Лиганд-зависимые ионные каналы
Лиганд-зависимые ионные каналы, лиганд-управляемые или лиганд-активируемые ионные каналы — также обычно называемые ионотропными рецепторами, представляют собой группу трансмембранных белков ионных каналов, которые позволяют ионам, например, Na+, K+, Ca2+ и/или Cl-, проходить через биологическую мембрану, посредством изменения конформации (окрытия) в ответ на связывание химического мессенджера (то есть лиганда), такого как, например, молекулы нейромедиатора[1][2][3].
| Нейромедиатор-зависимый ионный канал, трансмембранный регион | |
|---|---|
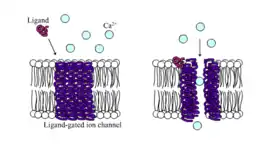 Лиганд-зависимый ионный канал | |
| Идентификаторы | |
| Символ | Neur_chan_memb |
| Pfam | PF02932 |
| InterPro | IPR006029 |
| PROSITE | PDOC00209 |
| SCOP | 1cek |
| SUPERFAMILY | 1cek |
| TCDB | 1.A.9 |
| OPM superfamily | 14 |
| OPM protein | 2bg9 |
| Доступные структуры белков | |
| Pfam | структуры |
| PDB | RCSB PDB; PDBe; PDBj |
| PDBsum | 3D-модель |
Ионные каналы с цистеиновой петлёй
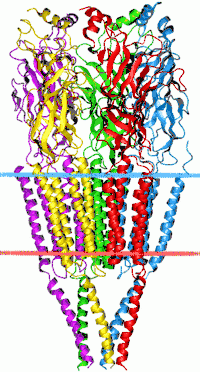
Цис-петлевые рецепторы названы в честь характерного цикла, который образован дисульфидной связью между двумя остатками молекул цистеина в N-концевом внеклеточном домене. Они являются частью большого семейства пентамерных лиганд-ионных каналов, которые обычно не имеют данной дисульфидной связи, поэтому их условное название «рецепторы Pro-loop»[4][5].
Структура
Цис-петлевые рецепторы имеют структурные элементы, которые высококонсервативны, причём большой внешнеклеточный домен (ECD) содержит альфу-спираль и 10 бета-тяжей. Следующие за ECD четыре трансмембранных сегмента (TMS) связаны внутриклеточными и внешнеклеточными структурами петли[6]. За исключением петли TMS 3-4, их длина составляет всего 7-14 остатков. Петля TMS 3-4 образует наибольшую часть внутриклеточного домена (ICD), при этом она является наиболее вариабельной (изменчивой) областью между всеми этими гомологичными рецепторами. ICD определяется петлёй TMS 3-4 вместе с петлей TMS 1-2, предшествующей поре ионного канала[6]. Кристаллизация рецепторов выявила структуры для некоторых членов семейства, но для обеспечения возможности кристаллизации внутриклеточную петлю обычно заменяли коротким линкером, присутствующим в прокариотических цис-петлевых рецепторах, поэтому их структуры не известны. Тем не менее, эта внутриклеточная петля, по-видимому, функционирует при десенсибилизации, модуляции физиологии канала лекарственными веществами и посттрансляционных модификациях. В нём присутствуют мотивы важные для движения, и ICD взаимодействует с белками каркаса, обеспечивая ингибирующее образование синапсов[6].
Прототипный лиганд-зависимый ионный канал представляет собой никотиновый ацетилхолиновый рецептор. Он состоит из пентамерных белковых субъединиц (обычно ααβγδ), включая два сайта связывания ацетилхолина (по одному на границе раздела каждой альфа-субъединицы). В нормальных физиологических условиях рецептору нужно ровно две молекулы ацетилхолина, чтобы открыть канал[7]. Открытие канала позволяет положительно заряженным ионам перемещаться по нему; в частности, ионы натрия (Na+) входят во внутрь клетки, а ионы калия выходят (К+).
Глициновые рецепторы
Глициновый рецептор (сокращенно GlyR или GLR) является рецептором аминокислоты-нейромедиатора — глицина. GlyR является цис-петлевым ионотропным рецептором, который оказывает своё действие посредством потока ионов хлора (Cl-). Он является одним из наиболее широко распространённых ингибиторных (тормозных) рецепторов в центральной нервной системе и играет важную роль в различных физиологических процессах, особенно в обеспечении торможения нейротрансмиссии в спинном мозге и стволе мозга[8].
Рецептор может активироваться рядом простых аминокислот, включая глицин, β-аланин и таурин, и может избирательно блокироваться конкурентным антагонистом с высоким сродством — стрихнином[9]. Кофеин является конкурентным антагонистом GlyR[10].
Рецепторы этого семейства расположены в виде пяти субъединиц (пентамеров), окружающих центральную пору, причём каждая субъединица состоит из четырёх α-спиральных трансмембранных сегментов[11]. В настоящее время известно четыре изоформы α-субъединицы (α1-4) GlyR, которые необходимы для связывания лигандов (GLRA1, GLRA2, GLRA3, GLRA4) и одной β-субъединицы (GLRB).
Типы катионных цис-петлевых рецепторов
| Тип | Класс | Название белков согласно рекомендации ИЮФАР[12] | Ген | Предыдущие названия |
|---|---|---|---|---|
| Серотониновые рецепторы (5-HT) |
5-HT3 | 5-HT3A 5-HT3B 5-HT3C 5-HT3D 5-HT3E |
HTR3A HTR3B HTR3C HTR3D HTR3E |
5-HT3A 5-HT3B 5-HT3C 5-HT3D 5-HT3E |
| Никотиновый ацетилхолиновый рецептор (nAChR) |
альфа | α1 α2 α3 α4 α5 α6 α7 α9 α10 |
CHRNA1 CHRNA2 CHRNA3 CHRNA4 CHRNA5 CHRNA6 CHRNA7 CHRNA9 CHRNA10 |
ACHRA, ACHRD, CHRNA, CMS2A, FCCMS, SCCMS |
| бета | β1 β2 β3 β4 |
CHRNB1 CHRNB2 CHRNB3 CHRNB4 |
CMS2A, SCCMS, ACHRB, CHRNB, CMS1D EFNL3, nAChRB2 | |
| гамма | γ | CHRNG | ACHRG | |
| дельта | δ | CHRND | ACHRD, CMS2A, FCCMS, SCCMS | |
| эпсилон | ε | CHRNE | ACHRE, CMS1D, CMS1E, CMS2A, FCCMS, SCCMS | |
| Цинк-активируемые ионные каналы (ZAC) |
ZAC | ZACN | ZAC1, L2m LICZ, LICZ1 |
Типы анионных цис-петлевых рецепторов
| Тип | Класс | Название белков согласно рекомендации ИЮФАР[12] | Ген | Предыдущие названия |
|---|---|---|---|---|
| ГАМКA | альфа | α1 α2 α3 α4 α5 α6 |
GABRA1 GABRA2 GABRA3 GABRA4 GABRA5 GABRA6 |
EJM, ECA4 |
| бета | β1 β2 β3 |
GABRB1 GABRB2 GABRB3 |
ECA5 | |
| гамма | γ1 γ2 γ3 |
GABRG1 GABRG2 GABRG3 |
CAE2, ECA2, GEFSP3 | |
| дельта | δ | GABRD | ||
| эпсилон | ε | GABRE | ||
| пи | π | GABRP | ||
| тета | θ | GABRQ | ||
| ро | ρ1 ρ2 ρ3 |
GABRR1 GABRR2 GABRR3 |
ГАМКC[13] | |
| Глициновый рецептор (GlyR) |
альфа | α1 α2 α3 α4 |
GLRA1 GLRA2 GLRA3 GLRA4 |
STHE |
| бета | β | GLRB |
Глутаматные ионотропные рецепторы
Ионотропные глутаматные рецепторы связывают молекулы нейромедиатора — глутамат. Они образуют тетрамеры с каждой субъединицей, состоящей из внешнеклеточного амино-концевого домена (ATD, в котором происходит тетрамерная сборка), внешнеклеточный лиганд-связывающий домен (LBD, который связывает глутамат) и трансмембранный домен (TMD, который образует ионный канал). Трансмембранный домен каждой субъединицы содержит три трансмембранные спирали, а также полумембранную спираль с реентерабельной петлёй. Структура белка начинается с ATD на N-конце, за которым следует первая половина LBD, которая прерывается спиралями 1,2 и 3 TMD перед продолжением второй половины LBD, а затем заканчивается спиралью 4 TMD на C-конце[14]. Это означает, что между TMD и внеклеточными доменами существует три связи. Каждая субъединица тетрамера имеет сайт связывания для глутамата, образованного двумя секциями LBD, образующими форму с раскладушкой. Только два из этих сайтов в тетрамере должны быть заняты, чтобы открыть ионный канал. Пора в основном образована половинной спиралью 2, которая по структуре напоминает перевернутый калиевый канал.
| Тип | Класс | Название белков согласно рекомендации ИЮФАР[12] | Ген | Предыдущее название |
|---|---|---|---|---|
| AMPA-рецептор | GluA | GluA1 GluA2 GluA3 GluA4 |
GRIA1 GRIA2 GRIA3 GRIA4 |
GLUA1, GluR1, GluRA, GluR-A, GluR-K1, HBGR1 GLUA2, GluR2, GluRB, GluR-B, GluR-K2, HBGR2 GLUA3, GluR3, GluRC, GluR-C, GluR-K3 GLUA4, GluR4, GluRD, GluR-D |
| Каинатный рецептор | GluK | GluK1 GluK2 GluK3 GluK4 GluK5 |
GRIK1 GRIK2 GRIK3 GRIK4 GRIK5 |
GLUK5, GluR5, GluR-5, EAA3 GLUK6, GluR6, GluR-6, EAA4 GLUK7, GluR7, GluR-7, EAA5 GLUK1, KA1, KA-1, EAA1 GLUK2, KA2, KA-2, EAA2 |
| NMDA-рецептор | GluN | GluN1 NRL1A NRL1B |
GRIN1 GRINL1A GRINL1B |
GLUN1, NMDA-R1, NR1, GluRξ1 |
| GluN2A GluN2B GluN2C GluN2D |
GRIN2A GRIN2B GRIN2C GRIN2D |
GLUN2A, NMDA-R2A, NR2A, GluRε1 GLUN2B, NMDA-R2B, NR2B, hNR3, GluRε2 GLUN2C, NMDA-R2C, NR2C, GluRε3 GLUN2D, NMDA-R2D, NR2D, GluRε4 | ||
| GluN3A GluN3B |
GRIN3A GRIN3B |
GLUN3A, NMDA-R3A, NMDAR-L, chi-1 GLU3B, NMDA-R3B | ||
| ‘Орфанный’ (рецептор-сирота) | (GluD) | GluD1 GluD2 |
GRID1 GRID2 |
GluRδ1 GluRδ2 |
АМРА-рецептор
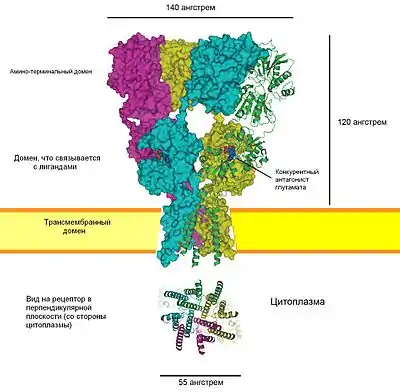
Рецептор α-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты (также известный как рецептор AMPA или рецептор квисквалята) представляет собой ионотропный трансмембранный рецептор глутамата не-NMDA-типа, который опосредует быструю синаптическую передачу в центральной нервной системе (ЦНС). AMPA-рецепторы обнаружены во многих частях мозга и являются наиболее часто встречаемым рецепторами в нервной системе. Тетрамер AMPA-рецептора GluA2 (GluR-2) был первым из ионных каналов глутаматного рецептора, который удалось кристаллизовать[15].
Лиганды
- Агонисты: глутамат, AMPA, 5-фторвиллардин, домоевая кислота, квискваловая кислота и др.
- Антагонисты: CNQX, Кинуреновая кислота, NBQX, перампанель, пирацетам и др.
- Положительные аллостерические модуляторы: анирацетам, циклотиазид, CX-516, CX-614 и др.
- Отрицательные аллостерические модуляторы: этанол, перампанель, талампанель, GYKI-52,466 и др.
Рецепторы NMDA
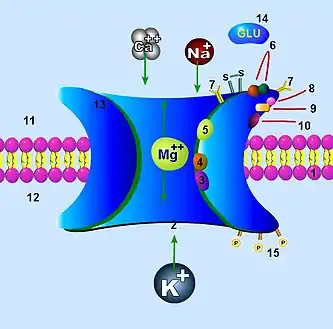
1. Клеточная мембрана
2. Канал, который блокируется магнием Mg2+ (3)
3. Участок блокировки Mg2+
4. Участок связывания галлюциногенов
5. Участок связывания цинка Zn2+
6. Участок связывания агонистов (глутамат) и антагонистов (APV)
7. Участки гликозилирования
8. Участки связывания протонов
9. Участки связывания глицина
10. Участок связывания полиаминов
11. Внеклеточное пространство
12. Внутриклеточное пространство
13. Комплексная (сложная) субъединица
N-метил-D-аспартатный рецептор (NMDA-рецептор) — один из типов ионотропного глутаматного рецептора, является потенциал-зависимым лиганд-ионным каналом, который активируется одновременным связыванием глутамата и коагониста (им являются молекулы D-серина или глицина)[16]. Исследования показывают, что NMDA-рецептор участвует в регулировании синаптической пластичности и памяти[17][18].
Когда NMDA-рецептор активируется одновременным связыванием двух коагонистов, открывается катионный канал, позволяя ионам Na+ и Ca2+ проникать в клетку, в свою очередь происходит повышение трансмембранного потенциала клетки. Таким образом, NMDA-рецептор является возбуждающим рецептором. При потенциалах покоя связывание двухвалентных ионов Mg2+ или Zn2+ происходит на внеклеточных сайтах связывания рецептора, это приводит к блокированию потока ионов, который протекает через ионный канал NMDA-рецептора. Однако, когда нейроны деполяризуются, например, путём интенсивной активации колокализованных постсинаптических AMPA-рецепторов, потенциал-зависимый блок Mg2+ частично ослабляется, что позволяет увеличить приток ионов через активированные NMDA-рецепторы. Результирующий приток ионов Ca2+ может запускать различные внутриклеточные сигнальные каскады, которые могут в конечном итоге изменить функции нейронов посредством активации различных киназ и фосфатаз[19].
Лиганды
- Первичные эндогенные коагонисты: глутамат и D-серин или глицин
- Другие агонисты: аминоциклопропанкарбоновая кислота; Д-циклосерин; L-аспартат; хинолинат и др.
- Частичные агонисты: N-метил-D-аспарагиновая кислота (NMDA); NRX-1074; 3,5-дибром-L-фенилаланин, и др[20].
- Антагонисты: кетамин, фенциклидин, декстропропоксифен, кетобемидон, трамадол, кинуреновая кислота (эндогенная) и др.
Каинатные рецепторы
Каинатные рецепторы, или рецепторы каиновой кислоты (KAR), представляют собой ионотропные рецепторы, которые могут активироваться под действием молекул нейромедиатора глутамата. Изначально они были идентифицированы как особый тип рецептора посредством их селективной активации агонистом каината, лекарственного средства, выделенного из клеток красной водоросли Digenea simplex. Они традиционно классифицируются как рецептор не-NMDA-типа, наряду с рецептором AMPA. KAR недостаточно изучены, вследствие меньшего распространения в головном мозге, чем AMPA и NMDA-рецепторы, или другие ионотропные глутаматные рецепторы. Постсинаптические каинатные рецепторы участвуют в возбуждающей нейротрансмиссии. Пресинаптические каинатные рецепторы участвуют в ингибирующей нейротрансмиссии путём модулирования высвобождения тормозного нейромедиатора ГАМК посредством пресинаптического механизма (пресинаптическое торможение).
Каинатный рецептор состоит из четырёх субъединиц, сходных с субъединицами AMPA- и NMDA-рецепторов. Всего известно 5 типов этих субъединиц: GluR5, GluR6, GluR7, KA1 и KA2[21].
Ионный канал, образованный каинатными рецепторами, проницаем для ионов натрия и калия. Проводимость каналов каинатного рецептора по одному каналу аналогична проводимости каналов AMPA, около 20 пикосименс (2*10-11 См). Тем не менее, нарастание и спад постсинаптических потенциалов порожденных каинатным рецептором происходит медленнее, чем для АМРА-рецептора. Проницаемость для кальция обычно очень небольшая, но варьируеться в зависимости от особенностей сегмента М2[22].
Лиганды
Агонисты:
- 5-Йодовилардин
- Домоевая кислота
- Глутаминовая кислота (глутамат) — эндогенный агонист
- Каиновая кислота — синтетический агонист, в честь которого назван рецептор
- LY-339434
- SYM-2081
Антагонисты:
- CNQX
- DNQX
- Этанол — неселективный
- NS102
- Кинуреновая кислота — эндогенный лиганд
- Тезампанель — также антагонист AMPA-рецептора
- UBP-302
- Теанин
Рецепторы ГАМК
ГАМК-рецепторы — группа клеточных рецепторов, эндогенным агонистом которых является γ-аминомасляная кислота (ГАМК), основной тормозной нейромедиатор в нервной системе позвоночных, экспрессируются в интернейронах коры головного мозга животных и человека. Существует 3 типа ГАМК-рецепторов, два из которых являются ионотропными — ГАМКА и ГАМКС, а один метаботропный — ГАМКB. Быстро реагирующие ГАМК-рецепторы являются членами семейства лиганд-управляемых ионных каналов с цистеиновой петлёй[23][24][25].
ГАМКА-рецептор
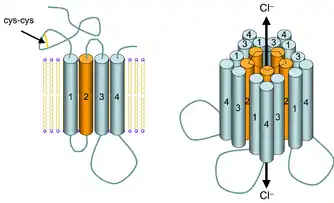
ГАМКА-рецепторы представляют собой лиганд-управляемые анионные каналы. ГАМК (гамма-аминомасляная кислота), эндогенный лиганд для данного типа рецепторов, является основным ингибирующим нейромедиатором в центральной нервной системе. При активации он опосредует поступление ионов хлора Cl– в нейрон, при этом происходит гиперполяризация мембраны клетки. ГАМКА-рецепторы встречаются во всех организмах, имеющих нервную систему. Благодаря их широкому распространению в нервной системе млекопитающих, они играют роль практически во всех функциях, выполняемых головным мозгом[26].
Различные лиганды могут специфически связываться с ГАМКА-рецепторами, активируя или ингибируя хлоридный канал Cl–.
Лиганды:
- Агонисты: ГАМК, мускимол, прогабид, габоксадол
- Антагонисты: бикукулин, габазин
- Частичный агонист: пиперидин-4-сульфокислота.
ГАМКC-рецептор
Рецептор ГАМКA-ро (ранее известный как рецептор ГАМКC) представляет собой подкласс ГАМКА-рецепторов, состоящий полностью из субъединиц ро (ρ). Рецептор ГАМКA-ро, как и другие рецепторы ГАМКA, экспрессируется во многих областях мозга, но, в отличие от других рецепторов ГАМКА, данный рецептор обладает особенно высокой экспрессией в сетчатке глаза[27].
Серотониновые рецепторы
Среди большого числа суперсемейств серотониновых рецепторов только одно принадлежит к суперсемейству цис-петлевых лиганд-зависимых ионных каналов — 5-HT3 и поэтому структурно и функционально отличается от всех других рецепторов 5-HT-рецепторов (5-гидрокситриптамина или серотонина), которые являются рецепторами, сопряжённые с G-белком (GPCR)[28][29][30]. 5-HT3 представляет собой селективный катионный канал, он обеспечивает деполяризацию и возбуждение нейронов в центральной и периферической нервной системе[28]. Как и в случае других лиганд-управляемых ионных каналов, рецептор 5-HT3 состоит из пяти субъединиц, расположенных вокруг центральной ионопроводящей поры, которая проницаема для ионов натрия (Na+), калия (K+) и кальция (Ca2+). Связывание нейромедиатора 5-гидрокситриптамина (серотонина) с рецептором 5-HT3 открывает канал, что, в свою очередь, приводит к возбуждающему ответу в нейронах (потенциалу действия). 5-HT3-рецепторы имеют незначительную проницаемость для анионов[28]. По структуре они наиболее гомологичны с никотиновыми рецепторами ацетилхолина.
Проявляемые эффекты
При активации рецептора агонистами происходит открытие катионного канала, что приводит к следующим эффектам:
АТФ-зависимые ионные каналы
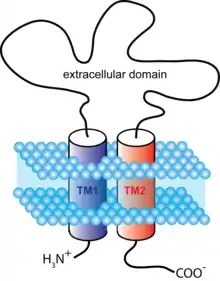
АТФ-зависимые ионные каналы открываются в ответ на связывание молекул нуклеотида АТФ[35]. Они образованы из тримеров с двумя трансмембранными спиралями на субъединицу и обоих терминалей (С и N-концов), расположенных на внутриклеточной стороне. К такому типу ионотропных рецепторов относится семейство P2X-пуринорецепторов. P2X-рецепторы присутствуют в разнообразных организмах, включая человека, позвоночных (млекопитающие, птицы, земноводные, рыбы итд.), беспозвоночных (трематоды) и простейших (амёбы)[36].
| Тип | Класс | Название белков согласно рекомендации ИЮФАР[12] | Ген | Предыдущее название |
|---|---|---|---|---|
| P2X-рецептор | N/A | P2X1 P2X2 P2X3 P2X4 P2X5 P2X6 P2X7 |
P2RX1 P2RX2 P2RX3 P2RX4 P2RX5 P2RX6 P2RX7 |
P2X1 P2X2 P2X3 P2X4 P2X5 P2X6 P2X7 |
PIP2-лиганд зависимые ионные каналы
Фосфатидилинозитол-4,5-бисфосфат (PIP2) связывается и непосредственно активирует калиевые каналы внутреннего выпрямления (Kir или IRK)[37]. PIP2 является липидом клеточной мембраны, и его роль в качестве лиганда ионных каналов представляет новую роль для данной молекулы[38][39]. Калиевые каналы внутреннего выпрямления обнаружены также у растений[40] и бактерий[41].
Клиническое значение
Лиганд-управляемые ионные каналы, вероятно, являются основным местом действия анестетиков и этанола, хотя однозначные доказательства, данного воздействия ещё предстоит установить[42][43]. В частности, анестетики воздействуют на ГАМК- и NMDA-рецепторы в концентрациях, аналогичных тем, которые используются в клинической анестезии[44].
Мемантин одобрен USFDA и Европейским агентством лекарственных средств для лечения болезни Альцгеймера средней и тяжёлой степени[45], и в настоящее время получил ограниченную рекомендацию Национального института здравоохранения и медицинского обслуживания Великобритании для пациентов, не прошедших другие варианты лечения[46].
- Лечение антидепрессантами
Агомелатин, является типом лекарственного средства, действующего по двойному мелатонергическому-серотонинергическому пути, он показал свою эффективность в лечении тревожной депрессии во время клинических испытаний[47][48], исследование также предполагает эффективность в лечении атипичных и меланхоличных типов депрессии[49].
Примечания
- Gene Family: Ligand gated ion channels. HUGO Gene Nomenclature Committee.
- "ligand-gated channel" at Dorland's Medical Dictionary
- Purves, Dale, George J. Augustine, David Fitzpatrick, William C. Hall, Anthony-Samuel LaMantia, James O. McNamara, and Leonard E. White. Neuroscience. 4th ed (неопр.). — Sinauer Associates, 2008. — С. 156—157. — ISBN 978-0-87893-697-7.
- Tasneem A., Iyer L., Jakobsson E., Aravind L. Identification of the prokaryotic ligand-gated ion channels and their implications for the mechanisms and origins of animal Cys-loop ion channels (англ.) // BioMed Central : journal. — 2004. — Vol. 6, no. 1. — P. R4. — doi:10.1186/gb-2004-6-1-r4. — PMID 15642096.
- Jaiteh M., Taly A., Hénin J. Evolution of Pentameric Ligand-Gated Ion Channels: Pro-Loop Receptors (англ.) // PLOS ONE : journal. — 2016. — Vol. 11, no. 3. — P. e0151934. — doi:10.1371/journal.pone.0151934. — PMID 26986966.
- Langlhofer, Georg; Villmann, Carmen. The Intracellular Loop of the Glycine Receptor: It's not all about the Size (англ.) // Frontiers in Molecular Neuroscience : journal. — 2016. — 1 January (vol. 9). — P. 41. — ISSN 1662-5099. — doi:10.3389/fnmol.2016.00041. — PMID 27330534.
- J., Aidley, David. The physiology of excitable cells (неопр.). — 4th. — Cambridge, UK: Cambridge University Press, 1998. — ISBN 978-0521574150.
- Lynch J. W. Molecular structure and function of the glycine receptor chloride channel (англ.) // Physiological Reviews : journal. — 2004. — October (vol. 84, no. 4). — P. 1051—1095. — doi:10.1152/physrev.00042.2003. — PMID 15383648.
- Rajendra, Sundran; Lynch, Joseph W.; Schofield, Peter R. The glycine receptor (англ.) // Pharmacology & Therapeutics : journal. — 1997. — Vol. 73, no. 2. — P. 121—146. — doi:10.1016/S0163-7258(96)00163-5.
- Duan L., Yang J., Slaughter M. M. Caffeine inhibition of ionotropic glycine receptors (англ.) // The Journal of Physiology : journal. — 2009. — August (vol. 587, no. Pt 16). — P. 4063—4075. — doi:10.1113/jphysiol.2009.174797. — PMID 19564396.
- Miyazawa A., Fujiyoshi Y., Unwin N. Structure and gating mechanism of the acetylcholine receptor pore (англ.) // Nature : journal. — 2003. — June (vol. 423, no. 6943). — P. 949—955. — doi:10.1038/nature01748. — PMID 12827192.
- Collingridge G. L., Olsen R. W., Peters J., Spedding M. A nomenclature for ligand-gated ion channels (неопр.) // Neuropharmacology. — 2009. — January (т. 56, № 1). — С. 2—5. — doi:10.1016/j.neuropharm.2008.06.063. — PMID 18655795.
- Olsen R. W., Sieghart W. International Union of Pharmacology. LXX. Subtypes of gamma-aminobutyric acid(A) receptors: classification on the basis of subunit composition, pharmacology, and function. Update (англ.) // Pharmacological Reviews : journal. — 2008. — September (vol. 60, no. 3). — P. 243—260. — doi:10.1124/pr.108.00505. — PMID 18790874.
- Traynelis S. F., Wollmuth L. P., McBain C. J., Menniti F. S., Vance K. M., Ogden K. K., Hansen K. B., Yuan H., Myers S. J., Dingledine R. Glutamate receptor ion channels: structure, regulation, and function (англ.) // Pharmacol. Rev. : journal. — 2010. — September (vol. 62, no. 3). — P. 405—496. — doi:10.1124/pr.109.002451. — PMID 20716669.
- Sobolevsky AI; Rosconi M. P., and Gouaux E. X-ray structure, symmetry and mechanism of an AMPA-subtype glutamate receptor (англ.) // Nature : journal. — 2009. — Vol. 462. — P. 745—756. — doi:10.1038/nature08624.
- Malenka R. C., Nestler E. J., Hyman S. E. Chapter 5: Excitatory and Inhibitory Amino Acids // Molecular Neuropharmacology: A Foundation for Clinical Neuroscience (англ.) / Sydor A., Brown R. Y.. — 2nd. — New York, USA: McGraw-Hill Medical, 2009. — P. 124—125. — ISBN 9780071481274.. — «At membrane potentials more negative than approximately −50 mV, the Mg2+ in the extracellular fluid of the brain virtually abolishes ion flux through NMDA receptor channels, even in the presence of glutamate. ... The NMDA receptor is unique among all neurotransmitter receptors in that its activation requires the simultaneous binding of two different agonists. In addition to the binding of glutamate at the conventional agonist-binding site, the binding of glycine appears to be required for receptor activation. Because neither of these agonists alone can open this ion channel, glutamate and glycine are referred to as coagonists of the NMDA receptor. The physiologic significance of the glycine binding site is unclear because the normal extracellular concentration of glycine is believed to be saturating. However, recent evidence suggests that D-serine may be the endogenous agonist for this site.».
- Li F., Tsien J. Z. Memory and the NMDA receptors (англ.) // The New England Journal of Medicine : journal. — 2009. — July (vol. 361, no. 3). — P. 302—303. — doi:10.1056/NEJMcibr0902052. — PMID 19605837.
- Cao X., Cui Z., Feng R., Tang Y. P., Qin Z., Mei B., Tsien J. Z. Maintenance of superior learning and memory function in NR2B transgenic mice during ageing (англ.) // The European Journal of Neuroscience : journal. — 2007. — March (vol. 25, no. 6). — P. 1815—1822. — doi:10.1111/j.1460-9568.2007.05431.x. — PMID 17432968.
- Dingledine R., Borges K., Bowie D., Traynelis S. F. The glutamate receptor ion channels (англ.) // Pharmacological Reviews : journal. — 1999. — March (vol. 51, no. 1). — P. 7—61. — PMID 10049997.
- Yarotskyy V., Glushakov A. V., Sumners C., Gravenstein N., Dennis D. M., Seubert C. N., Martynyuk A. E. Differential modulation of glutamatergic transmission by 3,5-dibromo-L-phenylalanine (англ.) // Molecular Pharmacology : journal. — 2005. — May (vol. 67, no. 5). — P. 1648—1654. — doi:10.1124/mol.104.005983. — PMID 15687225.
- Dingledine R., Borges K., Bowie D., Traynelis S. F. The glutamate receptor ion channels (англ.) // Pharmacol. Rev. : journal. — 1999. — Vol. 51, no. 1. — P. 7—61. — PMID 10049997. Архивировано 13 февраля 2009 года.
- Huettner J. E. Kainate receptors and synaptic transmission (англ.) // Prog. Neurobiol. : journal. — 2003. — Vol. 70, no. 5. — P. 387—407. — doi:10.1016/S0301-0082(03)00122-9. — PMID 14511698.
- Barnard E. A., Skolnick P., Olsen R. W., Mohler H., Sieghart W., Biggio G., Braestrup C., Bateson A. N., Langer S. Z. International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function (англ.) // Pharmacol. Rev. : journal. — 1998. — June (vol. 50, no. 2). — P. 291—313. — PMID 9647870.
- Hevers W., Lüddens H. The diversity of GABAA receptors. Pharmapoo and electrophysiological properties of GABAA channel subtypes (англ.) // Mol. Neurobiol. : journal. — 1998. — August (vol. 18, no. 1). — P. 35—86. — doi:10.1007/BF02741459. — PMID 9824848.
- Sieghart W., Sperk G. Subunit composition, distribution and function of GABA(A) receptor subtypes (англ.) // Curr Top Med Chem : journal. — 2002. — August (vol. 2, no. 8). — P. 795—816. — doi:10.2174/1568026023393507. — PMID 12171572.
- Wu C., Sun D. GABA receptors in brain development, function, and injury (англ.) // Metabolic Brain Disease : journal. — 2015. — April (vol. 30, no. 2). — P. 367—379. — doi:10.1007/s11011-014-9560-1. — PMID 24820774.
- Qian H. 2000. GABAc receptors in the vertebrate retina Архивная копия от 31 декабря 2010 на Wayback Machine. Retrieved on February 14, 2007.
- Barnes N. M., Hales T. G., Lummis S. C., Peters J. A. The 5-HT3 receptor--the relationship between structure and function (англ.) // Neuropharmacology : journal. — 2009. — January (vol. 56, no. 1). — P. 273—284. — doi:10.1016/j.neuropharm.2008.08.003. — PMID 18761359.
- Thompson A. J., Lummis S. C. 5-HT3 Receptors (англ.) // Current Pharmaceutical Design : journal. — 2006. — Vol. 12, no. 28. — P. 3615—3630. — doi:10.2174/138161206778522029. — PMID 17073663.
- Reeves D. C., Lummis S. C. The molecular basis of the structure and function of the 5-HT3 receptor: a model ligand-gated ion channel (review) (англ.) // Molecular Membrane Biology : journal. — 2002. — Vol. 19, no. 1. — P. 11—26. — doi:10.1080/09687680110110048. — PMID 11989819.
- Rang, H. P. Pharmacology (неопр.). — Edinburgh: Churchill Livingstone, 2003. — ISBN 0-443-07145-4., page 187.
- Gholipour T., Ghasemi M., Riazi K., Ghaffarpour M., Dehpour A. R. Seizure susceptibility alteration through 5-HT(3) receptor: modulation by nitric oxide (англ.) // Seizure : journal. — 2010. — January (vol. 19, no. 1). — P. 17—22. — doi:10.1016/j.seizure.2009.10.006. — PMID 19942458.
- Patel, Ryan; Dickenson, Anthony H. Modality selective roles of pro-nociceptive spinal 5-HT2A and 5-HT3 receptors in normal and neuropathic states (англ.) // Neuropharmacology : journal. — 2018. — September (vol. 143). — P. 29—37. — ISSN 0028-3908. — doi:10.1016/j.neuropharm.2018.09.028. — PMID 30240783.
- Suzuki, Rie; Rahman, Wahida; Rygh, Lars J; Webber, Mark; Hunt, Stephen P; Dickenson, Anthony H. Spinal-supraspinal serotonergic circuits regulating neuropathic pain and its treatment with gabapentin (англ.) // Pain : journal. — 2005. — October (vol. 117, no. 3). — P. 292—303. — ISSN 0304-3959. — doi:10.1016/j.pain.2005.06.015. — PMID 16150546.
- Сергей Козловский Двойная жизнь АТФ: и «батарейка», и нейромедиатор // Наука и жизнь, 2021, № 10. — с. 20-30
- North R. A. Molecular physiology of P2X receptors (англ.) // Physiological Reviews : journal. — 2002. — Vol. 82, no. 4. — P. 1013—1067. — doi:10.1152/physrev.00015.2002. — PMID 12270951.
- Hansen S. B., Tao X., MacKinnon R. Structural basis of PIP2 activation of the classical inward rectifier K+ channel Kir2.2 (англ.) // Nature : journal. — 2011. — August (vol. 477, no. 7365). — P. 495—498. — doi:10.1038/nature10370. — . — PMID 21874019.
- Hansen S. B. Lipid agonism: The PIP2 paradigm of ligand-gated ion channels (англ.) // Biochimica et Biophysica Acta : journal. — 2015. — May (vol. 1851, no. 5). — P. 620—628. — doi:10.1016/j.bbalip.2015.01.011. — PMID 25633344.
- Gao Y., Cao E., Julius D., Cheng Y. TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action (англ.) // Nature : journal. — 2016. — June (vol. 534, no. 7607). — P. 347—351. — doi:10.1038/nature17964. — . — PMID 27281200.
- Hedrich R. et al. Inward rectifier potassium channels in plants differ from their animal counterparts in response to voltage and channel modulators (англ.) // European Biophysics Journal. — 1995-10-01. — Vol. 24, no. 2. — P. 107—115. — ISSN 0175-7571. — doi:10.1007/BF00211406. — PMID 8582318.
- Choi S. B. et al. Identification and characterization of a novel bacterial ATP-sensitive K+ channel (англ.) // Journal of Microbiology (Seoul, Korea). — 2010. — 1 June (vol. 48, iss. 3). — P. 325—330. — ISSN 1976-3794. — doi:10.1007/s12275-010-9231-9.
- Krasowski M. D., Harrison N. L. General anaesthetic actions on ligand-gated ion channels (англ.) // Cellular and Molecular Life Sciences : journal. — 1999. — August (vol. 55, no. 10). — P. 1278—1303. — doi:10.1007/s000180050371. — PMID 10487207.
- Dilger J. P. The effects of general anaesthetics on ligand-gated ion channels (англ.) // British Journal of Anaesthesia : journal. — 2002. — July (vol. 89, no. 1). — P. 41—51. — doi:10.1093/bja/aef161. — PMID 12173240.
- Harris R. A., Mihic S. J., Dildy-Mayfield J. E., Machu T. K. Actions of anesthetics on ligand-gated ion channels: role of receptor subunit composition (англ.) // The FASEB Journal : journal. — Federation of American Societies for Experimental Biology, 1995. — November (vol. 9, no. 14). — P. 1454—1462. — PMID 7589987.
- Mount C., Downton C. Alzheimer disease: progress or profit? (англ.) // Nature Medicine : journal. — 2006. — July (vol. 12, no. 7). — P. 780—784. — doi:10.1038/nm0706-780. — PMID 16829947.
- NICE technology appraisal January 18, 2011 Azheimer's disease - donepezil, galantamine, rivastigmine and memantine (review): final appraisal determination
- Heun, R; Coral, RM; Ahokas, A; Nicolini, H; Teixeira, JM; Dehelean, P. 1643 – Efficacy of agomelatine in more anxious elderly depressed patients. A randomized, double-blind study vs placebo (англ.) // European Psychiatry : journal. — 2013. — Vol. 28, no. Suppl 1. — P. 1. — doi:10.1016/S0924-9338(13)76634-3.
- Brunton, L; Chabner, B; Knollman, B (2010). Goodman and Gilman's The Pharmacological Basis of Therapeutics (12th ed.). New York: McGraw-Hill Professional. ISBN 978-0-07-162442-8.
- Avedisova, A; Marachev, M. 2639 – The effectiveness of agomelatine (valdoxan) in the treatment of atypical depression (англ.) // European Psychiatry : journal. — 2013. — Vol. 28, no. Suppl 1. — P. 1. — doi:10.1016/S0924-9338(13)77272-9.
Внешние ссылки
- Ligand-Gated Ion Channel database at European Bioinformatics Institute. Verified availability April 11, 2007.
- Revised Recommendations for Nomenclature of Ligand-Gated Ion Channels. IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.
- www.esf.edu
- www.genenames.org
Шаблон:CCBYSASource