Транскрипция (биология)
Транскри́пция (от лат. transcriptio «переписывание») — происходящий во всех живых клетках процесс синтеза РНК с использованием ДНК в качестве матрицы; перенос генетической информации с ДНК на РНК.
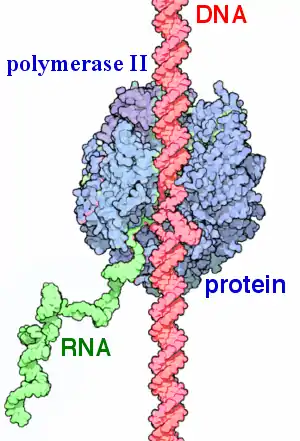


Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. РНК-полимераза движется по молекуле ДНК в направлении 3`- 5`[1].
Если говорить о транскрипции белок-кодирующих участков, то единицей транскрипции бактерий является оперон — фрагмент молекулы ДНК, состоящий из промотора (оператора, с которым связывается белок-репрессор), транскрибируемой части (которая может содержать несколько белок-кодирующих последовательностей) и терминатора. У эукариот транскрибируемая часть обычно содержит одну белок-кодирующую последовательность.
Цепочка ДНК, которая служит шаблоном для достраивания РНК, называется кодирующей или матричной. Последовательность, полученная в результате такого синтеза РНК будет идентична последовательности некодирующей цепочки ДНК (исключая замену тимина ДНК на урацил РНК) согласно принципу комплементарности.
Транскрипция про- и эукариот
В бактериях транскрипцию катализирует единственная РНК-полимераза. Она состоит из основной части из пяти субъединиц (α2ββ'ω) и σ-субъединицы (сигма-фактор), которая определяет связывание с промотором и является единственным фактором инициации транскрипции. У Escherichia coli, например, самая распространенная форма сигма-фактора — σ70.
Клетки эукариот содержат как минимум 3 РНК-полимеразы, а растения — 5, которые для инициации и элонгации требуют набора факторов. РНК-полимераза II — основной фермент эукариотических клеток, катализирующий транскрипцию белок-кодирующих мРНК (и некоторых других РНК).
В бактериях, мРНК после транскрипции никак не модифицируется, и непосредственно во время транскрипции может происходить трансляция. В эукариотических клетках мРНК модифицируется в ядре — на неё навешивается 5'-кэп и синтезируется 3'-полиА-хвост, происходит сплайсинг. Затем мРНК может попасть в цитоплазму, где будет происходит трансляция.
Процесс транскрипции
Транскрипция состоит из стадий инициации, элонгации и терминации.
Инициация
Инициация транскрипции — процесс связывания ДНК-зависимой РНК-полимеразы с промотором и образования стабильного комплекса для продолжения транскрипции.
Инициация транскрипции может быть разбита на несколько шагов[2].
- РНК-полимераза (вместе с факторами инициации транскрипции у эукариот) связывается с промотором с образованием закрытого комплекса. В такой форме внутри комплекса находится двойная спираль ДНК.
- Преобразование в открытый комплекс. Спираль ДНК на расстоянии около 13 пар нуклеотидов от точки старта транскрипции плавится, то есть цепи ДНК отделяются друг от друга. Участок разделённых спиралей ДНК называется транскрипционным пузырем.
- Разделение цепей открывает доступ к некодирующей цепочке ДНК. Первые два рибонуклеотида выравниваются с шаблонной ДНК и соединяются. Далее удлинение РНК происходит при присоединении рибонуклеотидов к 3'-концу цепочки. Соединение первых 10 нуклеотидов — неэффективный процесс, поэтому транскрипция на этой стадии часто обрывается, короткий транскрипт высвобождается и синтез начинается снова. Такая пробуксовка полимеразы называется абортивной транскрипцией.
- Как только полимеразно-промоторный комплекс образует транскрипт длиннее 10 нуклеотидов, он становится достаточно стабильным, чтобы продолжать транскрипцию и переходит в стадию элонгации. Также это называется избеганием промотора.
Инициация транскрипции — сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариот также и от более далеких участков генома — энхансеров и сайленсеров) и от наличия или отсутствия различных белковых факторов.
Элонгация
Момент перехода РНК-полимеразы от инициации транскрипции к элонгации точно не определён. Три основных биохимических события характеризуют этот переход в случае РНК-полимеразы кишечной палочки: отделение сигма-фактора, первая транслокация молекулы фермента вдоль матрицы и сильная стабилизация транскрипционного комплекса, который кроме РНК-полимеразы включает растущую цепь РНК и транскрибируемую ДНК. Эти же явления характерны и для РНК-полимераз эукариот. Переход от инициации к элонгации сопровождается разрывом связей между ферментом, промотором, факторами инициации транскрипции, а в ряде случаев — переходом РНК-полимеразы в состояние компетентности в отношении элонгации (например, фосфорилирование CTD-домена у РНК-полимеразы II). Фаза элонгации заканчивается после освобождения растущего транскрипта и диссоциации фермента от матрицы (терминация).
На стадии элонгации в ДНК расплетено примерно 18 пар нуклеотидов. Примерно 12 нуклеотидов матричной нити ДНК образует гибридную спираль с растущим концом цепи РНК. По мере движения РНК-полимеразы по матрице впереди неё происходит расплетание, а позади — восстановление двойной спирали ДНК. Одновременно освобождается очередное звено растущей цепи РНК из комплекса с матрицей и РНК-полимеразой. Эти перемещения должны сопровождаться относительным вращением РНК-полимеразы и ДНК. Трудно себе представить, как это может происходить в клетке, особенно при транскрипции хроматина. Поэтому не исключено, что для предотвращения такого вращения двигающуюся по ДНК РНК-полимеразу сопровождают топоизомеразы.
Элонгация осуществляется с помощью основных элонгирующих факторов, необходимых, чтобы процесс не останавливался преждевременно[3].
В последнее время появились данные, показывающие, что регуляторные факторы также могут регулировать элонгацию. РНК-полимераза в процессе элонгации делает паузы на определённых участках гена. Особенно четко это видно при низких концентрациях субстратов. В некоторых участках матрицы длительные задержки в продвижении РНК-полимеразы, т. н. паузы, наблюдаются даже при оптимальных концентрациях субстратов. Продолжительность этих пауз может контролироваться факторами элонгации.
Терминация
У бактерий есть два механизма терминации транскрипции:
- ро-зависимый механизм, при котором белок Rho ([ро]) дестабилизирует водородные связи между матрицей ДНК и мРНК, высвобождая молекулу РНК.
- ро-независимый, при котором транскрипция останавливается, когда только что синтезированная молекула РНК формирует стебель-петлю, за которой расположено несколько урацилов (…УУУУ), что приводит к отсоединению молекулы РНК от матрицы ДНК.
Терминация транскрипции у эукариот менее изучена. Она завершается разрезанием РНК, после чего к её 3' концу фермент добавляет несколько аденинов (…АААА), от числа которых зависит стабильность данного транскрипта[4].
Транскрипционные фабрики
Существует ряд экспериментальных данных, свидетельствующих о том, что транскрипция осуществляется в так называемых транскрипционных фабриках: огромных, по некоторым оценкам, до 10 МДа комплексах, которые содержат около 8 РНК-полимераз II и компоненты последующего процессинга и сплайсинга, а также корректирования новосинтезированного транскрипта[5]. В ядре клетки происходит постоянный обмен между пулами растворимой и задействованной РНК-полимеразы. Активная РНК-полимераза задействована в таком комплексе, который в свою очередь является структурной организовывающей компактизацию хроматина единицей. Последние данные[6] свидетельствуют о том, что транскрипционные фабрики существуют и в отсутствие транскрипции, они фиксированы в клетке (пока не ясно, взаимодействуют ли они с ядерным матриксом клетки или нет) и представляют собой независимый ядерный субкомпартмент. Комплекс транскрипционных фабрик, содержащих РНК полимеразу I, II или III, был проанализирован с помощью масс-спектрометрии.[7]
Обратная транскрипция
Некоторые вирусы (такие как вирус иммунодефицита человека, вызывающий ВИЧ-инфекцию), имеют возможность транскрибировать РНК в ДНК. ВИЧ имеет РНК-геном, который встраивается в ДНК. В результате, ДНК вируса может быть объединена с геномом клетки-хозяина. Главный фермент, ответственный за синтез ДНК из РНК, называется ревертазой. Одной из функций ревертазы является создание комплементарной ДНК (кДНК) из вирусного генома. Ассоциированный фермент рибонуклеаза H расщепляет РНК, а ревертаза синтезирует кДНК из двойной спирали ДНК. кДНК интегрируется в геном клетки-хозяина с помощью интегразы. Результатом является синтез вирусных протеинов клеткой-хозяином, которые образуют новые вирусы. В случае с ВИЧ так же программируется апоптоз (смерть клетки) Т-лимфоцитов.[8] В иных случаях клетка может остаться распространителем вирусов.
Некоторые клетки эукариот содержат фермент теломеразу, также проявляющую активность обратной транскрипции. С её помощью синтезируются повторяющиеся последовательности в ДНК. Теломераза часто активируются в раковых клетках для бесконечной дупликации генома без потери кодирующей протеины последовательности ДНК. Некоторые РНК-содержащие вирусы животных при помощи РНК-зависимой ДНК-полимеразы способны синтезировать ДНК, комплементарную по отношению к вирусной РНК. Она встраивается в геном эукариотической клетки, где может многие поколения оставаться в скрытом состоянии. При определённых условиях (например, воздействии канцерогенов) вирусные гены могут активироваться, и здоровые клетки превратятся в раковые.
Примечания
- Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts. Molecular Biology of the Cell. — 4th. — Garland Science, 2002. — ISBN 978-0-8153-3218-3, 978-0-8153-4072-0.
- James D. Watson. Molecular Biology of the Gene. — W. A. Benjamin, 1965. — 530 с.
- D. B. Nikolov, S. K. Burley. RNA polymerase II transcription initiation: A structural view (англ.) // Proceedings of the National Academy of Sciences. — 1997-01-07. — Vol. 94, iss. 1. — P. 15–22. — ISSN 1091-6490 0027-8424, 1091-6490. — doi:10.1073/pnas.94.1.15.
- Benjamin Lewin. Genes 9. — Jones & Bartlett Learning, 2008. — 912 с. — ISBN 978-0-7637-4063-4.
- Peter R. Cook. The Organization of Replication and Transcription (англ.) // Science. — 1999-06-11. — Vol. 284, iss. 5421. — P. 1790–1795. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.284.5421.1790.
- Jennifer A. Mitchell, Peter Fraser. Transcription factories are nuclear subcompartments that remain in the absence of transcription (англ.) // Genes & Development. — 2008-01-01. — Vol. 22, iss. 1. — P. 20–25. — ISSN 1549-5477 0890-9369, 1549-5477. — doi:10.1101/gad.454008.
- Svitlana Melnik, Binwei Deng, Argyris Papantonis, Sabyasachi Baboo, Ian M. Carr. The proteomes of transcription factories containing RNA polymerases I, II or III (англ.) // Nature Methods. — 2011-11. — Vol. 8, iss. 11. — P. 963–968. — ISSN 1548-7105. — doi:10.1038/nmeth.1705.
- Ирина Николаевна Колесникова. Некоторые особенности механизмов апоптоза при ВИЧ-инфекции. — Ростов-на-Дону, 2000.
| В библиографических каталогах |
|---|