Процессинг РНК
Созревание РНК или процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК.
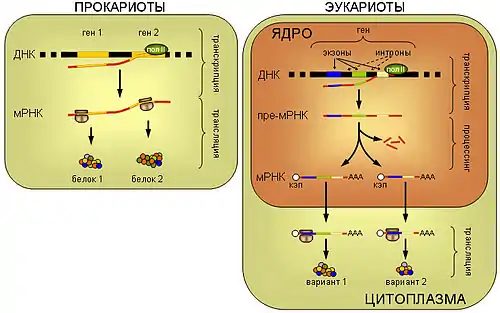
ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция) пространственно не отделён от транскрипции и может происходить ещё до завершения синтеза мРНК РНК-полимеразой. Прокариотические мРНК часто полицистронные, то есть содержат несколько независимых генов.
ЭУКАРИОТЫ. мРНК эукариот синтезируется в виде предшественника, пре-мРНК, претерпевающего затем сложное стадийное созревание — процессинг, включающий присоединение кэп-структуры к 5'-концу молекулы, присоединение нескольких десятков остатков аденина к её 3'-концу (полиаденилирование), выщепление незначащих участков — интронов и соединение друг с другом значащих участков — экзонов (сплайсинг). При этом соединение экзонов одной и той же пре-мРНК может проходить разными способами, приводя к образованию разных зрелых мРНК, и в конечном итоге разных вариантов белка (альтернативный сплайсинг). Только мРНК, успешно прошедшая процессинг, экспортируется из ядра в цитоплазму и вовлекается в трансляцию.
В зависимости от типа РНК (матричные, рибосомные, транспортные, малые ядерные) их предшественники подвергаются разным последовательным модификациям. Например, предшественники матричных РНК подвергаются кэпированию, сплайсингу, полиаденилированию, метилированию и иногда редактированию.
Процессинг мРНК
Кэпирование

Кэпирование представляет собой присоединение к 5'-концу транскрипта 7-метилгуанозина через необычный для РНК 5',5'-трифосфатный мостик, а также метилирование остатков рибозы двух первых нуклеотидов. Процесс кэпирования происходит во время синтеза молекулы пре-мРНК. Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные связи в направлении 5’→3'.[1]:221
Функции кэпа и связанных с ним белков:
- участие в сплайсинге;
- участие в процессинге 3'-конца мРНК;
- экспорт мРНК из ядра;
- защита 5'-конца транскрипта от экзонуклеаз;
- участие в инициации трансляции.
Полиаденилирование
Фермент поли(А)-полимераза присоединяет 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты. Полиаденилирование осуществляется при наличии сигнальной последовательности 5'- AAUAAA-3' на 3'-конце транскрипта, за которой следует 5'-CA-3'. Вторая последовательность является сайтом разрезания[1]:225.
Сплайсинг
После полиаденилирования мРНК подвергается сплайсингу, в ходе которого удаляются интроны (участки, которые не кодируют белки), а экзоны (участки, кодирующие белки) сшиваются и образуют единую молекулу[2]. Сплайсинг катализируется крупным нуклеопротеидным комплексом — сплайсосомой, состоящей из белков и малых ядерных РНК. Многие пре-мРНК могут быть подвергнуты сплайсингу разными путями, при этом образуются разные зрелые мРНК, кодирующие разные последовательности аминокислот (альтернативный сплайсинг).
Редактирование
Редактирование РНК — изменение содержащейся в молекуле РНК информации путём химической модификации оснований.
Метилирование
мРНК эукариот подвергаются посттранскрипционному метилированию. То есть сметению (метилированию) наружу ингибиторного гена. Наиболее распространённой модификацией является метилирование остатков аденина по положению N6 с образованием N6-метиладенозина (m6A). Этот процесс метилируют ферменты N6-аденозинметилтрансферазы, которые распознают остатки аденина в консенсусных последовательностях GAC (70 % случаев) и AAC (30 % случаев). Соответствующие деметилазы ингибируют обратный процесс деметилирования. Учитывая обратимость и динамичность процесса метилирования мРНК, а также повышенную концентрацию m6A в длинных экзонах и вокруг стоп-кодонов, предполагают, что метилирование мРНК выполняет регуляторную функцию[3].
Примечания
- Hames, David & Hooper, Nigel (2006), Instant Notes Biochemistry (3 ed.), Leeds: Taylor and Francis, ISBN 0-415-36778-6
- Lodish H. F., Berk A., Kaiser C., Krieger M., Scott M. P., Bretscher A., Ploegh H., Matsudaira P. T. Chapter 8: Post-transcriptional Gene Control // Molecular Cell .Biology (неопр.). — San Francisco: WH Freeman, 2007. — ISBN 0-7167-7601-4.
- Wang X., Lu Z., Gomez A., Hon G. C., Yue Y., Han D., Fu Y., Parisien M., Dai Q., Jia G., Ren B., Pan T., He C. N6-methyladenosine-dependent regulation of messenger RNA stability (англ.) // Nature. — 2014. — Vol. 505, iss. 7481. — P. 117—120. — doi:10.1038/nature12730. — PMID 24284625.
Посттранскрипционные модификации | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ядерные |
| ||||||||
| Цитозольные |
| ||||||||