Экологическая ниша
Экологи́ческая ни́ша — место, занимаемое видом в биоценозе, включающее комплекс его биоценотических связей и требований к факторам среды.[1] Термин введён в 1914 году Дж. Гриннеллом и в 1927 году Чарльзом Элтоном[2]. В настоящее время определение Гриннелла принято называть пространственной нишей (по смыслу термин ближе понятию местообитание), а определение Элтона называют трофической нишей (экологическая ниша представляет собой сумму факторов существования данного вида, основным из которых является его место в пищевой цепочке)[3]. В настоящее время доминирует модель гиперобъёма Дж. Э. Хатчинсона[4][5]. Модель представлена как n-мерный куб, на осях которого отложены экологические факторы. По каждому фактору у вида есть диапазон, в котором он может существовать (экологическая валентность). Если провести проекции от крайних точек диапазонов каждой оси факторов, мы получим n-мерную фигуру, где n — количество значимых для вида экологических факторов. Модель в основном умозрительна, но позволяет получить хорошее представление об экологической нише[6]. По Хатчинсону[7] экологическая ниша может быть:
- фундаментальной — определяемой сочетанием условий и ресурсов, позволяющим виду поддерживать жизнеспособную популяцию;
- реализованной — свойства которой обусловлены конкурирующими видами.

Допущения модели:
- Реакция на один фактор не зависит от воздействия другого фактора;
- Независимость факторов друг от друга;
- Пространство внутри ниши однородное с одинаковой степенью благоприятности.
Это различие подчёркивает, что межвидовая конкуренция приводит к снижению плодовитости и жизнеспособности и что в фундаментальной экологической нише может быть такая часть, занимая которую вид в результате межвидовой конкуренции не в состоянии больше жить и успешно размножаться. Эта часть фундаментальной ниши вида отсутствует в его реализованной нише[8]. Таким образом, реализованная ниша всегда входит в состав фундаментальной или равна ей.
Принцип конкурентного исключения
Суть принципа конкурентного исключения, также известного как принцип Гаузе, состоит в том, что каждый вид имеет свою собственную экологическую нишу[9]. Никакие два разных вида не могут занять одну и ту же экологическую нишу. Сформулированный таким образом принцип Гаузе подвергался критике. Например, одним из известных противоречий этому принципу является «планктонный парадокс». Все виды живых организмов, относящихся к планктону, живут на очень ограниченном пространстве и потребляют ресурсы одного рода (главным образом солнечную энергию и морские минеральные соединения). Современный подход к проблеме разделения экологической ниши несколькими видами указывает, что в одних случаях два вида могут разделять одну экологическую нишу, а в других такое совмещение приводит к вымиранию одного из видов.
Вообще, если речь идёт о конкуренции за определённый ресурс, становление биоценозов связано с расхождением экологических ниш и уменьшением уровня межвидовой конкуренции[10][11]:стр.423. При таком варианте правило конкурентного исключения подразумевает пространственное (иногда функциональное) разобщение видов в биоценозе. Абсолютное вытеснение, при подробном изучении экосистем, зафиксировать почти невозможно[11]:стр.423
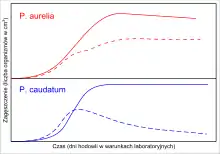
Гаузе сформулировал принцип конкурентного исключения работая с инфузориями Paramecium caudatum, P. aurelia, P. bursaria. Все три вида хорошо росли в монокультуре, достигая в пробирках с жидкой средой стабильных значений предельных плотностей популяции. Пищей инфузориям служили бактериальные или дрожжевые клетки, растущие на регулярно добавляемой овсяной муке. Однако, когда он фактически смоделировал экологическую нишу, совместно выращивая P. caudatum и P. aurelia, было показано что P. aurelia вытеснила P. caudatum. При совместном выращивании Р. caudatum и Р. bursaria сосуществуют, но на более низком уровне плотности, чем в монокультуре. Как выяснилось, они были пространственно разобщены в пробирке Р.bursaria — на дне пробирке и питалась дрожжами, тогда как Р. caudatum — наверху и питается бактериями.
С тех пор принцип конкурентного исключения, гласящий, что «полные конкуренты не могут существовать бесконечно», стал одним из главных догматов теоретической экологии. Таким образом, если два вида сосуществуют, то между ними должно быть какое-то экологическое различие, а это означает, что каждый из них занимает свою особую нишу.
Конкурируя с более приспособленным видом, менее приспособленный конкурент утрачивает свою реализованную нишу. Таким образом, выход из конкуренции достигается расхождением требований к среде, изменению образа жизни или, другими словами, является разграничением экологических ниш видов. В этом случае они приобретают способность сосуществовать в одном биоценозе. Так, в мангровых зарослях побережья Южной Флориды обитают самые разные цапли и нередко на одной и той же отмели кормятся рыбой до девяти разных видов. При этом они практически не мешают друг другу, так как в их поведении — в том, какие охотничьи участки они предпочитают и как ловят рыбу, — выработались приспособления, позволяющие им занимать различные ниши в пределах одной и той же отмели.
Однако, желая выяснить, работает ли принцип конкурентного исключения в какой-либо конкретной ситуации, мы можем столкнуться с очень серьёзной методологической проблемой. Рассмотрим, например, случай с саламандрами из работы Хейрстона. В этом примере — два вида сухопутных саламандр Plethodon glutinosus и Plethodon jordani, обитающих в южной части Аппалачских гор в США. Обычно P. jordani встречается на больших высотах, чем P. glutinosus, но в некоторых районах зоны их обитания перекрывались. Важным моментом является то, что неблагоприятное воздействие со стороны другого вида исходно испытывали особи обоих видов. После удаления одного из видов у оставшегося наблюдалось значительное увеличение численности и (или) плодовитости и (или) выживаемости. Из этого следует, что на контрольных площадках и в других местах совместного обитания эти виды обычно конкурировали друг с другом, но все же существовали.
Два вида конкурируют и сосуществуют; в соответствии с принципом конкурентного исключения можно предполагать, что это происходит вследствие разделения ниш. Это вполне правдоподобное предположение, но до тех пор, пока такое разделение не обнаружено или не подтверждено, что оно снимает напряжение межвидовой конкуренции, оно остаётся не более чем предположением. Таким образом, когда мы наблюдаем сосуществование двух конкурентов, часто бывает трудно установить, что их ниши разделены, и невозможно доказать обратное. Если экологу не удаётся обнаружить разделение ниш, то это может просто означать, что он искал его не там или не так. В XX веке принцип конкурентного исключения получил широкое признание из-за многочисленности подтверждающих его фактов; наличия некоторых теоретические предпосылок, свидетельствующих в его пользу, например, модели Лотки — Вольтерры (но сама эта модель в большой степени идеальна и почти неприменима для ненарушенных экосистем[11]:стр.355).
Однако при этом всегда будут оставаться такие случаи, в которых его невозможно проверить. Кроме того, есть случаи, когда принцип Гаузе просто не применим. Например, «планктонный парадокс». В действительности равновесие между конкурирующими видами может неоднократно нарушаться и преимущество будет переходить от одного вида к другому; следовательно, сосуществование возможно просто благодаря изменению условий среды. Такой аргумент использовал Хатчинсон (Hutchinson, 1961) для того, чтобы объяснить «планктонный парадокс». Парадокс этот состоит в том, что многочисленные виды планктонных организмов часто сосуществуют в простой среде, где, по-видимому, мало возможностей для разделения ниш. Хатчинсон предположил, что среда, хотя она и очень проста, постоянно претерпевает различные изменения, в частности сезонные. В любой отдельный промежуток времени условия среды могут способствовать вытеснению определённого вида, однако эти условия меняются, и ещё до того, как данный вид окажется окончательно вытесненным, они могут сложиться благоприятно для его существования. Другими словами, исход конкурентных взаимодействий в равновесном состоянии может не играть решающей роли, если условия среды обычно изменяются задолго до того, как достигается равновесие. А поскольку любая среда изменчива, равновесие между конкурентами должно постоянно сдвигаться и сосуществование нередко будет наблюдаться при таком разделении ниш, при котором в стабильных условиях происходило бы исключение одного из видов. Было высказано множество гипотез, дабы разрешить указанный парадокс:
- неравновесная модель Ф. Н. Стюарта и Б. Р. Левина, подразумевающая ситуации, когда виды не пересекаются фактически за счёт расхождения сезонов жизнедеятельности;
- неравновесная модель Ю. А. Домбровского, в которой высказывалась идея о наличии в планктоне неперемешиваемых «пятен»;
- равновесная модель Р. Петерсена;
Кроме того, существует проблема адекватного определения реальной экологической ниши, то есть исследователю может показаться, что виды пересекаются в пространстве факторов, а на самом деле виды могут сосуществовать за счёт неучтённых факторов. Очень показательным в этой связи является работа М. Джилпина «Едят ли зайцы рысей?» при исследовании статистических данных заготовок пушнины в Канаде[12].
Закон константности В. И. Вернадского
Количество живого вещества природы (для данного геологического периода) есть константа.
Согласно этой гипотезе, любое изменение количества живого вещества в одном из регионов биосферы должно быть компенсировано в каком-либо другом регионе. Правда, в соответствии с постулатами видового обеднения, высокоразвитые виды и экосистемы чаще всего будут заменяться эволюционно объектами более низкого уровня. Кроме того, будет происходить процесс рудерализации видового состава экосистем, и «полезные» для человека виды будут замещаться менее полезными, нейтральными или даже вредными.
Следствием этого закона есть правило обязательного заполнения экологических ниш. (Розенберг и др, 1999)
Правило обязательного заполнения экологической ниши
Экологическая ниша не может быть пустой. Если ниша пустеет в результате вымирания какого-то вида, то она тут же заполняется другим видом.
Среда обитания обычно состоит из отдельных участков («пятен») с благоприятными и неблагоприятными условиями; эти пятна нередко доступны лишь временно, и возникают они непредсказуемо как во времени, так и в пространстве.
Свободные участки или «бреши» в местообитаниях возникают непредсказуемо во многих биотопах. Пожары или оползни могут приводить к образованию пустошей в лесах; шторм может оголить открытый участок морского берега, а прожорливые хищники где угодно могут истребить потенциальных жертв. Эти освободившиеся участки неизменно заселяются вновь. Однако самыми первыми поселенцами не обязательно будут те виды, которые в течение длительного времени способны успешно конкурировать с другими видами и вытеснять их. Поэтому сосуществование преходящих и конкурентоспособных видов возможно так долго, как с подходящей частотой появляются незаселённые участки. Преходящий вид обычно первым заселяет свободный участок, осваивает его и размножается. Более конкурентоспособный вид заселяет эти участки медленно, но если заселение началось, то со временем он побеждает преходящий вид и размножается. (Бигон и др., 1989)
Экологическая ниша человека
Человек как биологический вид занимает свою собственную экологическую нишу. Человек может обитать в тропиках и субтропиках, на высотах до 3—3,5 км над уровнем моря. Реально в настоящее время человек живёт в значительно больших пространствах. Человек расширил свободную экологическую нишу благодаря использованию различных приспособлений: жилища, одежды, огня и пр.
См. также
Примечания
- Pocheville, A., 2015. The Ecological Niche: History and Recent Controversies, in: Heams, T., Huneman, P., Lecointre, G., Silberstein, M. (Eds.), Handbook of Evolutionary Thinking in the Sciences. Springer, Dordrecht, pp. 547—586. doi:10.13140/RG.2.1.3205.8405
- Экология окружающей среды (недоступная ссылка). Дата обращения: 13 ноября 2010. Архивировано 1 декабря 2009 года.
- Одум Ю. Экология: В 2-х т. — Пер. с англ. — М.:. Мир, 1986. Т.1. — 328 с. Т.2. — 376 с.
- Hutchinson G.E. Concluding remarks // Cold Spring Harbor Symposia on Quantitative Biology. 1957. № 22. P. 415—427. Reprinted in 1991: Classics in Theoretical Biology. Bull. of Math. Biol. 53: 193—213.
- Гиляров А. М. Популяционная экология: Учебное пособие. — М.: Изд-во МГУ, 1990—191 с.
- Джиллер П. Структура сообществ и экологическая ниша. — М.: Мир, 1988. — 184 с.
- В.Грант. Эволюция организмов. М.: Мир. Глава 22 (недоступная ссылка). Дата обращения: 29 октября 2010. Архивировано 9 мая 2011 года.
- Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества. Том 1. — М.: Мир, 1989. — 667 с.
- Розенберг Г. С., Мозговой Д. П., Гелашвили Д. Б. Экология. Элементы теоретических конструкций современной экологии. (Учебное пособие).-Самара: СНЦ РАН, 1999—396 с.
- Конкуренция // Биологический энциклопедический словарь / глав. ред. М. С. Гиляров. — М.: Советская энциклопедия, 1986. — С. 277.
- Шилов И. А. Экология. — М.: Высшая школа, 1997. — 512 с.
- Gilpin M.E. Do Hares Eat Lynx? // The American Naturalist. 1973. V.107, №.957. P. 727—730.
| История Земли |  | |
|---|---|---|
| Физические свойства Земли | ||
| Оболочки Земли | ||
| География и геология | ||
| Окружающая среда | ||
| См. также | ||
| ||