Сплайсинг
Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определённых нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в «зрелой» молекуле, в ходе процессинга РНК. Наиболее часто этот процесс встречается при созревании матричной, или информационной, РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК удаляются участки, не кодирующие белок (интроны) и соединяются друг с другом кодирующие аминокислотную последовательность участки — экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой считываются (транслируются) белки клетки. Большинство генов прокариот, кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК встречается редко. У представителей эукариот, бактерий и архей встречается также сплайсинг транспортных РНК (тРНК)[1] и других некодирующих РНК.
Введение
Работа Шарпа и Робертса, опубликованная в 1977 году, показала, что гены высших организмов имеют «прерывистую» структуру: кодирующие отрезки гена перемежаются с некодирующей ДНК, которая не используется при экспрессии генов. «Прерывистая» структура гена была обнаружена, когда аденовирусная мРНК была гибридизована с фрагментами одиночной цепи ДНК. В результате выяснилось, что мРНК-участки этих гибридных двухцепочечных молекул мРНК-ДНК содержат 5'- и 3'-концы участков, не обладающие водородными связями. Более длинные отрезки ДНК при гибридизации закольцовывались и образовывали ответвления. Стало ясно, что эти закольцованные участки, содержащие «ненужные» последовательности, извлекаются из пре-мРНК в результате процесса, который и был назван «сплайсингом». Впоследствии было также выяснено, что прерывистая структура крайне широко распространена у эукариотических генов.
Варианты сплайсинга
В природе обнаружены несколько вариантов сплайсинга. Какой из них будет проходить в каждом случае, зависит от структуры интрона и катализатора, необходимого для реакции.
Сплайсосомные интроны
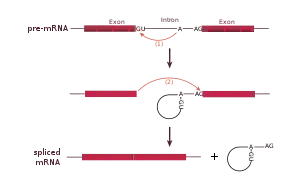
Сплайсосомные интроны часто находятся в генах, кодирующих белки. Для сплайсинга необходимо наличие специальных 3'- и 5'-последовательностей. Важная роль в защите 5'-конца мРНК от деградации экзонуклеазами принадлежит 5'-кэпу. Сплайсинг катализируется сплайсосомой — большим комплексом, состоящим из РНК и белков и включающим пять малых ядерных рибонуклеопротеидов (мяРНП). РНК-составляющая мяРНП взаимодействует с интроном и, возможно, участвует в катализе. Обнаружены два типа сплайсосом (главная и дополнительная), отличающиеся по входящим в их состав мяРНП.
Главная сплайсосома принимает участие в сплайсинге интронов, содержащих гуанин и урацил (GU) в 5'-сайте, и аденин и гуанин (AG) в 3'-сплайсинг-сайте. Она состоит из мяРНП: U1, U2, U4, U5 и U6.
Альтернативный сплайсинг
Пре-мРНК некоторых генов эукариот могут подвергаться альтернативному сплайсингу. При этом интроны в составе пре-мРНК вырезаются в разных альтернативных комбинациях, при которых вырезаются и некоторые экзоны. Разные варианты альтернативного сплайсинга одной пре-мРНК могут осуществляться в разные периоды развития организма или в разных тканях, а также у разных особей одного вида[2]. Некоторые из продуктов альтернативного сплайсинга пре-мРНК нефункциональны (такой вариант альтернативного сплайсинга осуществляется у дрозофилы при определении пола), но нередко в результате альтернативного сплайсинга пре-мРНК одного гена образуются многочисленные мРНК и их белковые продукты.[3]
Показано, что у человека 94 % генов подвержено альтернативному сплайсингу (у остальных 6 % генов нет интронов). Геном круглого червя Caenorhabditis elegans по количеству генов практически не отличается от генома человека, однако альтернативному сплайсингу подвергаются пре-мРНК только 15 % генов. Таким образом, альтернативный сплайсинг позволяет увеличить разнообразие белковых продуктов генов, не увеличивая пропорционально этому размер генома, в том числе не создавая дополнительных копий генов. Биологический смысл альтернативного сплайсинга для многоклеточных эукариот состоит в том, что он, по-видимому, является ключевым механизмом увеличения разнообразия белков, а также позволяет осуществлять сложную систему регуляции экспрессии генов, в том числе тканеспецифической[4].
Автосплайсинг
РНК тетрахимены обладает рибозимной активностью и может сплайсировать сама себя, замыкаясь в кольцо и одним своим концом вырезая интроны из другого.
Транс-сплайсинг
Особая форма сплайсинга у эукариот, при которой концом к концу соединяются и лигируются экзоны двух разных РНК-транскриптов.
Примечания
Посттранскрипционные модификации | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ядерные |
| ||||||||
| Цитозольные |
| ||||||||
