NMDA-рецептор
NMDA-рецептор (NMDAR; НМДА-рецептор) — ионотропный рецептор глутамата, селективно связывающий N-метил-D-аспартат (NMDA).
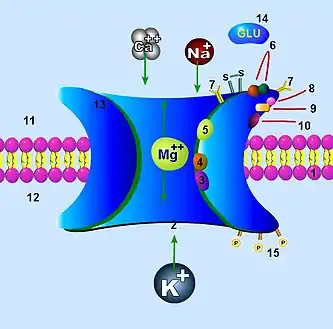
1. Клеточная мембрана
2. Канал, который блокируется магнием Mg2+ (3)
3. Участок блокировки Mg2+
4. Участок связывания галлюциногенов
5. Участок связывания цинка Zn2+
6. Участок связывания агонистов (глутамат) и антагонистов (APV)
7. Участки гликозилирования
8. Участки связывания протонов
9. Участки связывания глицина
10. Участок связывания полиаминов
11. Внеклеточное пространство
12. Внутриклеточное пространство
13. Комплексная (сложная) субъединица
Структурно NMDA-рецептор представляет собой полимер из 4 субъединиц — 2 из подтипа NR1 и 2 из подтипа NR2. В середине нулевых также был выделен новый класс субъединиц – NR3. В неактивированной форме канал рецептора закрыт ионом магния.
Ион магния удаляется при деполяризации постсинаптической мембраны, на которой находится рецептор. Одновременно с этим для функционирования рецептора должен поступить в синаптическую щель глутамат. Такая активация рецептора вызывает открытие ионного канала, селективного к катионам, что ведёт к притоку в клетку Na+ и, в небольшом объёме, Ca2+, а K+ покидает клетку. Ионы кальция, вошедшие через канал, активируют протеинкиназу CaMK-II. Происходит её аутофосфорилирование и фосфорилирование ряда белков нейрона-реципиента.
Данный процесс играет ключевую роль в синаптической пластичности, а следовательно и в процессах обучения и памяти. Также существует теория, что NMDA-рецепторы являются активаторами перехода REM-сон — Бодрствование; процесс опосредован СаМК-II[1]. В отличие от других рецепторов, NMDAR одновременно восприимчив к эндогенным лигандам-агонистам и антагонистам и к изменению мембранного потенциала (англ. voltage-dependent).
Альтернативный сплайсинг порождает множественные изоформы NR1, что вкупе с неоднородной экспрессией NR2 обусловливает различную структуру рецепторов в разных областях нервной системы.
В период эмбрионального и постнатального развития мозга отмечается изменение конструкции рецептора, в нём снижается число субъединиц NR2B, NR2D, NR3A, растёт число единиц NR2A и NR2C. Эти изменения различаются в разных областях мозга и подтипах нейронов, и происходят под действием разных факторов: так, в гранулярных клетках мозжечка замена NR2B на NR2C предположительно происходит под воздействием нейрегулинов. Множество работ посвящено возрастной смене преобладания NR2B на NR2A при созревании возбуждающих синапсов.[2]:163 Это изменение связано с действием рилина.[3][4][5]
См. также
- NMDA-антагонисты
- NR2B-субъединица
- Src — киназа, участвующая в сигнальном каскаде рецептора
- Анти-NMDA-рецепторный энцефалит
Примечания
- Петров, Гинниатулин. Нейробиология сна: современный взгляд.
- Antonius M. VanDongen. Biology of the NMDA Receptor (Frontiers in Neuroscience) (англ.). — Boca Raton: CRC, 2008. — ISBN 1-4200-4414-1.
- Sinagra M., Verrier D., Frankova D., Korwek K. M., Blahos J., Weeber E. J., Manzoni O. J., Chavis P. Reelin, very-low-density lipoprotein receptor, and apolipoprotein E receptor 2 control somatic NMDA receptor composition during hippocampal maturation in vitro (англ.) // J. Neurosci. : journal. — 2005. — June (vol. 25, no. 26). — P. 6127—6136. — doi:10.1523/JNEUROSCI.1757-05.2005. — PMID 15987942.
- Groc L., Choquet D., Stephenson F. A., Verrier D., Manzoni O. J., Chavis P. NMDA receptor surface trafficking and synaptic subunit composition are developmentally regulated by the extracellular matrix protein Reelin (англ.) // J. Neurosci. : journal. — 2007. — Vol. 27, no. 38. — P. 10165—10175. — doi:10.1523/JNEUROSCI.1772-07.2007. — PMID 17881522.
- Campo C. G., Sinagra M., Verrier D., Manzoni O. J., Chavis P. Reelin secreted by GABAergic neurons regulates glutamate receptor homeostasis (англ.) // PLoS ONE : journal. — 2009. — Vol. 4, no. 5. — P. e5505. — doi:10.1371/journal.pone.0005505. — PMID 19430527.
Ссылки
 Медиафайлы по теме NMDA-рецептор на Викискладе
Медиафайлы по теме NMDA-рецептор на Викискладе