Ракообразные
Ракообра́зные[1] (лат. Crustacea) — крупная группа членистоногих, в настоящее время рассматриваемая в ранге подтипа[2]. К ракообразным относятся такие широко известные животные, как крабы, омары, лангусты, широкопалый речной рак, креветки и криль. Описано около 73 000 видов[3][4]. Ракообразные освоили практически все типы водоёмов. Большинство из них являются активно передвигающимися животными, однако существуют и неподвижные формы — морские жёлуди (балянусы) и морские уточки. Некоторые ракообразные обитают на суше (мокрицы, некоторые крабы и крабоиды), а в почве влажных тропических районов встречаются рачки-бокоплавы. Ряд таксонов характеризуется паразитическим образом жизни, их хозяевами выступают водные беспозвоночные и рыбы[5]. Ракообразным посвящена наука карцинология.
| Ракообразные | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд: краб Grapsus grapsus, морская уточка Lepas sp. | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Ракообразные |
||||||||||
| Международное научное название | ||||||||||
| Crustacea Brünnich, 1772 | ||||||||||
| Классы | ||||||||||
|
||||||||||
| ||||||||||
Ракообразные, как и другие членистоногие, имеют хитиновый экзоскелет. Так как он ограничивает рост животного, экзоскелет периодически сбрасывается в ходе линьки до тех пор, пока ракообразное не достигнет нужного размера. От других членистоногих (хелицеровых, насекомых, многоножек) ракообразные отличаются наличием двуветвистых конечностей и особой формы личинки — науплиуса. Кроме того, у ракообразных одновременно присутствуют 2 пары усиков: антеннулы и антенны. Дыхание у большинства представителей осуществляется при помощи жабр, представляющих собой выросты ножек — эпиподиты[6].
Вымершие ракообразные оставили после себя многочисленные ископаемые остатки, древнейшие из которых датируются кембрийским периодом[7].
Многие ракообразные употребляются человеком в пищу, особо велико потребление креветок. Такие ракообразные, как веслоногие рачки и криль, возможно, обладают наибольшей биомассой среди всех животных на планете. Они являются важнейшим звеном пищевых цепей.
Строение и физиология
Размеры тела
Размеры и форма тела ракообразных широко варьируют. Самые мелкие ракообразные являются паразитами и относятся к группе тантулокарид; их длина тела составляет 0,15—0,3 мм[8][9]. К этой же группе относится самое мелкое членистоногое — паразитический рачок Stygotantulus stocki, длина тела которого меньше 0,1 мм[10]. Камчатский краб (Paralithodes camtschatica) достигает массы 10 кг, гигантский тасманийский краб (Pseudocarcinus gigas) — до 14 кг[11], а японский краб-паук (Macrocheira kaempferi) — до 20 кг и 3,8 м в размахе ног. Сильно видоизменённый внешний вид имеют сидячие формы с известковым панцирем, а также раки-паразиты[12].
Сегментация и конечности
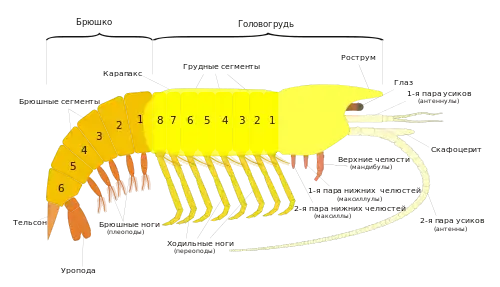
Исходно тело ракообразных включает 3 отдела: головной, грудной и брюшной. У некоторых примитивных видов грудной и брюшной отделы сегментированы почти гомономно (то есть состоят из практически одинаковых сегментов)[12]. Количество сегментов тела сильно варьирует: от 5—8 до 50. В настоящее время считается, что в процессе эволюции ракообразных, как и других членистоногих, происходило уменьшение числа сегментов. У высших раков число сегментов постоянно: акрон, четыре сегмента головы, восемь грудных сегментов и шесть брюшных[6].
- Конечности
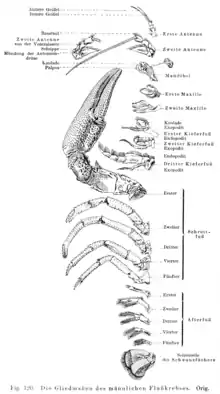
Сегменты тела несут по паре двуветвистых конечностей. В типичном случае конечность ракообразного состоит из базальной части — протоподита, несущего две ветви: наружную — экзоподит и внутреннюю — эндоподит. Протоподит включает два членика: коксоподит, обычно несущий жаберный придаток, и базиподит, к которому присоединены экзоподит и эндоподит. Экзоподит часто редуцируется, и конечность принимает одноветвистое строение. Первично конечности ракообразных выполняли несколько функций: двигательную, дыхательную, а также вспомогательную при питании, но у большинства имеет место морфофункциональная дифференциация конечностей[12].
- Голова
Голова состоит из головной лопасти — акрона и четырёх сегментов. Голова несёт придатки акрона — антенны первые (антеннулы) и конечности последующих четырёх сегментов: антенны вторые, мандибулы, или жвалы[14] (верхние челюсти) и две пары максилл (нижних челюстей)[12]. Иногда первую пару нижних челюстей называют максиллулами, а максиллами — вторую[15]. Антеннулы обычно одноветвисты и гомологичны пальпам многощетинковых червей[16]. Экзоподит второй антенны называется скафоцеритом. Усики выполняют функцию органов чувств, иногда движения, остальные головные придатки участвуют в захвате и измельчении пищи[14]. Мандибулы играют главную роль в измельчении пищи. У личинки — науплиуса — мандибула представляет собой типичную двуветвистую конечность с жевательным отростком. У взрослых редко имеется подобная форма мандибулы, обычно же обе ветви редуцируются, а протоподит с жевательным отростком образует верхнюю челюсть, к которой крепятся мышцы. Максиллы обычно имеют вид нежных листообразных ножек с жевательными отростками на протоподите и в некоторой степени редуцированными ветвями[17].
Голова может быть как слитной (синцефалон[15]), так и подразделённой на два сочленённых отдела: протоцефалон, который образуется путём слияния акрона и первого головного сегмента и несёт две первые пары антенн, и гнатоцефалон, образующийся слиянием трёх последних головных сегментов и несущий мандибулы и максиллы. Последний вариант имеет место у отрядов: жаброноги, мизиды, эвфаузиевые, десятиногие, ротоногие[14]. Ротовое отверстие спереди прикрыто непарной складкой кутикулы — верхней губой[16]. Нередко у высших раков (как, например, у речного рака) гнатоцефалон срастается с грудным отделом, формируя челюстегрудь (гнатоторакс), покрытую спинным панцирем — карапаксом. Тело высших раков подразделяется на следующие отделы: голова — протоцефалон (акрон и один сегмент), челюстегрудь — гнатоторакс (три головных и восемь грудных сегментов) и брюшко (6 сегментов и тельсон). В других случаях наблюдается слияние всей головы, не подразделённой на протоцефалон и гнатотоцефалон, с одним или несколькими грудными сегментами. Так образуется головогрудь, за которой следуют грудь и брюшко[18]. У некоторых ракообразных (например, ветвистоусых) голова вытянута в направленный вниз клюв — рострум[15].
- Грудной отдел
Грудной отдел, как и брюшной, может иметь разное число сегментов. Некоторые раки, например, жаброноги, имеют мультифункциональные брюшные конечности, у других же наблюдается разделение функций. Так, у речного рака три первые пары грудных ног — двуветвистые ногочелюсти, служащие для удерживания и отцеживания пищи, три последующие — одноветвистые ходильные и в то же время хватательные, с клешнёй на конце, однако все грудные ноги у основания несут жабры[19].
- Брюшной отдел

Брюшной отдел состоит из нескольких сегментов и тельсона; как правило, он лишён конечностей. Только у высших раков[15] на брюшке располагаются двувествистые конечности, выполняющие различные функции: у креветок — плавательную, у ротоногих раков — дыхательную, у самцов речного рака две первые пары видоизменены в копулятивные органы, а у самок редуцирована первая пара, остальные же брюшные ножки предназначены для вынашивания молоди. У большинства десятиногих последняя пара брюшных ног имеет пластинчатую форму (уроподы) и вместе с тельсоном образует пятилопастной «плавник»[19].
Ракообразные, лишённые брюшных конечностей, обычно имеют на конце тела вилочку (фурку), образованную членистыми придатками тельсона. Одновременно и вилочка, и брюшные ноги имеются только у рачка Nebalia. У крабов брюшной отдел редуцирован[20].
У некоторых паразитических ракообразных конечности тела значительно редуцируются или даже совсем исчезают (Sacculina, самки Dendrogaster)[17].
Покровы
Как и другие членистоногие, ракообразные имеют прочный хитиновый экзоскелет (кутикулу). Кутикула состоит из нескольких слоёв, её периферические слои пропитаны известью, а внутренние состоят в основном из мягкого и эластичного хитина. У мелких низших форм скелет мягкий и прозрачный[21]. Кроме того, в состав хитиновой кутикулы входят разнообразные пигменты, придающие животному покровительственную окраску. Пигменты содержатся также в гиподерме. Некоторые ракообразные способны изменять окраску за счёт изменения распределения пигментных зёрен в клетках (если пигмент сконцентрирован в центре клетки, то окраска исчезает, если же пигмент распределён в клетке равномерно, то окраска будет проявляться в покровах). Этот процесс регулируется нервно-гуморальными факторами[20].
Функция наружного скелета не исчерпывается защитой животного, к кутикуле также прикрепляются различные мышцы. Нередко для их прикрепления на нижней стороне кутикулы имеются специальные отростки в виде гребней и перекладин[22].
Подвижность таких частей тела ракообразного обеспечивается особыми мягкими мембранами, располагающимися между слитными отделами тела, сегментами или члениками конечностей и придатков. Уплотнённые участки сегментов на спинной стороне называют тергитами, а на брюшной — стернитами. Уже упоминавшийся выше карапакс представляет собой особую складку покровов. Он может иметь форму щита, двустворчатой раковины или полуцилиндра[15]. Карапакс может прикрывать различные отделы: голову, грудь (речной рак, щитень) или всё тело (дафнии, ракушковые раки), у высших раков его боковые части прикрывают жабры[20].
Внутреннее строение
Мускулатура
Мускулатура ракообразных представлена поперечно-полосатой мышечной тканью, как и у всех членистоногих. У них отсутствует единый кожно-мускульный мешок, и мышцы представлены отдельными более или менее крупными пучками. Обычно один конец мускула прикрепляется к стенке одного сегмента тела или членика конечности, другой — к стенке другого сегмента. У ракушковых рачков, имеющих двустворчатую раковину, имеется специальная замыкательная мышца, идущая поперёк тела и соединяющая две створки раковины[22].
Пищеварительная система
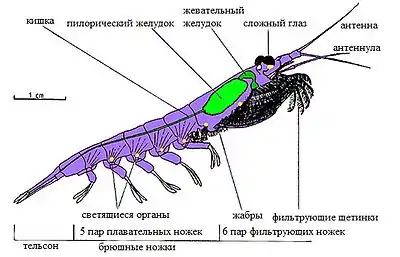
Пищеварительная система ракообразных развита хорошо, имеет вид прямой или чуть согнутой трубки[23]. Как и у всех членистоногих, она состоит из эктодермальной передней, энтодермальной средней и эктодермальной задней кишки[20].
Передняя кишка представлена пищеводом и желудком и выстлана хитиновой кутикулой. Желудок может подразделяться на жевательный (кардиальный), в котором пища измельчается при помощи жевательных пластинок — зазубренных, пропитанных известью утолщений кутикулы на стенках желудка, и пилорический, в котором пища фильтруется с помощью тонких кутикулярных выростов, образующих нечто вроде фильтра, отделы (например, у речного рака)[20][23].
В среднюю кишку впадают протоки парных печёночных придатков, представляющей собой боковые выпячивания стенки. В случае обильного развития эти придатки называются печенью. Печень ракообразных не только выделяет пищеварительные ферменты, но и всасывает переваренную пищу. Её ферменты действуют на жиры, белки и углеводы. Таким образом, функционально печень ракообразных соответствует печени и поджелудочной железе позвоночных. В печени осуществляется как полостное, так и внутриклеточное пищеварение. Между размерами средней кишки и печени имеется обратная связь[24]. У веслоногих рачков средняя кишка имеет вид простой трубки и лишена печёночных выпячиваний. В зачаточном состоянии печень имеется у некоторых ветвистоусых, у бокоплавов и равноногих печень имеет вид двух пар длинных трубчатых мешков[25].
Задняя кишка прямая, выстлана хитиновой кутикулой. Анальное отверстие открывается на брюшной стороне тельсона (анальной лопасти)[23]. Во время линьки у ракообразных, помимо наружного хитинового покрова, сбрасывается также выстилка переднего и заднего отделов. Пока новые покровы не отвердеют, рак не питается[26].
У некоторых паразитических ракообразных (например, Sacculina) кишечник целиком атрофирован[25].
Дыхательная система
Большая часть ракообразных дышит кожными жабрами, представляющими собой перистые или пластинчатые выросты — эпиподиты, отходящие от протоподитов ног. Как правило, они располагаются на грудных конечностях, лишь у ротоногих и равноногих брюшные ножки полностью превращены в жабры. У десятиногих ракообразных жабры также формируются на стенке тела в жаберных полостях под карапаксом, постепенно переходя с протоподитов на стенку тела. При этом жабры у десятиногих располагаются тремя продольными рядами: в первом ряду жабры сохраняют первичное расположение на протоподитах тела, во втором они сидят на местах соединения протоподитов с телом, в третьем — уже полностью перешли на боковую стенку тела. В жабрах продолжается полость тела, в которую попадает гемолимфа. Газообмен происходит через очень нежную кутикулу жабр[25].
Поток воды в жабрах осуществляется следующим образом. Вода входит в жаберные полости с одного конца тела через щель между карапаксом и телом, а выталкивается с другого, причём направление тока воды может меняться. Проведению воды также способствует движение особых отростков второй пары максилл, совершающих до 200 машущих движений в 1 мин[25].
У многих мелких рачков с тонким карапаксом нет жабр, а дыхание идёт через всю поверхность тела. У сухопутных ракообразных имеются специальные приспособления для дыхания атмосферным кислородом, например, псевдотрахеи (глубокие впячивания) на брюшных ножках мокриц. Полость конечности заполнена гемолимфой, омывающей впячивания и осуществляющей газообмен[27]. Сухопутные крабы дышат кислородом, растворённым в воде, покрывающей тонкой плёнкой мембраны жаберной полости и защищённой от испарения карапаксом. Однако для дыхания сухопутным ракообразным всё равно необходима повышенная влажность воздуха[26].
Кровеносная система

Как и все членистоногие, ракообразные имеют смешанную полость тела (миксоцель) и незамкнутую кровеносную систему (то есть гемолимфа течёт по сосудам и синусам миксоцеля). Сердце располагается над кишечником, на спинной стороне тела[15] и находится вблизи органов дыхания (если жабры имеются только на грудных ножках, сердце в грудном отделе и т. д.). У наиболее примитивных ракообразных сердце метамерное, многокамерное, и представлено длинной трубкой, идущей вдоль всего тела (некоторые жаброногие) и имеющей в каждом сегменте (камере) пару остий (отверстий). У других ракообразных сердце укорачивается: у водяных блох сердце укорочено до степени бочонковидного мешочка с одной парой остий, у десятиногих сердце — небольшой мешочек с тремя парами остий. Среди высших раков есть представители как с длинным, так и с укороченным сердцем[27].
Сердце ракообразных находится в перикардиальном синусе миксоцеля. Оттуда гемолимфа через остии поступает в сердце. При сокращении камер сердца клапаны остий закрываются, клапаны сердечных камер открываются, и гемолимфа изгоняется в артерии: переднюю и заднюю[27]. Оттуда гемолимфа изливается в промежутки между органами, где отдаёт кислород и насыщается углекислым газом. Функцию газообмена она выполняет благодаря наличию дыхательных пигментов — гемоцианина (у высших раков) или гемоглобина (у веслоногих, ракушковых, усоногих и жаброногих раков), связывающих кислород[28]. Частично гемолимфа омывает почки, где освобождается от продуктов обмена. Далее она собирается в систему венозных сосудов, доставляется в жаберную систему капилляров, отдаёт углекислый газ и насыщается кислородом. Затем выносящие жаберные сосуды доставляют её в перикардиальный синус[29].
Степень развития кровеносной системы связана с развитием дыхательной системы. У мелких ракообразных, осуществляющих газообмен через стенку тела, от кровеносной системы остаётся лишь сердце или она исчезает совсем.
Выделительная система

Выделительная система ракообразных представлена почками, являющимися видоизменёнными целомодуктами. Каждая почка состоит из мешочка целомического происхождения и извитого выделительного канальца, который может расширяться, образуя мочевой пузырь. В зависимости от места, где открываются выделительные поры, выделяют два типа почек: антеннальные (первая пара; выделительные поры открываются у основания вторых антенн) и максиллярные (вторая пара; у основания второй пары максилл). Высшие раки во взрослом состоянии имеют только антеннальные почки, все остальные — только максиллярные[30]. Обе пары почек имеются только у уже упоминавшегося рачка Nebalia из группы высших раков, а также у морских ракушковых рачков. У остальных ракообразных имеется лишь одна из двух пар почек, причём в процессе онтогенеза происходит их смена: если в личиночном состоянии функционируют максиллярные железы, то во взрослом — антеннальные. По-видимому, изначально ракообразные имели 2 пары почек, как Nebalia, но в ходе последующей эволюции сохранили лишь одну[31].
Нервная система

Нервная система ракообразных, как и всех членистоногих, представлена парными надглоточными ганглиями, нервным кольцом и брюшной нервной цепочкой. У примитивных жаброногих раков нервная система лестничного типа: парные ганглии в сегментах широко расставлены и соединяются комиссурами. У большинства ракообразных брюшные стволы сблизились, правый и левый ганглии слились, комиссуры исчезли, и только двойственность продольных перемычек между ганглиями соседних сегментов свидетельствует о парном происхождении брюшной нервной цепочки[32]. Как и у большинства членистоногих, у ракообразных проявляется тенденция к олигомеризации (слиянию) ганглиев из разных сегментов, что отличает брюшную нервную цепочку членистоногих от таковой у кольчатых червей[33]. Так, у речного рака, тело которого состоит из 18 сегментов, имеется только 12 нервных узлов[34].
Головной мозг ракообразных представлен парными долями протоцеребрума (иннервация акрона и глаз) с грибовидными телами и дейтоцеребрума (иннервация антеннул). Обычно с головным мозгом сливаются смещающиеся вперёд ганглии сегмента, несущего вторую пару антенн. В этом случае обособлен третий отдел — тритоцеребрум (иннервация антенн), у остальных ракообразных антенны управляются окологлоточным кольцом[33][35].
У ракообразных имеется хорошо развитая симпатическая нервная система, в основном иннервирующая кишечник. Она состоит из церебрального отдела и непарного симпатического нерва, по ходу которого располагаются несколько ганглиев[35].
Нервная система ракообразных тесно связана с эндокринной. В состав ганглиев у раков входят нейросекреторные клетки, выделяющие гормоны, которые поступают в гемолимфу. Эти гормоны оказывают влияние на обменные процессы, линьку и развитие. Нейросекреторные клетки располагаются в различных частях протоцеребрума, тритоцеребрума и ганглиев брюшной нервной цепочки[35]. У некоторых ракообразных гормоны из нейросекреторных клеток зрительных нервов поступают в особую синусовую железу и оттуда в гемолимфу. Они ответственны за описанный выше механизм изменения окраски тела[33].
Органы чувств
- Органы зрения


Почти у всех ракообразных хорошо развиты глаза: простые или фасеточные (сложные), глаза отсутствуют лишь у глубоководных, сидячих и паразитических видов. Некоторые ракообразные (циклопы) имеют только простые глазки, большинство же высших раков имеет только сложные глаза, а у карпоедов имеются глаза обоих типов[36].
Простой глазок представляет собой пигментный бокал, внутрь которого обращены зрительные клетки. Он покрыт прозрачной кутикулой, образующей хрусталик. Свет сначала проходит через хрусталик, зрительные клетки и только потом — на их светочувствительные концы. Такие глазки называют инвертированными (то есть обращёнными). Простые глазки собраны по 2—4 и образуют непарный науплиусов (науплиальный) глаз, характерный для личинки ракообразных — науплиуса[37]. У взрослых науплиусов глаз расположен между основаниями антенн[38].
Фасеточные глаза состоят из простых глазков — омматидиев. Каждый простой глазок представляет собой конусовидный бокал, ограниченный пигментными клетками и сверху покрытый шестигранной роговицей. Светопреломляющую часть омматидия составляют клетки хрустального конуса, а светочувствительную — ретинальные клетки, в месте соприкосновения которых образуется светочувствительная палочка — рабдом. У ракообразных с фасеточными глазами имеет место мозаичное зрение, то есть общее зрительное восприятие складывается из частей, воспринимаемых отдельными омматидиями[39]. Сложные глаза нередко сидят на особых подвижных выростах головы — стебельках[40].
У некоторых раков для запуска описанного выше механизма изменения окраски тела необходимы зрительные восприятия определённых световых раздражений[40].
- Органы равновесия
У некоторых ракообразных есть органы равновесия — статоцисты. У речного рака они находятся у основания антеннул. В период линьки меняется выстилка статоциста, и животное теряет координацию движения[39]. Статоцисты характерны для десятиногих и некоторых других высших раков[38].
- Другие органы чувств
Органами осязания и обоняния у ракообразных являются многочисленные сенсиллы и осязательные волоски, в основном расположенные на антеннах, конечностях и вилочке[39]. Чувство осязания приурочено только к тем участкам покровов, на которых располагаются чувствительные волоски. У основания таких волосков под гиподермальным эпителием находятся биполярные нейроны. Волоски с особенно проницаемой кутикулой, локализованные на антеннах, являются органами обоняния[35].
Половая система
Ракообразные — в подавляющем большинстве раздельнополые животные и размножаются половым путём[41]. Известны и случаи гермафродитизма — у некоторых усоногих, ремипедий[42], цефалокарид[43]. Нередко выражен половой диморфизм: так, у некоторых паразитических ракообразных самцы в несколько раз мельче самок[39]. Некоторые ракообразные в течение жизни способны сменять пол[43]. Кроме того, среди ракообразных широко распространён партеногенез[41]. Он имеет место среди многих жаброногих, некоторых ракушковых, ветвистоусых (дафнии)[44], равноногих раков, а также среди некоторых высших ракообразных, например, у Procambarus fallax subsp. virginalis.

Иногда у самцов антенны или антеннулы выполняют роль хватательных органов, а у речного рака 1—2 пары брюшных ног функционируют как копулятивные органы. Гонады у примитивных форм, половые протоки и отверстия парные. Гораздо чаще гонады полностью или частично срастаются. Стенки яйцеводов выделяют вокруг яиц плотную скорлупу. В некоторых случаях у самок имеются семяприёмники. В этом случае оплодотворение происходит, когда самка откладывает яйца и опрыскивает их спермой из отверстий семяприёмников. У некоторых раков имеет место сперматофорное оплодотворение; при спаривании самцы этих видов приклеивают сперматофоры к телу самки или вводят их в её половое отверстие[45][30].
У ракообразных широко различаются форма и размеры сперматозоидов. Так, у некоторых мелких ракушковых рачков длина сперматозоидов составляет 6 мм, что в 10 раз длиннее самого животного. У галатеи (Galathea) и высших раков сперматозоид похож на песочные часы. При оплодотворении сперматозоид прикрепляется к яйцеклетке отростками, затем хвостовая часть сперматозоида, впитывая влагу, разбухает и взрывается, а головной конец с ядром вонзается в яйцо[46].
Для большинства раков характерна забота о потомстве, хотя некоторые из них просто выбрасывают яйца в толщу воды. Часто самки вынашивают яйца приклеенными к половым отверстиям в виде яйцевых мешков (характерны для веслоногих) или длинных нитей. Десятиногие приклеивают яйца к конечностям брюшка. У перакарид, щитней, жаброногов и многих равноногих из карапакса и грудных ножек образуется выводковая сумка (марсупиум)[41]. Большая часть тонкопанцирных и крилевых рачков вынашивают яйца между грудными ножками[41]. Самки карпоедов не вынашивают яйца, но откладывают их рядами на камни и другие предметы[47].
Плодовитость раков различна[30].
Яйца некоторых раков (щитней и жаброногов) обладают большой стойкостью: они легко переносят высыхание, промерзание и переносятся ветром[48].
Жизненный цикл
Эмбриональное развитие
Характер дробления ракообразных зависит от количества желтка в яйцах. Когда желтка в яйце мало (например, некоторые веслоногие), дробление идёт подобно дроблению кольчатых червей: оно полное, неравномерное, детерминированное, с телобластической закладкой мезодермы (то есть из клетки — телобласта)[49].
У большинства раков яйца богаты желтком, и дробление становится частичным и поверхностным. В ходе нескольких делений ядра без деления клетки образуются дочерние ядра, уходящие к периферии и располагающиеся там в один слой (поэтому дробление ракообразных называется поверхностным). Далее вокруг каждого ядра обособляется участок цитоплазмы, и образуется небольшая клетка; центральная же масса желтка остаётся неразделённой. Эта стадия аналогична бластуле с бластоцелем, заполненным желтком. Потом часть клеток бластулы на будущей брюшной стороне уходит под наружный слой, образуя многоклеточную пластинку — зародышевую полоску. Её наружный слой образован эктодермой, более глубокие — мезодерму, самый глубокий, прилегающий к желтку, — энтодерму[50].
Дальнейшее развитие зародыша происходит в основном за счёт зародышевой полоски. Она начинает сегментироваться, и из самого переднего и мощного её участка появляются парные головные ганглии, за счёт которых возникают сложные глаза. Позади неё закладываются зачатки акрона, антеннального и мандибулярного сегментов. Иногда мезодерма закладывается в виде парных целомических мешков, как у кольчатых червей, которые впоследствии разрушаются: их клетки идут на построение мезодермальных органов (мышц, сердца и др.), а полости сливаются с остатками первичной полости тела. Так формируется миксоцель, или смешанная полость тела. В некоторых случаях мезодерма теряет отчётливую сегментацию, а выраженный целом вовсе не формируется[51].
Постэмбриональное развитие
Личинки ракообразных

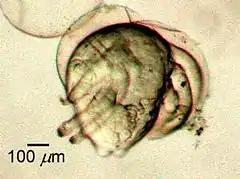

Постэмбриональное развитие большинства ракообразных происходит с метаморфозом. Как правило, из яйца выходит планктонная личинка — науплиус, эта личинка наиболее характерна для ракообразных. Строение науплиуса характеризуется следующими особенностями. Тело состоит из акрона, двух сегментов тела и анальной лопасти, имеются одноветвистые антеннулы и 2 пары двуветвистых плавательных ног, которые голомологичны антеннам и мандибулам взрослых раков. Науплиус имеет кишечник, почки (чаще антеннальные), головные ганглии и уже упоминавшийся непарный науплиальный глаз на головной лопасти. Перед анальной лопастью находится зона роста, где закладываются новые сегменты. За стадией науплиуса следует стадия метанауплиуса, имеющего все головные сегменты с конечностями и передние грудные сегменты с ногочелюстями. Личинки претерпевают несколько линек, в ходе которых их внешнее и внутреннее строение достигают уровня развития, характерного для взрослых особей[52].
У высших ракообразных за стадией метанауплиуса следует особая личиночная стадия — зоеа (такое название личинка получила, когда учёные считали её отдельным видом[53]). У этой личинки развиты головные и переднегрудные конечности, имеются зачатки остальных грудных ног, сформированное брюшко с последней парой ног. Кроме того, у зоеа имеются фасеточные глаза. Далее зоеа развивается в мизидную личинку с сформированными грудными ножками и зачатками всех брюшных конечностей. После этого мизидная личинка линяет и преобразуется во взрослое животное[52].
От описанного выше жизненного цикла у некоторых высших раков имеются отличия. Так, у многих крабов из яйца сразу выходит зоеа, а у речного рака развитие прямое: из яйца появляется молодой рачок с полным составом сегментов и конечностей, далее он растёт и линяет, превращаясь во взрослую особь[54].
Наконец, у различных групп ракообразных могут иметься особенные личиночные стадии.
Линька
Линька у ракообразных наиболее хорошо изучена на примере высших раков. Она сопровождается как морфологическими, так и физиологическими изменениями[55].
Перед линькой в тканях и гемолимфе животного накапливается ряд органических (липиды, белки, витамины, углеводы и др.) и минеральных соединений. Частично они поступают из старой кутикулы. Усиливается потребление кислорода, возрастает интенсивность обменных процессов[55].
Одновременно с этим клетки гиподермы начинают выделять новую кутикулу за счёт веществ из гемолимфы и тканей. Новая кутикула постепенно утолщается, сохраняя, однако, гибкость и эластичность. Наконец старый кутикулярный покров лопается, животное выбирается из него, оставляя пустой чехол — экзувий. Перелинявший рак быстро увеличивается в размерах, но не за счёт разрастания тканей, а накопления в них воды. За счёт деления клеток объём тканей увеличивается только между линьками. Через некоторое время после сбрасывания экзувия в новой кутикуле откладываются минеральные соли, и она быстро твердеет[55].
Процесс линьки регулируется гормональной системой. Важную роль в ней играют нейросекреторные клетки, связанные с упоминавшейся выше синусовой железой, и небольшой эндокринной головной железой. Её гормоны запускают и ускоряют линьку, а в нейросекреторных клетках глазных стебельков вырабатываются гормоны, подавляющие её деятельность, то есть препятствующие наступлению линьки. Особо велико их содержание в период после линьки и между линьками, потом же деятельность головной железы активизируется и начинается подготовка к новой линьке. Кроме вышеописанных, в регуляции линьки принимают участие и другие гормоны[56].
Другие особенности жизненных циклов
Для некоторых ракообразных, например, дафний, характерны сложные жизненные циклы с чередованием партеногенетического и полового размножения. Кроме того, у поколений дафний, живущих в разное время года, происходят сезонные изменения, выражающиеся в изменении формы головы, длины рострума, шипов и др[57].
Экология и образ жизни
Распространение
В настоящее время считается, что исходным морфоадаптивным типом ракообразных были мелкие пелаго-бентосные формы, ведущие плавающий образ жизни. От них произошли группы, специализировавшиеся к планктонному, нектонному и бентосному образу жизни. Часть групп приспособилась к паразитическому образу жизни, некоторые вышли на сушу[58].
Паразитические формы имеются во многих группах ракообразных: веслоногие, усоногие раки, паразитический отряд карпоеды, представители которого живут на коже рыб[59]. При этом они претерпевают упрощения в организации в разной степени: веслоногий рачок Ergasilus внешне напоминает циклопа, Lamproglena ещё сохранила частичную сегментацию, а паразит рыб Lernaeocera branchialis и усоногие рачки — паразиты десятиногих Sacculina и Peltogaster характеризуются столь глубоким упрощением организации, что их систематическую принадлежность удалось установить, лишь пронаблюдав историю развития[60]. Ещё один паразитический отряд ракообразных — мешкогрудые — паразитируют на коралловых полипах и иглокожих[61]. Существуют и паразитические высшие раки, например, некоторые равноногие рачки. Среди них есть временные (Aega) и постоянные (Cymothoa, Livoneca) эктопаразиты рыб[62]. Как видно из примеров, среди ракообразных есть как эктопаразиты, так и эндопаразиты.
В морях и океанах ракообразные распространены настолько же широко, как насекомые на суше. Ракообразные многообразны в пресных водоёмах, а некоторые жаброноги встречаются во временных лужах, остающихся после таяния снега. Другой жаброногий рачок — Artemia salina — обитает в осолоненных водоёмах в степях и полупустынях: в лиманах, солёных озёрах[63].
Питание
Большинство планктонных ракообразных питается бактериями, а также одноклеточными организмами, детритом. Живущие на дне питаются частицами органических веществ, растениями или животными. Бокоплавы поедают трупы животных, способствуя тем самым очищению водоёмов[14].
Был проведён ряд исследований пищевого поведения краба Portunus pelagicus, в которых исследовались реакции животного на конкретные пищевые вещества, а также сравнивались с реакциями на естественную пищу (рыбу, моллюсков). В результате было установлено, что реакция ракообразного на некоторые аминокислоты и сахариды была такой же, как на естественную пищу, причём реакции на аминокислоты и сахариды были очень похожими. Особо сильный ответ наблюдался на аланин, бетан, серин, галактозу и глюкозу. Эти данные могут быть полезными для разведения крабов в аквакультуре[64].
Для щитней характерен древний тип питания, имевший место также у трилобитов: они питаются кусочками детрита и мелкими донными животными, которые захватываются жевательными отростками всех ножек и передаются затем по брюшному желобку ко рту[65].
Практическое значение
Ракообразные — важный объект промысла, включая добычу креветок, крабов, лангустов, лангустинов, раков, омаров (лобстеров), разнообразных балянусов, включая морскую уточку (или персебеса), который является самым дорогим из деликатесных ракообразных.
На рыбоводных заводах разводят рачков в качестве корма для рыб. Кроме того, мелкие рачки являются одним из основных видов пищи многих промысловых рыб. Важна роль ракообразных в биологической очистке вод, они представляют одну из самых многочисленных групп биофильтраторов и детритофагов. С другой стороны, некоторые ракообразные могут быть переносчиками разных инфекций. Сидячие формы ракообразных прикрепляются к основанию судов и замедляют их скорость. Некоторые ракообразные ведут паразитический образ жизни (карповая вошь).
Классификация

В настоящее время известно более 73 000 видов ракообразных (включая более 5 тыс. ископаемых видов), объединяемых в 1003 семейства, более 9500 родов (Zhang, 2013)[3], 42 отряда и 6 классов[4]:
- Класс Branchiopoda — Жаброногие
- Подкласс Phyllopoda — Листоногие
- Подкласс Sarsostraca — Сарсостраки
- Класс Cephalocarida — Цефалокариды
- Класс Malacostraca — Высшие раки
- Подкласс Eumalacostraca — Эумалакостраки
- Подкласс Hoplocarida — Гоплокариды
- Подкласс Phyllocarida — Филлокариды
- Класс Maxillopoda — Максиллоподы
- Подкласс Branchiura — Карповые вши
- Подкласс Copepoda — Веслоногие
- Подкласс Mystacocarida — Мистакокариды
- Подкласс Pentastomida — Пятиустки
- Подкласс Tantulocarida — Тантулокариды
- Подкласс Thecostraca — Текостраки
- Класс Ostracoda — Ракушковые
- Подкласс Myodocopa — Миодокоповые
- Подкласс Podocopa — Подокоповые
- Класс Remipedia — Ремипедии
По последним данным, от ракообразных произошли насекомые — класс Hexapoda, которые являются сестринской группой жаброногих. В случае принятия этой концепции (концепции Pancrustacea или Tetraconata, см., например[66][67]) приходится изменить таксономическое положение[уточнить] ракообразных (например, для них уже не является общим признаком наличие двух пар антенн). В противном случае ракообразные оказываются парафилетическим таксоном.
Альтернативная классификация
Представленную выше классификацию разделяют не все систематики. Сайт World Register of Marine Species использует другую, отличающуюся прежде всего расформированием мусорного класса максиллопод и выделением двух надклассов. Классификация до подклассов включительно[2]:
- Класс Branchiopoda — Жаброногие
- Подкласс Calmanostraca
- Подкласс Diplostraca
- Подкласс Sarsostraca — Сарсостраки
- Класс Cephalocarida — Цефалокариды
- Класс Remipedia — Ремипедии
- Надкласс Multicrustacea
- Класс Hexanauplia
- Подкласс Copepoda — Веслоногие
- Подкласс Tantulocarida — Тантулокариды
- Подкласс Thecostraca — Текостраки
- Класс Malacostraca — Высшие раки
- Подкласс Eumalacostraca — Эумалакостраки
- Подкласс Hoplocarida — Гоплокариды
- Подкласс Phyllocarida — Филлокариды
- Класс Hexanauplia
- Надкласс Oligostraca
- Подкласс Mystacocarida — Мистакокариды
- Класс Ichthyostraca
- Подкласс Branchiura — Карповые вши
- Подкласс Pentastomida — Пятиустки
- Класс Ostracoda — Ракушковые
- † Подкласс Metacopa
- Подкласс Myodocopa — Миодокоповые
- Подкласс Palaeocopa
- Подкласс Platycopa
- Подкласс Podocopa — Подокоповые
См. также
Примечания
- Ракообразные / Чесунов А. В. // Пустырник — Румчерод. — М. : Большая российская энциклопедия, 2015. — С. 201—202. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 28). — ISBN 978-5-85270-365-1.
- Подтип Crustacea (англ.) в Мировом реестре морских видов (World Register of Marine Species). (Дата обращения: 2 февраля 2019).
- Zhang Z.-Q. Phylum Athropoda // Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013) (англ.) // Zootaxa : монография; журнал / Zhang Z.-Q. (Ed.). — Auckland, New Zealand: Magnolia Press, 2013. — Vol. 3703, no. 1. — P. 17—26. — ISBN 978-1-77557-248-0 (paperback), ISBN 978-1-77557-249-7 (online edition). — ISSN 1175-5326. — doi:10.11646/zootaxa.3703.1.6. (Дата обращения: 7 марта 2015)
- Martin J. W., Davis G. E. An Updated Classification of the Recent Crustacea. (англ.) — Los Angeles: Natural History Museum of Los Angeles County, 2001. — 132 p. (PDF)
- Шарова, 2002, с. 348.
- Догель, 1981, с. 293.
- Шарова, 2002, с. 363.
- Корнев П. Н. Первое нахождение представителей подкласса Tantulocarida в Белом море // Зоология беспозвоночных : журнал. — 2004. — Т. 1, № 1. (PDF)
- Корнев П. Н., Чесунов А. В. Тантулокариды — микроскопические обитатели Белого моря // Природа : журнал. — Наука, 2005. — № 2. (PDF)
- McClain C. R., Boyer A. G. Biodiversity and body size are linked across metazoans (англ.) // Proceedings of the Royal Society B: Biological Sciences : журнал. — 2009. — Vol. 296, no. 1665. — P. 2209—2215. — doi:10.1098/rspb.2009.0245. — PMID 19324730. (Дата обращения: 2 марта 2015)
- Crustacea — National History Museum
- Шарова, 2002, с. 349.
- Иллюстрация из книги Вилли Георга Кюкенталя: Leitfaden für das Zoologische Praktikum, 1912. (нем.)
- Ракообразные // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- Ракообразные — статья из Биологического энциклопедического словаря
- Догель, 1981, с. 295.
- Догель, 1981, с. 296.
- Шарова, 2002, с. 349—351.
- Шарова, 2002, с. 351.
- Шарова, 2002, с. 352.
- Догель, 1981, с. 296—297.
- Догель, 1981, с. 297.
- Догель, 1981, с. 298.
- Шарова, 2002, с. 352—353.
- Догель, 1981, с. 299.
- Шарова, 2002, с. 353.
- Догель, 1981, с. 300.
- Urich K. Respiratory pigments // Comparative Animal Biochemistry / K. Urich; Translated from the German by P. J. King. — Berlin—Heidelberg: Springer-Verlag, 1994. — P. 249—287. — 782 p. — ISBN 3-540-57420-4. (англ.) (Дата обращения: 31 января 2017)
- Шарова, 2002, с. 354.
- Догель, 1981, с. 306.
- Шарова, 2002, с. 355—356.
- Догель, 1981, с. 301.
- Шарова, 2002, с. 356.
- Догель, 1981, с. 302.
- Догель, 1981, с. 303.
- Шарова, 2002, с. 357.
- Шарова, 2002, с. 357—358.
- Догель, 1981, с. 304.
- Шарова, 2002, с. 358.
- Догель, 1981, с. 305.
- Crustacean (arthropod) // Encyclopædia Britannica.
- G. L. Pesce. Remipedia Yager, 1981.
- Aiken D. E., Tunnicliffe V., Shih C. T., Delorme L. D. Crustacean. The Canadian Encyclopedia. Дата обращения: 11 декабря 2009.
- Шарова, 2002, с. 364.
- Шарова, 2002, с. 358—359.
- Шарова, 2002, с. 359—360.
- Alan P. Covich & James H. Thorp. Introduction to the Subphylum Crustacea // Ecology and classification of North American freshwater invertebrates (англ.) / James H. Thorp & Alan P. Covich. — 2nd. — Academic Press, 2001. — P. 777—798. — ISBN 978-0-12-690647-9.
- Догель, 1981, с. 313.
- Догель, 1981, с. 307.
- Догель, 1981, с. 307—308.
- Догель, 1981, с. 308—309.
- Шарова, 2002, с. 360.
- William Thomas Calman Crab // Encyclopædia Britannica Eleventh Edition (англ.). — 1911. Архивированная копия (недоступная ссылка). Дата обращения: 11 ноября 2013. Архивировано 29 мая 2013 года.
- Шарова, 2002, с. 360—361.
- Догель, 1981, с. 310.
- Догель, 1981, с. 311.
- Догель, 1981, с. 314.
- Шарова, 2002, с. 383.
- Догель, 1981, с. 317.
- Догель, 1981, с. 317—319.
- Догель, 1981, с. 320.
- Догель, 1981, с. 326.
- Догель, 1981, с. 312.
- Archdale M. V., Anraku K. Feeding behavior in scyphozoa, crustacea and cephalopoda (англ.) // Chemical Senses : журнал. — 2005. — Vol. 30, no. Suppl. 1. — P. 303—304. — ISSN 0379-864X. — doi:10.1093/chemse/bjh235. — PMID 15738170.
- Догель, 1981, с. 312—313.
- Shultz J. W., Regier J. C. Phylogenetic analysis of arthropods using two nuclear protein-encoding genes supports a crustacean + hexapod clade (англ.) // Proceedings. Biological sciences. — L.: The Royal Society, 2000. — Vol. 267, no. 1447 (22 May). — P. 1011—1019. — doi:10.1098/rspb.2000.1104. — PMID 10874751. (PDF)
- Элементы — новости науки: Новые данные позволили уточнить родословную животного царства
Литература
- Догель В. А. Зоология беспозвоночных. — 7-е изд.. — М.: Высшая школа, 1981. — 614 с.
- Пименова И. Н., Пименов А. В. Зоология беспозвоночных. Теория. Задания. Ответы. — Саратов: Лицей, 2005. — 288 с. — ISBN 5-8053-0308-6.
- Шарова И. Х. Зоология беспозвоночных. — М.: Владос, 2002. — 592 с. — ISBN 5-691-00332-1.
- Шевяков В. Т. Ракообразные // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- Carbonnier P. L'écrevisse: mœurs — reproduction — éducation. — Paris, 1869. (фр.)
- Huys R., Boxshall G. A. Copepod Evolution. — The Ray Society, 1991. — 468 p. (англ.)
- McLaughlin P. A. Comparative Morphology of Recent Crustacea. — San Francisco: W. H. Freeman and Company, 1979. — 177 p. (англ.)
Ссылки
- «Ракообразные» — статья в Энциклопедии «Кругосвет».
- «Ракообразные» — статья А. В. Рыбакова на сайте Дальневосточного геологического института ДВО РАН.
- Lowry J. K. (1999 onwards). Crustacea, the Higher Taxa: Description, Identification, and Information Retrieval. Version: 2 October 1999. (англ.)
.svg.png.webp) Ракообразные. Учебный фильм для средней советской школы.
Ракообразные. Учебный фильм для средней советской школы.
| Таксономия | |
|---|---|
| В библиографических каталогах |
Современные членистоногие | |
|---|---|
| |
| Ракообразные (Crustacea) |
|
| Хелицеровые (Chelicerata) |
|
| Многоножки (Myriapoda) |
|
| Шестиногие (Hexapoda) |
|
