AMPA-рецептор
АМРА-рецептор (рецептор α-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты, AMPAR) — ионотропный рецептор глутамата, который передаёт быстрые возбуждающие сигналы в синапсах нервной системы позвоночных. Данные рецепторы также активируются синтетическим аналогом глутамата — аминокислотой АМРА, откуда и получили своё название. АМРА-рецепторы обнаружены практически во всех структурах головного мозга, их считают наиболее распространённым типом рецепторов в нервной системе. Эти рецепторы представляют собой тетрамерные ионные каналы, которые могут состоять из субъединиц четырёх типов[1]. АМРА-рецепторы имеют отношение к развитию некоторых заболеваний центральной нервной системы человека, таких как синдром Мартина — Белл, поэтому их изучению уделяется большое внимание[2].
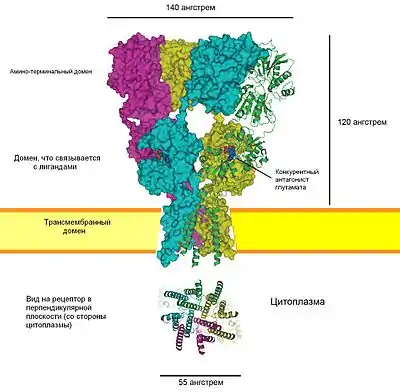
История
Рецептор был открыт группой учёных из фармакологического отделения университета Копенгагена под руководством Таге Хонор[3]. Гомотетрамерный АМРА-рецептор, состоящий из четырёх GluR2-субъединиц, стал первым из глутаматных рецепторов, который получили в виде кристаллов[4].
Распространение
АМРА-рецепторы — многочисленный и широко распространённый тип рецепторов в центральной нервной системе. Высокая концентрация субъединиц GluR1, GluR2 и GluR3 обнаружена в гиппокампе, наружных слоях коры переднего мозга, базальных ганглиях, обонятельных долях, миндалевидном теле и других зонах мозга. Субъединица GluR4 во многих участках мозга содержится в низкой концентрации, но в мозжечке, таламусе и спинном мозге её концентрация велика[5].
Методом иммунопреципитации было установлено, что в пирамидальных клетках гиппокампа экспрессируются АМРА-рецепторы, которые состоят из субъединицы GluR2 в сочетании с GluR1 или GluR3. В некоторых небольших популяциях нейронов встречаются гомомерные (то есть состоящие только из одного типа субъединиц) рецепторы GluR1. Такие рецепторы значительно отличаются по ионной проницаемости от других АМРА-рецепторов[6].
Экспрессия генов АМРА-рецепторов изменяется в онтогенезе. Субъединица GluR2 появляется начиная с 16-х суток эмбрионального развития мозга крысы, в то время как другие субъединицы появляются значительно позже[5]. Также относительное количество субъединиц GluR2 может изменяться вследствие синаптической пластичности, механических повреждений нервной ткани и других факторов.
АМРА-рецепторы были обнаружены и на постсинаптической, и на пресинаптической мембране химического синапса, и в меньшем количестве на внесинаптических участках плазматической мембраны нейронов. Около 60-70 % общего количества АМРА-рецепторов в клетке постоянно находятся внутри эндоплазматического ретикулума[7]. АМРА-рецепторы также присутствуют в клетках нейроглии, они участвуют в процессе апоптоза, вызванного глутаматной токсичностью[8]. Активация АМРА-рецепторов в клетках глии может приводить к Ca2+-зависимой активации NO-синтазы и последующему синтезу циклического ГМФ[9].
| Варианты названия субъединицы | Локализация гена в хромосомах человека |
Количество аминокислот в длинном сплайс-варианте |
|---|---|---|
| GluR1, GluRA, GRIA1, GluA1 | 5 q31.1 | 906 |
| GluR2, GluRB, GRIA2, GluA2 | 4 q32-q33 | 901 |
| GluR3, GluRC, GRIA3, GluA3 | X q25-q26 | 894 |
| GluR4, GluRD, GRIA4, GluA4 | 11 q22 | 902 |
Структура и функции субъединиц
Как было отмечено выше, структурно AMPA-рецепторы — тетрамеры, в которые могут входить субъединицы четырёх типов (GluR1—GluR4) в разных сочетаниях. Большинство АМРА-рецепторов является гетеротетрамерами, составленными «димером димеров»: одна субъединица в каждом из двух димеров обычно GluR2, а другая — GluR1, GluR3 или GluR4[11][12][13][14]. AMPA-рецепторы, в состав которых входят GluR2-субъединицы, содержащие аргинин в Q/R-сайте (см. ниже), являются непроницаемыми для ионов кальция, остальные же проницаемы для этих ионов[15].
Субъединицы АМРА-рецепторов состоят из четырёх доменов (структурно-функциональных участков): внеклеточного N-концевого домена (англ. amino-terminal domain, ATD); внеклеточного домена, связывающего лиганды (англ. ligand-binding domain, LBD); трансмембранного домена (англ. transmembrane domain, TMD) и внутриклеточного С-концевого домена (англ. carboxyl-terminal domain, CTD) (см. Рисунок 1). Тетрамеризация субъединиц происходит благодаря взаимодействию между лиганд-связывающими, трансмембранными и N-концевыми доменами соответствующих субъединиц[16][17]. Сборка рецепторов происходит в шероховатом эндоплазматическом ретикулуме[18], где особые механизмы обеспечивают правильное сворачивание субъединиц и их взаимное расположение. Показано, что внутри эндоплазматического ретикулума происходят изменения конформации рецепторов, связанные с их функциональной активностью: связыванием лиганда (глутамата), активацией, десенситизацией и другие; эти конформационные изменения способны влиять на процесс транспортировки рецепторов на наружную клеточную мембрану[18][19]. Кроме того, значительную роль в олигомеризации рецепторов и их транспорте играют N-концевые домены их субъединиц[20][21]. После окончательного формирования АМРА-рецепторы высвобождаются в цитоплазму.
Домен связывания лиганда
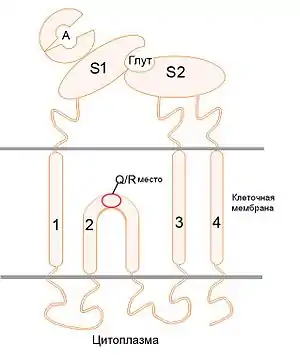
Домен связывания лиганда AMPA-рецептора формируется двумя внеклеточными сегментами, которые в силу исторических причин называются S1 и S2 (см. Рисунок 2)[22]. Эти два сегмента формируют структуру, напоминающую клешню, при этом сегмент S1, расположенный на N-конце мембранного сегмента М1 (см. ниже), формирует одну её половину, а сегмент S2, расположенный между сегментами М3 и М4, формирует другую (см. Рисунок 2). Участок связывания агониста помещается внутри «клешни» между двумя сегментами. Контакты между поверхностями сегментов S1, принадлежащих к разным субъединицам димера, создают несколько дополнительных мест связывания молекул аллостерических модуляторов[4].
Активация рецептора начинается со связывания агониста с доменом связывания лиганда. Глутамат, АМРА и их аналоги содержат структуры, соответствующие α-амино- и α-карбоксильным группам; эти группы связывают с определённые аминокислотные остатки в составе рецептора (см. Рисунок 1). Далее в процессе активации АМРА-рецептора благодаря связыванию молекулы лиганда происходит изменение конформации лиганд-связывающего домена. После связывания с агонистом сегменты S1 и S2 смыкаются гораздо теснее, чем когда рецептор находится в свободном состоянии. Сегмент S2 сдвигается и вызывает конформационную перестройку коротких цепочек аминокислотных остатков, которые объединяют домен связывания лиганда и трансмембранный домен; сегменты М3 в трансмембранных доменах субъединиц, в свою очередь, расходятся, открывая ионный канал в клеточной мембране (см. Рисунок 2)[23]. Движение сегментов S1 и S2 друг относительно друга приводит к нестабильному состоянию лиганд-связывающего и трансмембранного доменов. Стабильность макромолекулы может быть восстановлена в случае обратного открытия «клешни» в домене связывания лиганда, что происходит при закрытии ионного канала, и приводит к диссоциации комплекса лиганд-рецептор. Другой путь восстановления стабильности в макромолекуле заключается в изменении конформации контактной поверхности между субъединицами, которые формируют димер. В этом случае стабильность макромолекулы восстанавливается, лиганд остаётся связанным с ней, но ионный канал закрывается. Такое состояние рецептора называют «десенситизованным»: находясь в нём, рецептор неактивен (потому что ионный канал закрыт), но не может быть активирован, так как участок связывания агониста уже занят[24].
Альтернативный сплайсинг пре-мРНК субъединиц может приводить к образованию двух изоформ рецептора, называемых флип- и флоп-формами. Эти формы имеют разную чувствительность к аллостерическим модуляторам, а также у них по-разному происходят конформационные изменения в ходе активации, инактивации и десенситизации рецептора[25][26].
N-концевой домен
Первые 400—450 N-концевых аминокислотных остатков каждой субъединицы АМРА-рецептора (как и во всех других ионотропных глутаматных рецепторах) формируют N-концевой домен. По аминокислотной последовательности N-концевой домен ионотропных глутаматных рецепторов очень похож на лиганд-связывающий домен метаботропных глутаматных рецепторов и некоторые белки периплазмы бактерий. Было предположено, что N-концевой домен на ранних стадиях эволюции рецепторов был приспособлен для связывания эндогенных лигандов, но впоследствии потерял эту функцию[27][28][29][30][31]. С помощью методов генетической инженерии было создано большое количество мутантных субъединиц АМРА-рецептора, у которых N-концевой домен полностью отсутствует. Такие субъединицы способны формировать полностью функциональные рецепторы, однако, как было выяснено благодаря этим экспериментам, N-концевой домен имеет регуляторную функцию: его отсутствие влияет на вероятность открытия ионного канала рецептора, скорость инактивации, десенситизации и другие параметры.[20][21][32][33][34][35][36]. Кроме того, в N-концевом домене обнаружены центры связывания регуляторных молекул таких, как фенилэтаноламин, ифенпродил, а также пентраксины[37][38].
Трансмембранный домен
Трансмембранный домен АМРА-рецепторов состоит из четырёх трансмембранных сегментов: М1, М2, М3 и М4. В начале исследований рецептора такая структура трансмембранного домена вызвала некоторое недоумение: если аминокислотная цепь проходит сквозь клеточную мембрану чётное число раз, то её С-конец и N-конец должны быть расположены с одной стороны мембраны; но в то же время молекулярно-биологическими методами было установлено, что С-концевой фрагмент рецепторной субъединицы находится внутри клетки, а N-концевой — снаружи. Противоречие исчезло, когда выяснилось, что сегмент М2 не проходит мембрану насквозь, а изгибается и выходит на внутриклеточной стороне (см. Рисунок 2)[39].
Способность АМРА-рецепторов, содержащих GluR2-субъединицу, пропускать ионы зависит от посттранскрипционной модификации мРНК этой субъединицы: кодон, соответствующий в мРНК глутамину (Q), расположенному в белке на вершине перегиба сегмента М2 (Q/R-сайт), может быть заменён на кодон аргинина (R)[40]. Эта модификация существенно влияет на ионный транспорт через канал рецептора: Q-форма АМРА-рецепторов пропускает ионы Са2+ и может быть заблокирована полиаминными блокаторам ионного канала; в свою очередь, R-форма практически непроницаема для ионов кальция и почти нечувствительна к внутриклеточным полиаминным блокаторам[41]. Подавляющее большинство АМРА-рецепторов в нервной системе относится к R-форме.
При формировании рецепторного тетрамера сегменты М2 и М3 формируют собственно ионный канал. Сегмент М2 формирует его часть, выходящую на внутреннюю сторону клеточной мембраны; сегмент М3 — часть, выходящую на наружную сторону; сегмент М1, находясь кнаружи от ионного канала в плоскости мембраны, формирует внешний периметр трансмембранного домена рецептора; сегмент М4 формирует поверхность, комплементарную к поверхности сегментов М2 и М3 соседней субъединицы[4].
С-концевой домен
С-концевой домен АМРА-рецептора является наименее консервативным доменом: его первичная структура отличается у всех подтипов субъединиц. Этот домен содержит участок связывания многих внутриклеточных белков, которые регулируют движение рецепторов в клеточной мембране, их ионопроводность и другие характеристики[42]. Кроме того, С-концевые домены субъединиц различных типов могут взаимодействовать с различными клеточными сигнальными белками: например, С-концевой домен субъединицы GluR1 взаимодействует с гуанозинмонофосфат-зависимой протеинкиназой[43], С-концевой домен GluR4 — с протеинкиназой С[44]. Такое взаимодействие обеспечивает активацию или инактивацию, мембранный транспорт и другие функции рецепторов в ответ на внутриклеточные процессы.
| Субъединицы, составляющие рецептор |
Вероятность открытия при активации глутаматом |
Среднее время пребывания в открытом состоянии (мс) |
Электропроводность (pS) |
|---|---|---|---|
| GluR1-flip | 0,4-1,0[45][46] | 0,2-0,9[45] | 8, 15, 23, 31[45][47][48] |
| GluR2-flipQ | 0,61[49] | 0,32; 1,47[50] | 7, 15, 24, 36[50][51] |
| GluR3-flip | 0,82[52] | ||
| GluR4-flip | 0,77[46] | 0,14; 3,3[53] | 9, 20, 31, 45[53][54] |
Трансмембранные регуляторные белки
Исследования АМРА-рецепторов, экспрессированных в искусственных гетерогенных системах (ооциты лягушки, не-нейронные клеточные культуры) показали, что их характеристики отличаются от рецепторов, которые изучали в живой нервной ткани. Это несоответствие свидетельствует о существовании модулирующего компонента, присущего именно нервной ткани. Причины большого расхождения в характеристиках стали ясны после изучения трансмембранных белков, регулирующих активность АМРА-рецепторов (англ. transmembrane AMPA receptor regulatory Proteins, TARPs). TARP — это интегральные белки клеточной мембраны с четырьмя трансмембранными доменами, которые селективно взаимодействуют с АМРА-рецепторами, на ранних стадиях синтеза, во время транспортировки, встраивания в мембрану и передачи нервных сигналов[54][55][56]. С каждым тетрамером рецептора связаны два или четыре регуляторных белка, которые взаимодействуют с различными внутриклеточными белками[57][58]. Наиболее распространенные типы TARP (γ-2, γ-3, γ-4 и γ-8), взаимодействуют со всеми четырьмя типами субъединиц. TARP γ-2 (старгазин) был впервые обнаружен в мозжечке как белок, необходимый для транспорта АМРА-рецептора из эндоплазматического ретикулума к клеточной мембране[59]. В дополнение к транспортной функции, трансмембранные регуляторные белки, связываясь с АМРА-рецепторами, увеличивают проводимость ионного канала и вероятность его открытия, замедляют инактивацию и десенситизацию[54][60][61].
Фармакология
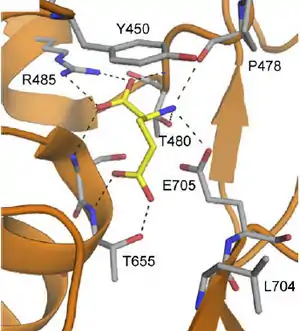
Основным эндогенным лигандом АМРА-рецепторов является глутамат, который связывается с «клешнеподобной» структурой в лиганд-связывающем домене каждой из субъединиц (см. выше), таким образом, рецептор имеет четыре участка связывания глутамата. Открытие ионного канала происходит после связывания агониста с двумя участками, но связывание с большим количеством участков увеличивает проводимость канала и среднее время его пребывания в открытом состоянии. Две карбоксильные и одна аминогруппа глутамата, образуют девять водородных связей с разными остатками аминокислот в лиганд-связывающем домене рецептора (см. Рисунок 3)[62].
Агонисты
Наряду с глутаматом, АМРА-рецептор может быть активирован другими природными и синтетическими лигандами: иботеновой кислотой, виллардиином, а также их многочисленными производными, а также производными АМРА (см. таблицу). Некоторые из этих агонистов селективны по отношению к субъединицам GluR1/GluR2 и GluR3/GluR4: например, Сl-НІВО (производное иботеновой кислоты) активирует GluR1 и GluR2 в 275 и в 1600 раз меньших концентрациях, чем GluR3 и GluR4 соответственно. Однако, несмотря на возможность фармакологического различия эффектов GluR1/GluR2 и GluR3/GluR4, к 2011 году не были открыты лиганды, которые позволяли бы различать эффекты индивидуальных субъединиц рецептора.
| Агонист | GluR1 | GluR2 | GluR3 | GluR4 |
|---|---|---|---|---|
| L-глутамат | 3,4-22[63][64][65][66] | 6.2-296[63][67][68] | 1.3-35[63][64][65] | 560[69] |
| АМРА | 1,3-8,7[65][70][71] | 66[68] | 1,4-130[65][70][71] | 1,3[71] |
| Каинат | 32-34[66][70] | 130-170[72] | 31-36[65][70] | |
| Виллардиин | 11,5[73] | 6.3[50] | ||
| F-Виллардиин | 0.47[73] | 0.2-0.5[50][74] | 20,9[74] | 11,9[74] |
| Br-Виллардиин | 2,8[73] | 0,84[50] | ||
| І-Виллардиин | 33,6[73] | 1,5[50] | ||
| Br-НІВО | 14[63] | 5,4[63] | 202[63] | 39[63] |
| Cl-НІВО | 4,7[75] | 1.7[75] | 2700[75] | 1300[75] |
| (S)-CPW399 | 24,9[76] | 13.9[76] | 224[76] | 34.3[76] |
| (S)-ATPA | 22[77] | 7.9[77] | 7.6[77] | |
| ACPA | 1,1-11[65][78] | 15[78] | 0,1-5[65][78] | 1,1[78] |
| (S)-4-AHCP | 4,5[79] | 7.2[79] | 15[79] | |
| (S)-Thio-ATPA | 5,2[80] | 13-40[80] | 32[80] | 20[80] |
| 2-Et-Tet-AMPA | 42[81] | 52[81] | 18[81] | 4[81] |
| (S)-2-Me-Tet-AMPA | 0,16[71] | 3,4[68] | 0,014[71] | 0,009[71] |
| SYM2081 | 132[64] | 453[64] | ||
| Домоевая кислота | 1,3[66] | 0,97[64] | 21[64] |
Конкурентные антагонисты
Конкурентные антагонисты АМРА-рецептора обычно содержат α-аминогруппу, соединённую с гетероциклическим участком[82]. Первыми изученными антагонистами рецептора были квиноксалиндионы (en:CNQX, DNQX, NBQX). Интересно, что в присутствии трансмембранных регуляторных белков, связанных с АМРА-рецепторами, CNQX и DNQX (но не NBQX) превращаются в слабые частичные агонисты. CNQX и DNQX вызывают частичное закрытие «клешни» домена связывания лиганда, что соответствует концепции действия частичного агониста[62]. По существующей гипотезе, трансмембраные регуляторные белки влияют на степень открытия «клешни» и делают её достаточной для индукции открытия ионного канала[83]. В отличие от квиноксалиндионов, соединения NS1209 и UВР282 стабилизируют комплекс S1-S2 в более «открытом» состоянии, чем характерно для несвязанного с лигандами рецептора.
| Антагонист | GluR1 | GluR2 | GluR3 | GluR4 |
|---|---|---|---|---|
| CNQX | 0,6[66] | 0,18[84] | 2,11[85] | |
| DNQX | 0,25[86] | 0,45[84] | 1,66[85] | 0,19-0,49[86] |
| NBQX | 0,4[87] | 0,59[78] | 0,31-0,63[78][85] | 0,1[87] |
| ATPO | 38[78] | 65[78] | 110[78] | 150[78] |
| YM90K | 1,96[85] | |||
| NS1209 | 0,12[88] | 0.13[88] | 0.11[88] | 0.06[88] |
| Кинуреновая кислота | 1900[89] | |||
| LY293558 | 9,2[90] | 0,4-3,2[90][91] | 32[92] | 51[90] |
| UBP310 | >100[93] | |||
| ACET | >100[93] |
Неконкурентные антагонисты
Основными классами неконкурентных антагонистов АМРА-рецептора являются 2,3-бензодиазепины (например, GYKI-53655), гидрофталазины и тетрагидроизокиналины[94]. В отличие от CNQX и DNQX, GYKI-53655 остается эффективным антагонистом АМРА-рецептора также и в присутствии трансмембранных регуляторных белков, к тому же его активность как антагониста даже повышается[95]. Доказано, что GYKI-53655 связывается одновременно с участками, которые объединяют сегменты S2 с М4 и S1 с М1[96]; последний участок является критическим звеном в открытии ионного канала[4].
| Aнтагонист | GluR1 | GluR2 | GluR3 | GluR4 |
|---|---|---|---|---|
| GYKI 52466 | 18-117[97][98] | 34[85] | 22-66[97][98] | |
| GYKI 53405 | 24[97] | 28[97] | ||
| GYKI 53655 | 6[97] | 5[97] | ||
| LY 300164 | 21[99] | 18[99] | 19[99] | 18[99] |
| CP-465,022 | 0,5[96] | 0,5[96] | 0,3[96] |
Бесконкурентные антагонисты
Бесконкурентные антагонисты АМРА-рецептора, такие как филантотоксины[100] или блокаторы каналов, для своего действия требуют предварительного перехода ионного канала рецептора к открытому состоянию, после связывания со специфическим участком внутри канала эти вещества механически блокируют прохождение ионов сквозь него[101]. Таким образом, эффект этих антагонистов (кривая доза-эффект) зависит от степени предварительной активации рецепторов в исследуемой ткани. В свою очередь, реактивация рецептора после их связывания происходит только под действием агониста, который способен вызвать открытие ионного канала, поэтому восстановление деятельности рецепторов после воздействия таких антагонистов происходит, как правило, медленнее, чем для антагонистов предыдущих классов.
| Антагонист | GluR1 | GluR2 | GluR3 | GluR4 |
|---|---|---|---|---|
| Аргиотоксин 636 | 0,35-3,4[102][103] | Н. Д.[102] | 0,23[102] | 0,43[102] |
| Токсин джоро | 0,04[104] | Н. Д.[104] | 0,03[104] | |
| Филантотоксин 433 | 0,8[105] | |||
| Филантотоксин 343 | 2,8[103] | |||
| Филантотоксин 56 | 3,3pM[106] | |||
| Филантотоксин 74 | 2,8[106] | |||
| IEM-1460 | 1,6[107] | Н. Д.[108] | 1,6[107] | |
| IEM-1754 | 6,0[107] | 6,0[107] |
Аллостерические модуляторы
Аллостерическими модуляторами называют вещества, которые изменяют активность рецептора путём изменения хода процессов инактивации и десенситизации[109]. Связывание агониста с доменом связывания лиганда приводит к возникновению «напряжения» в рецепторе, которые могут быть сняты двумя путями: открытием ионного канала (активация рецептора), либо изменением конформации молекулы на такую, где канал закрыт, но напряжение отсутствует (десенситизация рецептора). В первом случае после диссоциации лиганд-рецепторного комплекса ионный канал закрывается, а рецептор переходит к ненапряжённой конформации (отключение). Связывание положительных модуляторов АМРА-рецептора (например, пирацетама[110]) с лиганд-связывающим доменом приводит к повышению энергии перехода рецептора в ненапряжённое состояние после связывания с агонистом. Таким образом, модуляторы предотвращают десенситизацию рецептора. Некоторые из модуляторов также способны замедлять или ускорять диссоциацию комплекса агонист-рецептор, таким образом происходит модуляция процесса инактивации.
Важнейшим параметром, определяющим разницу между аллостерическими модуляторами, является именно механизм их действия. В частности, анирацетам замедляет процесс инактивации, но не влияет на силу действия агонистов; en:РЕРА усиливает действие АМРА-рецепторов, уменьшает десенситизацию, но не влияет на инактивацию; циклотиазид увеличивает сродство агонистов[111]. В свою очередь, соединение LY404187 стабилизирует АМРА-рецептор в открытом состоянии после связывания его с агонистом и не влияет на скорость его десенситизации. Кроме того, это соединение, вероятно, позволяет десенситизированным рецепторам переходить к открытому состоянию либо напрямую, либо через промежуточную десенситизованную и/или закрытую конформацию[112]. Некоторые соединения (например, СХ614) одновременно ингибируют и процесс десенситизации, и процесс инактивации по неизвестному механизму[113]. Сила аллостерических модуляторов может зависеть от сплайс-вариантов рецептора, с которыми они взаимодействуют. Например, циклотиазид практически полностью предотвращает десенситизацию флип-варианта рецептора, но является лишь умеренно активным в случае связывания с флоп-вариантом[52].
Активация и инактивация
Скорость активации и инактивации является одной из ключевых характеристик рецептора для физиологии синапсов, синаптической пластичности и в формировании нервных импульсов. Характеристики активации и инактивации отличаются в зависимости от субъединиц, составляющих рецептор, их сплайс-вариантов, наличия регулирующих белков и других факторов. По сравнению с другими типами ионотропных глутаматных рецепторов (NMDA-рецепторами, каинатными рецепторами) АМРА-рецепторы характеризуются быстрой активацией, инактивацией и десенситизацией. Это позволяет модулировать мембранные токи с большим временным разрешением, изменяя характеристики нервного сигнала в течение миллисекунд[114].
| Субъединицы, составляющие рецептор |
-инактивации | -десенситизации | -восстановления |
|---|---|---|---|
| GluR1-flip | 0,7-1,2[25][26][115][116] | 2,5-4,1[25][26][115][116][117] | 111-147[26][115][118] |
| GluR1-flop | 0,86-1,3[25][26][115][116][119] | 3,2-4,2[25][26][115][116][117][119] | 147-155[26][115][119] |
| GluR2-flipQ | 0,62-1,1[49][116] | 5,9-9,9[49][116][117] | 11,7[49] |
| GluR2-flopQ | 0,54-0,9[49][116] | 1,2-1,9[49][116][117] | 31,3[49] |
| GluR3-flip | 0,56[52] | 3,0-5,1[25][52][117][120] | 15-70[52][121] |
| GluR3-flop | 0,63-1,05[52][119] | 1,1-2,8[25][52][116][117][119][120] | 55-142[52][108][120] |
| GluR4-flip | 0,6[25] | 3,6-5,1[25][117] | 6-21[118][121] |
| GluR4-flop | 0,6[25] | 0,9[25][117] | 31-43[121] |
| GluR1-flip/GluR2-flip | 5,1[25] | 28-67[25] | |
| GluR3-flip/GluR2-flip | 4,9[25] | 15-26[25] |

Роль АМРА-рецепторов в синаптической пластичности
Синаптическая пластичность — это явление изменения силы синапса в ответ на его собственную активность или другие сигналы. Долговременная синаптическая потенциация и длительное синаптическое подавление являются широко известными формами синаптической пластичности, которые традиционно связывают с механизмами памяти и забывания. АМРА-рецепторы играют важную роль в двух этих процессах.
Роль в долговременной синаптической потенциации
Долговременная синаптическая потенциация — это усиление синаптической передачи между двумя нейронами, которое происходит после короткого периода активности синапса и может сохраняться в течение, по крайней мере, нескольких часов. Явление долговременной синаптической потенциации в глутаматных синапсах зависит от свойств как пресинаптической (высвобождения глутамата), так и постсинаптической (изменение количества рецепторов глутамата) мембран. Долговременную потенциацию считают одним из механизмов формирования и контроля памяти. Доказано, что АМРА-рецепторы играют важную роль в формировании эффекта долговременной потенциации и что их концентрация в синаптической области при этом увеличивается[122].
Ионный механизм
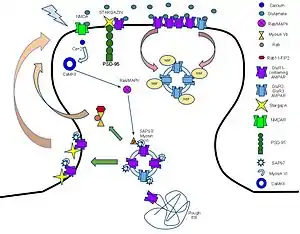
Роль АМРА-рецепторов в формировании быстрого компонента долговременной потенциации заключается в следующем. Глутамат, высвобождающийся из пресинаптического нейрона, связывается с несколькими рецепторами, содержащими ионные каналы, в частности с рецепторами к АМРА и к NMDA. Связывание с лигандом приводит к открытию каналов АМРА-рецепторов, которые пропускают ионы натрия внутрь клетки, что приводит к деполяризации клеточной мембраны. С другой стороны, NMDA-рецепторы в начале процесса долговременной потенциации не открываются, потому что их ионный канал при нормальных значениях мембранного потенциала блокирован ионами магния. Но, благодаря поступлению ионов натрия через АМРА-рецепторы, мембранный потенциал снижается настолько, что этого достаточно для высвобождения магния из NMDA-рецепторов и открытия их ионных каналов. В отличие от АМРА-рецепторов, NMDA-рецепторы пропускают не только натрий, но и ионы кальция. Кальций, поступающий в клетки, опосредует эффекты АМРА-рецепторов: в частности, он приводит к фосфорилированию фермента кальмодулин-зависимой протеинкиназы II (CaMKII), который вызывает фосфорилирование субъединиц АМРА-рецептора и повышает проводимость ионных каналов. Повышение ионной проводимости каналов АМРА-рецепторов приводит к активному поступлению натрия в клетку, таким образом осуществляется положительная обратная связь (Рисунок 4).
Кальмодулин-зависимая протеинкиназа II способна инициировать несколько различных путей транспортировки АМРА-рецепторов на внешнюю перисинаптическую мембрану. Во-первых, она напрямую фосфорилирует синапс-ассоциированный белок 97 (англ. synaptic-associated protein 97, SAP97)[123], который совместно с миозином-VI связывается с С-концевыми участками субъединиц АМРА-рецептора. После фосфорилирования кальмодулин-зависимой протеинкиназой II этот комплекс транспортируется в перисинаптическую мембрану[124]. Во-вторых, возможна активация транспорта по МАРК-зависимому пути. В этом случае кальмодулин-зависимая протеинкиназа II активирует белки Ras, которые, в свою очередь, активируют МАРК p42/44 (митоген-активируемые протеинкиназы), что приводит к транспорту и встраиванию АМРА-рецепторов в синаптическую мембрану[125].
Транспорт к постсинаптическому уплотнению
После попадания АМРА-рецептора на перисинаптические участки клеточной мембраны по CaMKII- или MAPK-зависимому пути, рецепторы движутся к постсинаптическому уплотнению. Одним из возможных механизмов этого процесса является непосредственный латеральный транспорт АМРА-рецепторов из перисинаптической мембраны к постсинаптическому уплотнению при долговременной потенциации[126]. Другой возможный механизм транспорта — это захват рецепторов на внесинаптических участках и их перенос к синапсу в везикулах внутри клетки[127]. В течение долговременной потенциации происходят оба описанных процесса, но только латеральный транспорт рецепторов в клеточной мембране непосредственно увеличивает их количество в постсинаптическом уплотнении. Везикулярный транспорт, в свою очередь, обеспечивает поступление новых порций АМРА-рецепторов в околосинаптическую область, откуда они могут далее транспортироваться в постсинаптическое уплотнение[128]. Обнаружено несколько белков, которые являются критически важными для транспорта рецепторов. Например, повышенный синтез белка SAP97 приводит к более активному, чем при обычных условиях, движению АМРА-рецепторов к синапсам[129]. Другие белки, активность которых влияет на мембранный транспорт АМРА-рецепторов, это миозин и кальций-зависимые моторные белки[130].
Роль в длительном синаптическом подавлении
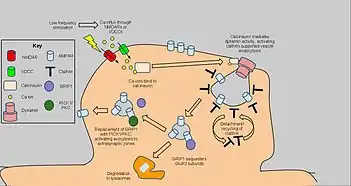
Длительное синаптическое подавление (англ. Long-term Depression, LTD) — уменьшение синаптической передачи между двумя нейронами после периода активности синапса. Этот процесс противоположен долговременной синаптической потенциации. Установление длительного синаптического подавления сопровождается уменьшением количества АМРА-рецепторов в постсинаптических участках дендритов по клатрин- и кальцинейрин-зависимым механизмам. Также при этом происходит транспорт рецепторов по другому механизму, чем при долговременной потенциации. Сигналом для начала эндоцитоза АМРА-рецепторов является поступление кальция из внеклеточной среды через NMDA-рецептор, ионы Са2+ активируют фосфатазы и кальцинейрин. Запуск эндоцитоза также зависит от потенциал-зависимых кальциевых каналов, вероятно, эндоцитоз АМРА-рецепторов индуцируется повышением внутриклеточной концентрации кальция независимо от конкретного механизма[7]. В то время как ингибирование фосфатаз почти не влияет на эндоцитоз рецепторов, добавление антагонистов кальцинейрина существенно его угнетает[131].
В постсинаптической зоне кальцинейрин контактирует с белковым комплексом, осуществляющим эндоцитоз. Этот комплекс представляет собой массив клатрина, расположенный под участком мембраны, содержащим АМРА-рецепторы, а также белки, которые осуществляют эндоцитоз рецепторов (особенно эффективно в случае, если они содержат субъединицу GluR2 и/или GluR3). Активация кальцинейрина вызывает активацию ГТФазы динамина, при этом происходит продвижение клатринового массива внутрь клетки и формируется внутриклечтоная везикула[132]. АМРА-рецепторы, перенесённые в цитоплазму, далее разрушаются в лизосомах или вновь переносятся в клеточную мембрану в перисинаптической зоне благодаря действию белков РІСК1 и РКС (см. Рисунок 5)[133][134].
Примечания
- Nakagawa T. The biochemistry, ultrastructure, and subunit assembly mechanism of AMPA receptors (англ.) // Mol Neurobiol. : journal. — 2010. — Vol. 42. — P. 161—184. — PMID 21080238.
- Bowie D. Ionotropic glutamate receptors & CNS disorders (неопр.) // CNS Neurol Disord Drug Targets. — 2008. — Т. 7. — С. 129—143. — doi:10.1007/s12035-010-8149-x. — PMID 18537642.
- Honore T., Lauridsen J., Krogsgaard-Larsen P. The binding of [3H]AMPA, a structural analogue of glutamic acid, to rat brain membranes (англ.) // Journal of Neurochemistry : journal. — 1982. — Vol. 38, no. 1. — P. 173—178. — doi:10.1111/j.1471-4159.1982.tb10868.x. — PMID 6125564.
- Sobolevsky AI; Rosconi M. P., and Gouaux E. X-ray structure, symmetry and mechanism of an AMPA-subtype glutamate receptor (англ.) // Nature : journal. — 2009. — Vol. 462. — P. 745—756. — doi:10.1038/nature08624.
- Allen Brain Atlas
- Dingledine R; Borges K., Bowie D., et al. The glutamate receptor ion channels (англ.) // Pharmacol Rev : journal. — 1999. — Vol. 51, no. 1. — P. 7—61.
- Carroll RC; Beattie E. C., Xia H., Luscher C., Altschuler Y., Nichol R. A., Malenka RC and Zastrow M. Dynamin-dependent endocytosis of ionotropic glutamate receptors (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1999. — Vol. 96, no. 24. — P. 14112—14117. — doi:10.1073/pnas.96.24.14112.
- Matute C; Domercq M., Sánchez-Gómez M. V. Glutamate-mediated glial injury: mechanisms and clinical importance (англ.) // Glia : journal. — 2006. — Vol. 53. — P. 212—224. — doi:10.1002/glia.20275.
- Baltrons MA; García A. AMPA receptors are coupled to the nitric oxide/cyclic GMP pathway in cerebellar astroglial cells (англ.) // Eur J Neurosci. : journal. — 1997. — Vol. 9, no. 11. — P. 2497—2501. — doi:10.1111/j.1460-9568.1997.tb01667.x.
- Collingridge GL; Olsen R. W., Peters J., and Spedding M. A nomenclature for ligand-gated ion channels (неопр.) // Neuropharmacology. — 2009. — Т. 56, № 1. — С. 2—5. — doi:10.1016/j.neuropharm.2008.06.063.
- Shi SH; Hayashi Y., Petralia R. S., et al. Rapid spine delivery and redistribution of AMPA receptors after synaptic NMDA receptor activation (англ.) // Science : journal. — 1999. — Vol. 284, no. 5421. — P. 1811—1816. — doi:10.1016/S0166-2236(02)02270-1.
- Mayer M. L. Glutamate receptor ion channels (неопр.) // Curr Opin Neurobiol. — 2005. — Т. 15, № 3. — С. 282—288. — doi:10.1016/j.conb.2005.05.004.
- Song I; Huganir R. L. Regulation of AMPA receptors during synaptic plasticity (англ.) // Trends Neurosci : journal. — 2002. — Vol. 25, no. 11. — P. 578—588. — doi:10.1016/S0166-2236(02)02270-1.
- Greger IH; Ziff E. B., Penn A. C. Molecular determinants of AMPA receptor subunit assembly (англ.) // Trends Neurosci : journal. — 2007. — Vol. 30, no. 8. — P. 407—416. — doi:10.1016/j.tins.2007.06.005.
- Liu S; Lau L., Wei J., Zhu D., Zou S., Sun H. S., Fu Y., Liu F., Lu Y. Expression of Ca(2+)-permeable AMPA receptor channels primes cell death in transient forebrain ischemia (англ.) // Neuron : journal. — Cell Press, 2004. — Vol. 43, no. 1. — P. 43—55. — doi:10.1016/j.neuron.2004.06.017.
- Mansour M; Nagarajan N., Nehring R. B., Clements J. D., and Rosenmund C. Heteromeric AMPA receptors assemble with a preferred subunit stoichiometry and spatial arrangement (англ.) // Neuron : journal. — Cell Press, 2001. — Vol. 32. — P. 841—853. — doi:10.1016/S0896-6273(01)00520-7.
- Kim KS; Yan D., Tomita S. Assembly and stoichiometry of the AMPA receptor and transmembrane AMPA receptor regulatory protein complex (англ.) // J Neurosci. : journal. — 2010. — Vol. 30, no. 3. — P. 1064—1072. — doi:10.1016/S0896-6273(01)00520-7.
- Greger IH; Khatri L., Ziff E. B. RNA editing at arg607 controls AMPA receptor exit from the endoplasmic reticulum (англ.) // Neuron : journal. — Cell Press, 2002. — Vol. 34, no. 5. — P. 759—772. — doi:10.1016/S0896-6273(02)00693-1.
- Penn AC; Williams S. R., and Greger I. H. Gating motions underlie AMPA receptor secretion from the endoplasmic reticulum (англ.) // EMBO J : journal. — 2008. — Vol. 27, no. 22. — P. 3056—3068. — doi:10.1038/emboj.2008.222.
- Ayalon G; Segev E., Elgavish S., and Stern-Bach Y. Two regions in the Nterminal domain of ionotropic glutamate receptor 3 form the subunit oligomerization interfaces that control subtype-specific receptor assembly (англ.) // J Biol Chem : journal. — 2005. — Vol. 280, no. 15. — P. 15053—15060. — doi:10.1074/jbc.M408413200.
- Kuusinen A; Abele R., Madden D. R., and Keinänen K. Oligomerization and ligand-binding properties of the ectodomain of the alpha-amino-3-hydroxy-5- methyl-4-isoxazole propionic acid receptor subunit GluRD (англ.) // J Biol Chem : journal. — 1999. — Vol. 274, no. 41. — P. 28937—28943. — doi:10.1074/jbc.274.41.28937.
- Stern-Bach Y; Bettler B., Hartley M., Sheppard P. O., O’Hara P. J., and Heinemann S. F. Agonist selectivity of glutamate receptors is specified by two domains structurally related to bacterial amino acid-binding proteins (англ.) // Neuron : journal. — Cell Press, 1994. — Vol. 13, no. 6. — P. 1345—1357. — doi:10.1016/0896-6273(94)90420-0.
- Hansen KB; Yuan H., and Traynelis S. F. Structural aspects of AMPA receptor activation, desensitization and deactivation (англ.) // Curr Opin Neurobiol : journal. — 2007. — Vol. 17, no. 3. — P. 281—288. — doi:10.1016/j.conb.2007.03.014.
- Weston МС; Schuck P., Ghosal A., Rosenmund C., and Mayer M. L. Conformational restriction blocks glutamate receptor desensitization (англ.) // Nat Struct Mol Biol : journal. — 2006. — Vol. 13. — P. 1120—1127. — doi:10.1038/nsmb1178.
- Mosbacher J; Schoepfer R., Monyer H., Burnashev N., Seeburg P. H., and Ruppersberg J. P. A molecular determinant for submillisecond desensitization in glutamate receptors (рум.) // Science. — 1994. — Т. 266, nr. 5187. — P. 1059—1062. — doi:10.1126/science.7973663.
- Partin KM; Fleck M. W., and Mayer M. L. AMPA receptor flip/flop mutants affecting deactivation, desensitization, and modulation by cyclothiazide, aniracetam, and thiocyanate (англ.) // J Neurosci : journal. — 1996. — Vol. 16, no. 21. — P. 6634—6647.
- Paoletti P; Perin-Dureau F., Fayyazuddin A., Le Goff A., Callebaut I., and Neyton J. Molecular organization of a zinc binding n-terminal modulatory domain in a NMDA receptor subunit (англ.) // Neuron. — Cell Press, 2000. — Vol. 28, no. 3. — P. 911—925. — doi:10.1016/S0896-6273(00)00163-X.
- Clayton A; Siebold C., Gilbert R. J., Sutton G. C., Harlos K., McIlhinney R. A., Jones E. Y., and Aricescu A. R. Crystal structure of the GluR2 amino-terminal domain provides insights into the architecture and assembly of ionotropic glutamate receptors (англ.) // J Mol Biol : journal. — 2009. — Vol. 392, no. 5. — P. 1125—1132. — doi:10.1016/j.jmb.2009.07.082.
- Jin R; Singh S. K., Gu S., Furukawa H., Sobolevsky A. I., Zhou J., Jin Y., and Gouaux E. Crystal structure and association behaviour of the GluR2 amino-terminal domain (англ.) // EMBO J : journal. — 2009. — Vol. 28, no. 12. — P. 1812—1823. — doi:10.1038/emboj.2009.140.
- Karakas E; Simorowski N., and Furukawa H. Structure of the zinc-bound amino-terminal domain of the NMDA receptor NR2B subunit (англ.) // EMBO J : journal. — 2009. — Vol. 28, no. 24. — P. 3910—3920. — doi:10.1038/emboj.2009.338.
- Kumar J; Schuck P., Jin R., and Mayer M. L. The N-terminal domain of GluR6-subtype glutamate receptor ion channels (англ.) // Nat Struct Mol Biol : journal. — 2009. — Vol. 16, no. 6. — P. 631—638. — doi:10.1038/nsmb.1613.
- Leuschner WD; and Hoch W. Subtype-specific assembly of alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor subunits is mediated by their N-terminal domains (англ.) // J Biol Chem : journal. — 1999. — Vol. 274, no. 24. — P. 16907—16916. — doi:10.1074/jbc.274.24.16907.
- Ayalon G; and Stern-Bach Y. Functional assembly of AMPA and kainate receptors is mediated by several discrete protein-protein interactions (англ.) // Neuron : journal. — Cell Press, 2001. — Vol. 31, no. 1. — P. 103—113. — doi:10.1016/S0896-6273(01)00333-6.
- Meddows E; Le Bourdelles B., Grimwood S., Wafford K., Sandhu S., Whiting P., and McIlhinney R. A. Identification of molecular determinants that are important in the assembly of N-methyl-D-aspartate receptors (англ.) // J Biol Chem : journal. — 2001. — Vol. 276, no. 22. — P. 18795—18803. — doi:10.1074/jbc.M101382200.
- Gielen M; Siegler Retchless B., Mony L., Johnson J. W., and Paoletti P. Mechanism of differential control of NMDA receptor activity by NR2 subunits (англ.) // Nature : journal. — 2009. — Vol. 459, no. 7247. — P. 703—707. — doi:10.1038/nature07993.
- Yuan H; Hansen K. B., Vance K. M., Ogden K. K., and Traynelis S. F. Control of NMDA receptor function by the NR2 subunit amino-terminal domain (англ.) // J Neurosci : journal. — 2009. — Vol. 29, no. 39. — P. 12045—12058. — doi:10.1523/JNEUROSCI.1365-09.2009.
- O’Brien RJ; Xu D., Petralia R. S., Steward O., Huganir R. L., and Worley P. Synaptic clustering of AMPA receptors by the extracellular immediate-early gene product Narp (англ.) // Neuron : journal. — Cell Press, 1999. — Vol. 23, no. 2. — P. 309—323. — doi:10.1016/S0896-6273(00)80782-5.
- Sia GM; Béïque J. C., Rumbaugh G., Cho R., Worley P. F., and Huganir R. L. Interaction of the N-terminal domain of the AMPA receptor GluR4 subunit with the neuronal pentraxin NP1 mediates GluR4 synaptic recruitment (англ.) // Neuron : journal. — Cell Press, 2007. — Vol. 55, no. 1. — P. 87—102. — doi:10.1016/j.neuron.2007.06.020.
- Hollmann M; Maron C., and Heinemann S. N-glycosylation site tagging suggests a three transmembrane domain topology for the glutamate receptor GluRI (англ.) // Neuron : journal. — Cell Press, 1994. — Vol. 13, no. 6. — P. 1331—1343. — doi:10.1016/0896-6273(94)90419-7.
- Bass B. L. RNA editing by adenosine deaminases that act on RNA (англ.) // Annu Rev Biochem : journal. — 2002. — Vol. 71. — P. 817—846. — doi:10.1146/annurev.biochem.71.110601.135501.
- Panchenko VA; Glasser C. R., Partin K. M., and Mayer M. L. Amino acid substitutions in the pore of rat glutamate receptors at sites influencing block by polyamines (англ.) // J Physiol : journal. — 1999. — Vol. 520, no. 2. — P. 337—357. — doi:10.1111/j.1469-7793.1999.t01-1-00337.x.
- Uchino S; Wada H., Honda S., Nakamura Y., Ondo Y., Uchiyama T., Tsutsumi M., Suzuki E., Hirasawa T., and Kohsaka S. Direct interaction of post-synaptic density-95/Dlg/ZO-1 domain-containing synaptic molecule Shank3 with GluR1 alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor (англ.) // J Neurochem : journal. — 2006. — Vol. 97, no. 4. — P. 1203—1214. — doi:10.1111/j.1471-4159.2006.03831.x.
- Serulle Y; Zhang S., Ninan I., Puzzo D., McCarthy M., Khatri L., Arancio O., and Ziff E. B. A GluR1-cGKII interaction regulates AMPA receptor trafficking (англ.) // Neuron : journal. — Cell Press, 2007. — Vol. 56, no. 4. — P. 670—688. — doi:10.1016/j.neuron.2007.09.016.
- Correia SS; Duarte C. B., Faro C. J., Pires E. V., and Carvalho A. L. Protein kinase C gamma associates directly with the GluR4 alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate receptor subunit. Effect on receptor phosphorylation (англ.) // J Biol Chem : journal. — 2003. — Vol. 278, no. 8. — P. 6307—6313. — doi:10.1074/jbc.M205587200.
- Banke TG; Bowie D., Lee H., Huganir R. L., Schousboe A., and Traynelis S. F. Control of GluR1 AMPA receptor function by cAMP-dependent protein kinase (англ.) // J Neurosci : journal. — 2000. — Vol. 20, no. 1. — P. 89—102.
- Robert A; Irizarry S. N., Hughes T. E., and Howe J. R. Subunit interactions and AMPA receptor desensitization (англ.) // J Neurosci : journal. — 2001. — Vol. 21, no. 15. — P. 5574—5586.
- Derkach V; Barria A., and Soderling T. R. Ca2+/calmodulin-kinase II enhances channel conductance of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate type glutamate receptors (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1999. — Vol. 96, no. 6. — P. 3269—3274. — doi:10.1073/pnas.96.6.3269.
- Prieto ML; and Wollmuth L. P. Gating modes in AMPA receptors (англ.) // J Neurosci : journal. — 2010. — Vol. 30, no. 12. — P. 4449—4459. — doi:10.1523/JNEUROSCI.5613-09.2010.
- Koike M; Tsukada S., Tsuzuki K., Kijima H., and Ozawa S. Regulation of kinetic properties of GluR2 AMPA receptor channels by alternative splicing (англ.) // J Neurosci : journal. — 2000. — Vol. 20, no. 6. — P. 2166—2174.
- Jin R; Banke T. G., Mayer M. L., Traynelis S. F., and Gouaux E. Structural basis for partial agonist action at ionotropic glutamate receptors (англ.) // Nat Neurosci : journal. — 2003. — Vol. 6. — P. 803—810. — doi:10.1038/nn1091.
- Zhang W; Cho Y., Lolis E., and Howe J. R. Structural and single-channel results indicate that the rates of ligand binding domain closing and opening directly impact AMPA receptor gating (англ.) // J Neurosci : journal. — 2008. — Vol. 28, no. 4. — P. 932—943.
- Sekiguchi M; Nishikawa K., Aoki S., and Wada K. A desensitization-selective potentiator of AMPA-type glutamate receptors (англ.) // Br J Pharmacol : journal. — 2002. — Vol. 136, no. 7. — P. 1033—1041. — doi:10.1038/sj.bjp.0704804.
- Swanson GT; Kamboj S. K., and Cull-Candy S. G. Single-channel properties of recombinant AMPA receptors depend on RNA editing, splice variation, and subunit composition (англ.) // J Neurosci : journal. — 1997. — Vol. 17, no. 1. — P. 58—69.
- Tomita S; Adesnik H., Sekiguchi M., Zhang W., Wada K., Howe J. R., Nicoll R. A., and Bredt D. S. Stargazin modulates AMPA receptor gating and trafficking by distinct domains (англ.) // Nature : journal. — 2005. — Vol. 435. — P. 1052—1058. — doi:10.1038/nature03624.
- Tomita S; Fukata M., Nicoll R. A., and Bredt D. S. Dynamic interaction of stargazin-like TARPs with cycling AMPA receptors at synapses (англ.) // Science : journal. — 2004. — Vol. 303, no. 5663. — P. 1508—1511. — doi:10.1126/science.1090262.
- Vandenberghe W; Nicoll R. A., and Bredt D. S. Stargazin is an AMPA receptor auxiliary subunit. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2005. — Vol. 102, no. 2. — P. 485—490. — doi:10.1073/pnas.0408269102.
- Milstein AD; and Nicoll. TARP modulation of synaptic AMPA receptor trafficking and gating depends on multiple intracellular domains (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2009. — Vol. 106, no. 27. — P. 11348—11351. — doi:10.1073/pnas.0905570106.
- Sager C; Terhag J., Kott S., and Hollmann M. C-terminal domains of transmembrane alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate (AMPA) receptor regulatory proteins not only facilitate trafficking but are major modulators of AMPA receptor function (англ.) // J Biol Chem : journal. — 2009. — Vol. 284, no. 47. — P. 32413—32424. — doi:10.1074/jbc.M109.039891.
- Chen L; Chetkovich D. M., Petralia R. S., Sweeney N. T., Kawasaki Y., Wenthold R. J., Bredt D. S., and Nicoll R. A. Stargazin regulates synaptic targeting of AMPA receptors by two distinct mechanisms (англ.) // Nature : journal. — 2000. — Vol. 408. — P. 936—943. — doi:10.1038/35050030.
- Yamazaki M; Ohno-Shosaku T., Fukaya M., Kano M., Watanabe M., and Sakimura K. A novel action of stargazin as an enhancer of AMPA receptor activity (англ.) // Neurosci Res : journal. — 2004. — Vol. 50, no. 4. — P. 369—374. — doi:10.1016/j.neures.2004.10.002.
- Priel A; Kolleker A., Ayalon G., Gillor M., Osten P., and Stern-Bach Y. Stargazin reduces desensitization and slows deactivation of the AMPA-type glutamate receptors (англ.) // J Neurosci : journal. — 2005. — Vol. 25, no. 10. — P. 2682—2686. — doi:10.1523/JNEUROSCI.4834-04.2005.
- Armstrong N; and Gouaux E. Mechanisms for activation and antagonism of an AMPA-sensitive glutamate receptor: crystal structures of the GluR2 ligand binding core (англ.) // Neuron : journal. — Cell Press, 2000. — Vol. 28, no. 1. — P. 165—181. — doi:10.1016/S0896-6273(00)00094-5.
- Coquelle T; Christensen J. K., Banke T. G., Madsen U., Schousboe A., and Pickering D. S. Agonist discrimination between AMPA receptor subtypes (англ.) // Neuroreport : journal. — 2000. — Vol. 11, no. 12. — P. 2643—2648.
- Donevan SD; Rogawski M. A. Allosteric regulation of alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionate receptors by thiocyanate and cyclothiazide at a common modulatory site distinct from that of 2,3-benzodiazepines (англ.) // Neuroscience : journal. — Elsevier, 1998. — Vol. 87, no. 3. — P. 615—629. — doi:10.1016/S0306-4522(98)00109-2.
- Banke TG; Schousboe A., and Pickering D. S. Comparison of the agonist binding site of homomeric, heteromeric, and chimeric GluR1(o) and GluR3(o) AMPA receptors (англ.) // J Neurosci Res : journal. — 1997. — Vol. 49, no. 2. — P. 176—185. — doi:10.1002/(SICI)1097-4547(19970715)49:2<176::AID-JNR6>3.0.CO;2-6.
- Dawson TL; Nicholas R. A., and Dingledine R. Homomeric GluR1 excitatory amino acid receptors expressed in Xenopus oocytes (англ.) // Mol Pharmacol : journal. — 1990. — Vol. 38, no. 6. — P. 779—784.
- Jin R; Horning M., Mayer M. L., and Gouaux E. Mechanism of activation and selectivity in a ligand-gated ion channel: structural and functional studies of GluR2 and quisqualate (англ.) // Biochemistry : journal. — 2002. — Vol. 41, no. 52. — P. 15635—15643. — doi:10.1021/bi020583k.
- Zhang W; Robert A., Vogensen S. B., and Howe J. R. The relationship between agonist potency and AMPA receptor kinetics (англ.) // Biophys J : journal. — 2006. — Vol. 91, no. 4. — P. 1336—1346. — doi:10.1529/biophysj.106.084426.
- Schiffer HH; Swanson G. T., and Heinemann S. F. Rat GluR7 and a carboxyterminal splice variant, GluR7b, are functional kainate receptor subunits with a low sensitivity to glutamate (англ.) // Neuron : journal. — 1997. — Vol. 19, no. 5. — P. 1141—1146. — doi:10.1016/S0896-6273(00)80404-3.
- Nakanishi N; Shneider N. A., and Axel R. A family of glutamate receptor genes: evidence for the formation of heteromultimeric receptors with distinct channel properties (англ.) // Neuron : journal. — Cell Press, 1990. — Vol. 5, no. 5. — P. 569—581. — doi:10.1016/0896-6273(90)90212-X.
- Vogensen SB; Jensen H. S., Stensbøl T. B., Frydenvang K., Bang-Andersen B., Johansen T. N., Egebjerg J., and Krogsgaard-Larsen P. Resolution, configurational assignment, and enantiopharmacology of 2-amino-3-[3-hydroxy-5-(2-methyl-2H- tetrazol-5-yl)isoxazol-4-yl]propionic acid, a potent GluR3- and GluR4-preferring AMPA receptor agonist (англ.) // Chirality : journal. — 2000. — Vol. 12, no. 10. — P. 705—713. — doi:10.1002/1520-636X(2000)12:10<705::AID-CHIR2>3.0.CO;2-9.
- Holm MM; Lunn M. L., Traynelis S. F., Kastrup J. S., and Egebjerg J. Structural determinants of agonist-specific kinetics at the ionotropic glutamate receptor 2 (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2005. — Vol. 102, no. 34. — P. 12053—12058. — doi:10.1073/pnas.0505522102.
- Kizelsztein P; Eisenstein M., Strutz N., Hollmann M., and Teichberg VІ. Mutant Cycle Analysis of the Active and Desensitized States of an AMPA Receptor Induced by Willardiines (англ.) // Biochemistry : journal. — 2000. — Vol. 39, no. 42. — P. 12819—12827. — doi:10.1021/bi000962i.
- Greenwood JR; Mewett K. N., Allan R. D., Martín B. O., and Pickering D. S. 3-hydroxypyridazine 1-oxides as carboxylate bioisosteres: a new series of subtype-selective AMPA receptor agonists (англ.) // Neuropharmacology : journal. — 2006. — Vol. 51, no. 1. — P. 52—9. — doi:10.1016/j.neuropharm.2006.02.013.
- Bjerrum EJ; Kristensen A. S., Pickering D. S., Greenwood J. R., Nielsen B., Liljefors T., Schousboe A., Bra¨uner-Osborne H., and Madsen U. Design, Synthesis, and Pharmacology of a Highly Subtype-Selective GluR1/2 Agonist, (RS)-2-Amino-3-(4-chloro-3-hydroxy-5-isoxazolyl)propionic Acid (Cl-HIBO) (англ.) // J Med Chem : journal. — 2003. — Vol. 46, no. 11. — P. 2246—2249. — doi:10.1021/jm020588f.
- Campiani G; Morelli E., Nacci V., Fattorusso C., Ramunno A., Novellino E., Greenwood J., Liljefors T., Griffiths R., Sinclair C., Reavy H., Kristensen A. S., Pickering D. S., Schousboe A., Cagnotto A., Fumagalli E., and Mennini T. Characterization of the 1H-cyclopentapyrimidine-2,4(1H,3H)-dione derivative (S)-CPW399 as a novel, potent, and subtype-selective AMPA receptor full agonist with partial desensitization properties (англ.) // J Med Chem : journal. — 2001. — Vol. 44, no. 26. — P. 4501—4504. — doi:10.1021/jm015552m.
- Stensbøl TB; Borre L., Johansen T. N., Egebjerg J., Madsen U., Ebert B., and Krogsgaard-Larsen P. Resolution, absolute stereochemistry and molecular pharmacology of the enantiomers of ATPA (англ.) // Eur J Pharmacol : journal. — 1999. — Vol. 380, no. 2—3. — P. 153—162. — doi:10.1016/S0014-2999(99)00512-9.
- Strange M; Bräuner-Osborne H., and Jensen A. A. Functional characterisation of homomeric ionotropic glutamate receptors GluR1-GluR6 in a fluorescence-based high throughput screening assay (англ.) // Comb Chem High Throughput Screen : journal. — 2006. — Vol. 9, no. 2. — P. 147—158. (недоступная ссылка)
- Brehm L; Greenwood J. R., Hansen K. B., Nielsen B., Egebjerg J., Stensbøl T. B., Bräuner-Osborne H., Sløk F. A., Kronborg T. T., and Krogsgaard-Larsen P. (S)-2-Amino-3-(3-hydroxy-7,8-dihydro-6H-cyclohepta[d]isoxazol-4-yl)propion ic acid, a potent and selective agonist at the GluR5 subtype of ionotropic glutamate receptors. Synthesis, modeling, and molecular pharmacology (англ.) // J Med Chem : journal. — 2003. — Vol. 46, no. 8. — P. 1350—1358. — doi:10.1021/jm0204441.
- Stensbøl TB; Jensen H. S., Nielsen B., Johansen T. N., Egebjerg J., Frydenvang K., and Krogsgaard-Larsen P. Stereochemistry and molecular pharmacology of (S)-thio-ATPA, a new potent and selective GluR5 agonist (англ.) // Eur J Pharmacol : journal. — 2001. — Vol. 411, no. 3. — P. 245—253. — doi:10.1016/S0014-2999(00)00916-X.
- Jensen AA; Christesen T., Bølcho U., Greenwood J. R., Postorino G., Vogensen S. B., Johansen T. N., Egebjerg J., Bra¨uner-Osborne H., and Clausen R. P. Functional Characterization of Tet-AMPA [Tetrazolyl-2-amino-3-(3-hydroxy-5-methyl- 4-isoxazolyl)propionic Acid] Analogues at Ionotropic Glutamate Receptors GluR1−GluR4. The Molecular Basis for the Functional Selectivity Profile of 2-Bn-Tet-AMPA (англ.) // J Med Chem : journal. — 2007. — Vol. 50, no. 17. — P. 4177—4185. — doi:10.1021/jm070532r.
- Szymańska Е; Pickering D. S., Nielsen B., and Johansen T. N. 3-Substituted phenylalanines as selective AMPA- and kainate receptor ligands (англ.) // Bioorg Med Chem : journal. — 2009. — Vol. 17, no. 17. — P. 6390—6401. — doi:10.1016/j.bmc.2009.07.021.
- Kott S; Sager C., Tapken D., Werner M., and HollmannM. Comparative analysis of the pharmacology of GluR1 in complex with transmembrane AMPA receptor regulatory proteins gamma2, gamma3, gamma4, and gamma8 (англ.) // Neuroscience : journal. — Elsevier, 2009. — Vol. 158, no. 1. — P. 78—88. — doi:10.1016/j.neuroscience.2007.12.047.
- Tygesen CK; Jørgensen M., and Andersen P. H. The importance of two specific domains in ligand binding to the AMPA/kainate glutamate receptors GluR2 and GluR6 (англ.) // FEBS Lett : journal. — 1995. — Vol. 363, no. 1—2. — P. 184—188. — doi:10.1016/0014-5793(95)00315-Z.
- Varney MA; Rao S. P., Jachec C., Deal C., Hess S. D., Daggett L. P., Lin F., Johnson E. C., and Veliçelebi G. Pharmacological characterization of the human ionotropic glutamate receptor subtype GluR3 stably expressed in mammalian cells (англ.) // J Pharmacol Exp Ther : journal. — 1998. — Vol. 285, no. 1. — P. 358—370.
- Andersen PH; Tygesen C. K., Rasmussen J. S., Søegaard-Nielsen L., Hansen A., Hansen K., Kiemer A., and Stidsen C. E. Stable expression of homomeric AMPA-selective glutamate receptors in BHK cells (англ.) // Eur J Pharmacol : journal. — 1996. — Vol. 311, no. 1. — P. 95—100. — doi:10.1016/0014-2999(96)00399-8.
- Stein E; Cox J. A., Seeburg P. H., and Verdoorn T. A. Complex pharmacological properties of recombinant alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate receptor subtypes (англ.) // Mol Pharmacol : journal. — 1992. — Vol. 42. — P. 864—871.
- Kasper C; Pickering D. S., Mirza O., Olsen L., Kristensen A. S., Greenwood J. R., Liljefors T., Schousboe A., Wätjen F., Gajhede M., Sigurskjold B. W., and Kastrup J. S. The Structure of a Mixed GluR2 Ligand-binding Core Dimer in Complex with (S)-Glutamate and the Antagonist (S)-NS1209 (англ.) // J Mol Biol : journal. — 2006. — Vol. 357, no. 4. — P. 1184—1201. — doi:10.1016/j.jmb.2006.01.024.
- Prescott C; Weeks A. M., Staley K. J., and Partin K. M. Kynurenic acid has a dual action on AMPA receptor responses (англ.) // Neurosci Lett : journal. — 2006. — Vol. 402, no. 1—2. — P. 108—112. — doi:10.1016/j.neulet.2006.03.051.
- Simmons RM; Li D. L., Hoo K. H., Deverill M., Ornstein P. L., and Iyengar S. Kainate GluR5 receptor subtype mediates the nociceptive response to formalin in the rat (англ.) // Neuropharmacology : journal. — 1998. — Vol. 37, no. 1. — P. 25—36. — doi:10.1016/S0028-3908(97)00188-3.
- Jones CK; Alt A., Ogden A. M., Bleakman D., Simmons R. M., Iyengar S., Dominguez E., Ornstein P. L., and Shannon H. E. Antiallodynic and antihyperalgesic effects of selective competitive GLUK5 (GluR5) ionotropic glutamate receptor antagonists in the capsaicin and carrageenan models in rats (англ.) // J Pharmacol Exp Ther : journal. — 2006. — Vol. 319, no. 1. — P. 396—404. — doi:10.1124/jpet.106.105601.
- Bleakman D; Ogden A. M., Ornstein P. L., and Hoo K. Pharmacological characterization of a GluR6 kainate receptor in cultured hippocampal neurons (англ.) // Eur J Pharmacol : journal. — 1999. — Vol. 378, no. 3. — P. 331—337. — doi:10.1016/S0014-2999(99)00478-1.
- Dolman NP; More J. C., Alt A., Knauss J. L., Pentika¨inen O. T., Glasser C. R., Bleakman D., Mayer M. L., Collingridge G. L., and Jane D. E. Synthesis and pharmacological characterization of N3-substituted willardiine derivatives: role of the substituent at the 5-position of the uracil ring in the development of highly potent and selective GLUK5 kainate receptor antagonists (англ.) // J Med Chem : journal. — 2007. — Vol. 50, no. 7. — P. 1558—1570. — doi:10.1021/jm061041u.
- Gitto R; Barreca M. L., De Luca L., De Sarro G., Ferreri G., Quartarone S., Russo E., Constanti A., and Chimirri A. Discovery of a novel and highly potent noncompetitive AMPA receptor antagonist (англ.) // J Med Chem : journal. — 2003. — Vol. 46, no. 1. — P. 197—200. — doi:10.1021/jm0210008.
- Cokić B; and Stein V. Stargazin modulates AMPA receptor antagonism (неопр.) // Neuropharmacology. — 2008. — Т. 54, № 7. — С. 1062—1070. — doi:10.1016/j.neuropharm.2008.02.012.
- Balannik V; Menniti F. S., Paternain A. V., Lerma J., and Stern-Bach Y. Molecular mechanism of AMPA receptor noncompetitive antagonism (англ.) // Neuron : journal. — Cell Press, 2005. — Vol. 48, no. 2. — P. 279—288. — doi:10.1016/j.neuron.2005.09.024.
- Bleakman D; Ballyk B. A., Schoepp D. D., Palmer A. J., Bath C. P., Sharpe E. F., Woolley M. L., Bufton H. R., Kamboj R. K., Tarnawa I., and Lodge D. Activity of 2,3-benzodiazepines at native rat and recombinant human glutamate receptors in vitro: stereospecificity and selectivity profiles (англ.) // Neuropharmacology : journal. — 1996. — Vol. 35, no. 12. — P. 1689—1702. — doi:10.1016/S0028-3908(96)00156-6.
- Johansen TH; Chaudhary A and Verdoorn T. A. Interactions among GYKI-52466, cyclothiazide, and aniracetam at recombinant AMPA and kainate receptors (англ.) // Mol Pharmacol : journal. — 1995. — Vol. 48, no. 5. — P. 946—955.
- Cotton JL; and Partin K. M. The contributions of GluR2 to allosteric modulation of AMPA receptors (англ.) // Neuropharmacology : journal. — 2000. — Vol. 39, no. 1. — P. 21—31. — doi:10.1016/S0028-3908(99)00105-7.
- Andersen PH; Tygesen C. K., Rasmussen J. S., Søegaard-Nielsen L., Hansen A., Hansen K., Kiemer A., and Stidsen C. E. Stable expression of homomeric AMPAselective glutamate receptors in BHK cells (англ.) // Eur J Pharmacol : journal. — 2006. — Vol. 311, no. 1. — P. 95—100. — doi:10.1016/0014-2999(96)00399-8.
- Kiskin NI; Kryshtal’ OA, Tsyndrenko AIa, Volkova T. M., and Grishin E. V. Argiopine, argiopinines and pseudoargiopinines—blockers of the glutamate receptors in hippocampal neurons (англ.) // Neirofiziologiia : journal. — 1989. — Vol. 21, no. 6. — P. 525—532. — doi:10.1007/BF01051949.
- Herlitze S; Raditsch M., Ruppersberg J. P., Jahn W., Monyer H., Schoepfer R., and Witzemann V. Argiotoxin detects molecular differences in AMPA receptor channels (англ.) // Neuron : journal. — Cell Press, 1993. — Vol. 10, no. 6. — P. 1131—1140. — doi:10.1016/0896-6273(93)90061-U.
- Brackley PT; Bell D. R., Choi S. K., Nakanishi K., and Usherwood P. N. Selective antagonism of native and cloned kainate and NMDA receptors by polyamine-containing toxins (англ.) // J Pharmacol Exp Ther : journal. — 1993. — Vol. 266, no. 3. — P. 1573—1580.
- Blaschke M; Keller B. U., Rivosecchi R., Hollmann M., Heinemann S., and Konnerth A. A single amino acid determines the subunit-specific spider toxin block of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionate/kainate receptor channels (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1993. — Vol. 90, no. 14. — P. 6528—6532.
- Washburn MS; and Dingledine R. Block of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors by polyamines and polyamine toxins (англ.) // J Pharmacol Exp Ther : journal. — 1996. — Vol. 278, no. 2. — P. 669—678.
- Kromann H; Krikstolaityte S., Andersen A. J., Andersen K., Krogsgaard-Larsen P., Jaroszewski J. W., Egebjerg J., and Strømgaard K. Solid-Phase Synthesis of Polyamine Toxin Analogues: Potent and Selective Antagonists of Ca2+-Permeable AMPA Receptors (англ.) // J Med Chem : journal. — 2002. — Vol. 45, no. 26. — P. 5745—5754. — doi:10.1021/jm020314s.
- Magazanik LG; Buldakova S. L., Samoilova M. V., Gmiro V. E., Mellor I. R., and Usherwood P. N. Block of open channels of recombinant AMPA receptors and native AMPA/kainate receptors by adamantane derivatives (англ.) // J Physiol : journal. — 1997. — Vol. 505, no. Pt 3. — P. 655—663. (недоступная ссылка)
- Schlesinger F; Tammena D., Krampfl K., and Bufler J. Two mechanisms of action of the adamantane derivative IEM-1460 at human AMPA-type glutamate receptors (англ.) // Br J Pharmacol : journal. — 2005. — Vol. 145, no. 5. — P. 656—663. — doi:10.1038/sj.bjp.0706233.
- Lauterborn JC; Lynch G., Vanderklish P., Arai A., and Gall C. M. Positive modulation of AMPA receptors increases neurotrophin expression by hippocampal and cortical neurons (англ.) // J Neurosci : journal. — 2000. — Vol. 20, no. 1. — P. 8—21.
- Ahmed HА; and Oswald R. E. Piracetam Defines a New Binding Site for Allosteric Modulators of α-Amino-3-hydroxy-5-methyl-4-isoxazole-propionic Acid (AMPA) Receptors (англ.) // J Med Chem : journal. — 2010. — Vol. 53, no. 5. — P. 2197—2203. — doi:10.1021/jm901905j.
- Arai AC; and Kessler M. Pharmacology of ampakine modulators: from AMPA receptors to synapses and behavior (англ.) // Curr Drug Targets : journal. — 2007. — Vol. 8, no. 5. — P. 583—602. (недоступная ссылка)
- Baumbarger PJ; Muhlhauser M., Zhai J., Yang C. R., and Nisenbaum E. S. Positive modulation of alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors in prefrontal cortical pyramidal neurons by a novel allosteric potentiator (англ.) // J Pharmacol Exp Ther : journal. — 2001. — Vol. 298, no. 1. — P. 86.
- Arai AC; Kessler M., Rogers G., and Lynch G. Effects of the potent ampakine CX614 on hippocampal and recombinant AMPA receptors: interactions with cyclothiazide and GYKI 52466 (англ.) // Mol Pharmacol : journal. — 2000. — Vol. 58, no. 4. — P. 802—813.
- Erreger K; Chen P. E., Wyllie D. J., and Traynelis S. F. Glutamate receptor gating (неопр.) // Crit Rev Neurobiol. — 2004. — Т. 16, № 3. — С. 187—224. — doi:10.1615/CritRevNeurobiol.v16.i3.
- Partin KM; Bowie D., and Mayer M. L. Structural determinants of allosteric regulation in alternatively spliced AMPA receptors (англ.) // Neuron : journal. — Cell Press, 1995. — Vol. 14, no. 4. — P. 833—843. — doi:10.1016/0896-6273(95)90227-9.
- Krampfl K; Schlesinger F., Wolfes H., Dengler R., and Bufler J. Functional diversity of recombinant human AMPA type glutamate receptors: possible implications for selective vulnerability of motor neurons (англ.) // J Neurol Sci : journal. — 2001. — Vol. 191, no. 1. — P. 19—23.
- Quirk JC; Siuda E. R., and Nisenbaum E. S. Molecular determinants responsible for differences in desensitization kinetics of AMPA receptor splice variants (англ.) // J Neurosci : journal. — 2004. — Vol. 24, no. 50. — P. 11416—11420. — doi:10.1523/JNEUROSCI.2464-04.2004.
- Robert A; and Howe J. R. How AMPA receptor desensitization depends on receptor occupancy (англ.) // J Neurosci : journal. — 2003. — Vol. 23, no. 3. — P. 847—858.
- Banke TG; Schousboe A., and Pickering D. S. Comparison of the agonist binding site of homomeric, heteromeric, and chimeric GluR1(o) and GluR3(o) AMPA receptors (англ.) // J Neurosci Res : journal. — 2001. — Vol. 49, no. 2. — P. 176—185. — doi:10.1002/(SICI)1097-4547(19970715)49:2<176::AID-JNR6>3.0.CO;2-6.
- Grosskreutz J; Zoerner A., Schlesinger F., Krampfl K., Dengler R., and Bufler J. Kinetic properties of human AMPA-type glutamate receptors expressed in HEK293 cells (англ.) // Eur J Neurosci : journal. — 2003. — Vol. 17, no. 6. — P. 1173—1178. — doi:10.1046/j.1460-9568.2003.02531.x.
- Lomeli H; Mosbacher J., Melcher T., Ho¨ger T., Geiger J. R., Kuner T., Monyer H., Higuchi M., Bach A., and Seeburg P. H. Control of kinetic properties of AMPA receptor channels by nuclear RNA editing (англ.) // Science : journal. — 1994. — Vol. 266, no. 5191. — P. 1709—1713. — doi:10.1126/science.7992055.
- Malinow R. AMPA receptor trafficking and long-term potentiation (англ.) // Philos Trans R Soc Lond B Biol Sci : journal. — 2003. — Vol. 358. — P. 707—714. — doi:10.1098/rstb.2002.1233.
- Mauceri D; Cattabeni F., Di Luca M., and Gardoni, F. Calcium/calmodulin-dependent protein kinase II phosphorylation drives synapse-associated protein 97 into spines (англ.) // J Biol Chem : journal. — 2004. — Vol. 279, no. 22. — P. 23813—23821. — doi:10.1074/jbc.M402796200.
- Wu H; Nash J. E., Zamorano P., and Garner C. C. Interaction of SAP97 with minus-end-directed actin motor myosin VI. Implications for AMPA receptor trafficking (англ.) // J Biol Chem : journal. — 2002. — Vol. 277. — P. 30928—30934. — doi:10.1074/jbc.M203735200.
- Zhu JJ; Qin Y., Zhao M., Van Aelst L., and Malinow R. Ras and Rap control AMPA receptor trafficking during synaptic plasticity (англ.) // Cell : journal. — Cell Press, 2002. — Vol. 110, no. 4. — P. 443—455. — doi:10.1016/S0092-8674(02)00897-8.
- Borgdorff AJ; and Choquet D. Regulation of AMPA receptor lateral movements (англ.) // Nature. — 2002. — Vol. 417, no. 6889. — P. 649—653. — doi:10.1038/nature00780.
- Park M; Penick E. C., Edwards J. G., Kauer JA and Ehlers M. D. Recycling endosomes supply AMPA receptors for LTP (англ.) // Science. — 2004. — Vol. 305, no. 5692. — P. 1972—1975. — doi:10.1126/science.1102026.
- Makino H; and Malinow R. AMPA receptor incorporation into synapses during LTP: the role of lateral movement and exocytosis (англ.) // Neuron : journal. — Cell Press, 2009. — Vol. 64, no. 3. — P. 381—390. — doi:10.1016/j.neuron.2009.08.035.
- Howard MA; Elias G. M., Elias L. A., Swat W., and Nicoll R. A. The role of SAP97 in synaptic glutamate receptor dynamics (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2010. — Vol. 107, no. 8. — P. 3805—3810. — doi:10.1073/pnas.0914422107.
- Wang Z; Edwards J. G., Riley N., Provance DW Jr, Karcher R., Li X. D., Davison I. G., Ikebe M., Mercer J. A., Kauer JA and Ehlers M. D. Myosin Vb mobilizes recycling endosomes and AMPA receptors for postsynaptic plasticity (англ.) // Cell : journal. — Cell Press, 2008. — Vol. 135, no. 3. — P. 535—548. — doi:10.1016/j.cell.2008.09.057.
- Beattie EC; Carroll R. C., Yu X., Morishita W., Yasuda H., Zastrow M and Malenka R. C. Regulation of AMPA receptor endocytosis by a mechanism shared with LTD (англ.) // Nat Neurosci : journal. — 2000. — Vol. 3, no. 12. — P. 1291—1300. — doi:10.1038/81823.
- Jung N; and Haucke V. Clathrin-mediated endocytosis at synapses (неопр.) // Traffic. — 2007. — Т. 8, № 9. — С. 1129—1136. — doi:10.1111/j.1600-0854.2007.00595.x.
- Lu W; and Ziff E. PICK1 interacts with ABP/GRIP to regulate AMPA Receptor Trafficking (англ.) // Neuron : journal. — Cell Press, 2005. — Vol. 47, no. 3. — P. 407—421. — doi:10.1016/j.neuron.2005.07.006.
- Ehlers M. D. Reinsertion or degradation of AMPA receptors determined by activity-dependent endocytic sorting (англ.) // Neuron : journal. — Cell Press, 2000. — Vol. 28, no. 2. — P. 511—525. — doi:10.1016/S0896-6273(00)00129-X.
