| |
|---|---|---|
| Класс Бесщупальцевые (Nuda) |
| |