Сигнальный путь Wnt
Сигнальный путь Wnt — один из внутриклеточных сигнальных путей животных, регулирующий эмбриогенез, дифференцировку клеток и развитие злокачественных опухолей[1][2].
Открытие
Название Wnt — это комбинация Wg (англ. wingless) и Int[3] (произносится как уинт). Ген дрозофилы wingless первоначально был идентифицирован по рецессивной мутации, подавляющей у мушки развитие крыльев[4][5][6]. Гомологичный ген позвоночных Int вначале изучали в связи с присутствием в его локусе нескольких мест интеграции генома вируса рака молочных желез мыши[7]. Исследование функций этих двух генов привело к открытию целого класса лигандов, регулирующих эмбриональное развитие животных.
Белки Wnt
| Виды | Wnt белки |
|---|---|
| Homo sapiens | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt9A, Wnt9B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Домовая мышь | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt9A, Wnt9B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Xenopus | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt11R |
| Danio rerio | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Drosophila | Wg, DWnt2, DWnt3/5, DWnt 4, DWnt6, WntD/DWnt8, DWnt10 |
Структура Wnt-белков
Структура Wnt напоминает кисть руки[8]. Роль большого пальца в этой структуре выполняет аминоконцевой домен, состоящий из пучка α-спиралей, содержащего 10 цистеиновых остатков, которые стабилизируют эту структуру, образуя 5 дисульфидных мостиков. Роль указательного пальца выполняет карбоксиконцевой домен, преимущественно состоящий из двух β-складок, поддерживаемых шестью дисульфидными мостиками. Участок между большим и указательным пальцем — «ладонь» — обладает высокой степенью гибкости. К аминоконцевому домену — «большому пальцу» ковалентно присоединяется ацильная группа — пальмолеиновая жирная кислота . Этот ковалентно присоединенный жир необходим для того, чтобы Wnt мог взаимодействовать с транспортными и с мембранными белками. Изменения аминоконцевого домена белков Wnt могут играть важную роль в регуляции его активности. Так например, трансмембранный белок протеаза Tiki может подавлять Wnt-сигнал, отщепляя восемь аминокислот с аминоконцевого участка[9]. Участок «ладони» является местом посадки гликозильных групп — олигосахаридных цепочек. Степень гликозилирования Wnt никак не влияет на ее активность. Однако предполагается, что N-гликозилирование может влиять на секрецию Wnt, так как негликозилированные молекулы Wnt не могут подвергнуться ацилированию, а значит не могут, как отмечено выше, взаимодействовать с транспортными белками, что необходимо для их секреции. Предполагается, что перенос липида на Wnt осуществляет ацилтранферраза эндоплазматического ретикулума Поркупин (PORCN), так как делеция этого гена нарушает секрецию Wnt[10].
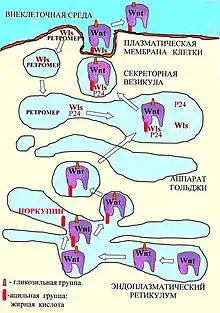
.
Секреция Wnt
После ацилирования белок Wnt становится способен связаться с экскортирующими его белками Аппарата Гольджи, такими как кодируемый геном Wntless трансмембранный белок GPR177 (называемый также Wls)[12] и «белки-грузчики» p24, которые переносят Wnt от эндоплазматического ретикулума на поверхность клетки[13]. Помимо этих белков в секреции Wnt участвует член семейства транспортных белков-липокаинов белок Swim, который поддерживает растворимость и сигнальную активность комплекса Wnt/Wls[14]).
Wnt во внеклеточной среде
Во внеклеточной среде Wnt образует комплексы с гликанами внеклеточного матрикса, что может существенно влиять на мощность и продолжительность их сигнала[15]. Попав во внеклеточную среду, Wnt оказывается вовлеченным в многочисленные процессы межклеточного общения. Инактивировать молекулу Wnt путем удаления липидного остатка (который ранее был посажен на молукулу Wnt Поркупином), и таким образом подавить Wnt сигнализацию, может фермент НОТУМ (NOTUM)[16]. Подавить активность НОТУМ и таким образом повысить активность Wnt сигнализации, позволяют ингибиторы вроде ABC99[17]. Есть данные что такая активация Wnt сигнализации может повысить способность старых клеток кишечного эпителия к регенерации[18]
Мембранные рецепторы Wnt
Для того чтобы воздействовать на клетку-мишень, Wnt должен связаться с клеточными рецепторами. В качестве таких рецепторов на поверхности клетки выступают трансмембранный белок Фрайзлед (Frizzled — Fz) и липопротеиды низкой плотности LRP5 / LRP6. Связаться с ними молекуле Wnt активно мешают различные антагонисты, которым противостоят агонисты.
Секретируемые антагонисты и агонисты Wnt
С секретируемыми молекулами Wnt непосредственно связываются их антагонисты: Wnt-ингибирующий фактор (WIF) и sFRP. Белки DKK и sclerostin (SOST) могут связаться с рецепторами LRP5 / LRP6 для предотвращения образования комплекса Fz-Wnt-LRP6. Белки Shisa, захватив Fz в эндоплазматическом ретикулуме, мешают ему выйти на поверхность клетки. Если же молекулы Wnt образуют комплекс с LRP5 / 6 и Fz, сигнализация активируется. Белок R-spondin (RSPO), стабилизируя рецепторы FZ и LRP 5 / 6, повышает Wnt-сигнализацию.
Механизмы воздействия Wnt на клетку
Отдельные компоненты сигнального пути Wnt консервативны от нематоды Caenorhabditis elegans до человека. Предполагается, что примитивные формы лиганда этого типа имелись уже у предков всех Bilateria, но их происхождение прослеживается от губок и слизевиков.

По традиции механизмы воздействия Wnt на клетку подразделяют на канонический (β-катенин-зависимый) путь, который изменяет концентрацию клеточного β-катенина и в конечном счете контролирует программы генной экспрессии, связанные с определением судьбы клетки и морфогенезом[20] и так называемые неканонические (β-катенин-независимые) пути, которые регулируют полярность клетки, стимулируя реорганизацию цитоскелета[21] и метаболизм кальция[22]. Исторически механизмы воздействия Wnt разделяли на канонические и неканонические, используя три стандартных метода определения: по активности в культуре клеток C57MG, по способности влиять на эмбриогенез у Xenopus и по индукции образования почечных канальцев в изолированной культуре мезенхимальных клеток почки[23]. Однако позже выяснилось, что выбор сигнального пути зависит не столько от лиганда, сколько от рецептора и других белков, составляющих собственно сигнальный путь.
Канонический путь передачи сигнала
В основе канонического пути Wnt-сигнализации лежит стабилизация цитоплазматического белка β-катенина. Канонический путь сигнала Wnt регулируется на многих уровнях, в том числе путем многочисленных противостоящих ему механизмов управления. В отсутствие сигнала β-катенин связан и фосфорилируется «деструктирующим комплексом», содержащим белок — супрессор опухолей — adenomatous polyposis coli (APC), цитоплазматический «поддерживающий» белок Axin, а также протеинкиназу GSK-3 и казеинкиназу (CK1). Фермент GSK-3 ответствен за «запуск» деградации порядка 20 % клеточных белков, давая фосфорилированием «добро» на посадку убиквитина, высококонсервативного белка, который служит в качестве молекулярного тега, обозначающего белки, предназначенные для деградации в протеасоме. Когда клетки получают Wnt, этот лиганд связывает мембранный рецептор семейства Frizzled, который, в свою очередь, активирует белок Dishevelled (DSH), ингибирующий мультибелковый «деструктирующий комплекс», что приводит к снижению скорости деградации β-катенина, так как Wnt «заставляет» GSK-3 оказаться в изоляции, прочно связанным с внутренней стороной мембраны так называемых мультивезикулярных телец цитоплазмы[24] и подавляет убиквитинилирование[25].
Рецептор Frizzled на клеточной мембране связывает белок LRP[26], который кроме Frizzled связывает также Wnt и аксин, стабилизируя комплекс Wnt/Frizzled/LRP/Dishevelled/аксин (рецепторный комплекс).
Когда β-катенин — ключевой внутриклеточный элемент этого сигнального пути — избегает деградации и накапливается в цитоплазме, он входит в ядро, где активирует Т-клеточные факторы (TCF), направляющие β-катенин на гены-мишени Wnt[27] регулирующие экспрессию множества генов.
GSK-3 помимо этого участвует в путях регуляции метаболизма другими важными протеинкиназами, такими как мишень рапамицина — mTOR и AKT. Таким образом, ингибирование GSK-3 под влиянием Wnt затрагивает очень многие механизмы[28][29].
Молекулярный механизм, связывающий Hippo-сигнализацию с сигнализацией Wnt / β-катенина
Наряду с β-катенином посредником сигнала Wnt служит посредник Hippo-сигнализации транскрипционный коактиватор TAZ (известный также как WWTR1). В отсутствие сигнала Wnt деструктирующий комплекс APC, Axin, и GSK3 поддерживает TAZ на низком уровне. Деградация TAZ зависит от фосфорилированного β-катенина, который связывает TAZ с его убиквитин-лигазой β-TrCP. В присутствии Wnt-сигнала β-катенин, избежавший разрушения деструктирующим комплексом, нарушает деградацию TAZ, что приводит к одновременному накоплению β-катенина и TAZ. Оба эти посредника далее действуют на уровне генома, влияя на транскрипцию[30]. Таким образом и некоторыми другими способами[31][32] эти два пути сигнализации могут влиять друг на друга и совместно регулировать рост тканей и их регенерацию.
Неканонические пути передачи сигнала
Помимо рецепторов Frizzled и LRP, Wnt может связываться с рецепторными тримерными G-белками[33][34] тирозинкиназами Ror и Ryk. Ror связавшись с Wnt5a, фосфорилирует белок Дишевеллед и таким образом контролирует морфогенез тканей[35], тогда как Ryk, фосфорилируя мембранный белок клеточной поверхности Vangl2, контролирует полярность клетки[36].
Неканонические (β-катенин-независимые) пути регулируют полярность клетки, стимулируя реорганизацию цитоскелета[37] и метаболизм кальция[22].
Важную роль в механизмах влияния Wnt на клеточные процессы, такие как агрегация клеток — слипание клеток, дифференцировка, пролиферация, онкогенез, секреция внеклеточного матрикса играют белки WISP-1, известный также как CCN4; WISP-2, известный также как CCN5 и WISP-3, известный также как CCN6[38][39]
Регуляция транскрипции генов-мишеней Wnt
В ядре β-катенин, захватив ядерные белки BCL9, пигопус (Pygopus) и некоторые белки-активаторы, становится способен активировать белки TCF/LEF, превращая их в мощные активаторы транскрипции генов-мишеней Wnt. TCF/LEF являются многофункциональными белками, которые, обладая способностью избирательно связываться с определенными последовательностями ДНК и с определенными белками-активаторами, «принимают решение», какие из генов будут активированы сигналом Wnt[42]. Обнаружено что связь между β-катенином и TCF4, необходимая для такой активации, может быть нарушена ресвератролом. Это позволяет предположить что ресвератрол, являющийся флавоноидом кожицы черного винограда и получаемого из него вина, может быть использован в качестве безвредного лекарства для подавления сигнала Wnt при раковых заболеваниях[43] .
Выяснилось что β-катенин, избежавший деградации благодаря сигналу Wnt, активирует синтез ферментативной субъединицы теломеразы (TERT) в стволовых и раковых клетках. В этом ему помогает один из транскрипционных факторов плюрипотенции — Klf4, направляющий его на промотор гена Tert[44] .
Роль Wnt / β-катенин в стволовых клетках
Стволовые клетки в основном характеризуются двумя свойствами: способностью к самообновлению и способностью к дифференцировке в различные типы клеток. Эти процессы регулируются различными факторами роста, в том числе белками Wnt[29]. Накопленные данные показывают, что сигнальный путь Wnt / β-катенина играет ключевую роль в поддержании плюрипотентности, а также в процессах перепрограммирования соматических клеток. Так, в частности, рецептор WNT из семейства Frizzled, называемый FZD7, играет ключевую роль в поддержании плюрипотентных клеток в недифференцированном и плюрипотентном состоянии[45], а также регулирует фенотип клеток их пролиферацию и морфологию[46]. В то же время сигнализация Wnt / β-катенин играет также важную роль и в процессах детерминации и дифференцировки, в частности, путем регуляции синтеза морфогена Sox17[47]. Обнаружено что добавление белка Wnt или же, наоборот, малой молекулы ингибитора Wnt (IWP2) снижает неоднородность популяции клеток, при этом соответственно образуются либо клетки со стабильно высоким уровнем синтеза Wnt, либо клетки с низким уровнем синтеза Wnt. При дифференцировке эмбриональные клетки с высоким уровнем синтеза Wnt преимущественно образуют энтодермальные и сердечные клетки, а с низким образуют в первую очередь нейроэктодермальные клетки[48] . Знание того, что сигнализация Wnt на ранних стадиях дифференцировки повышает, а на поздних стадиях наоборот угнетает развитие сердца, позволило путем правильной стратегии использования малых молекул и механизмов сигнализации Wnt получить in vitro из индуцированных плюрипотентных стволовых клеток человека кардиомиоциты с недостижимой до сих пор эффективностью до 98 %[49].
Считается, что одной из основных причин истощения тканей и активации возрастных заболеваний при старении организма является снижение качества и количества соматических стволовых клеток. Важную роль в этом процессе, как выяснилось, играет переход от канонической к неканонической Wnt-сигнализации, вызванный повышением при старении синтеза Wnt5a, который вызывает одряхление стволовых клеток, проявляющееся в аполярности, снижении регенеративной способности и сдвиге дифференциации с лимфоидного на миелоидный тип клеток, вследствие активации небольшой Rho ГТФазы, называемой Cdc42[50] и перестройки актинового цитоскелета.[51][52]
Рецептором Wnt5a является белок FZD5. Нокдаун гена FZD5 приводит к потере свойств стволовых клеток, включая поддержание пролиферации и мультилинейной потенции, тогда как сверхэкспрессия FZD5 ингибирует старение мезенхимальных стволовых/ стромальных клеток человека.[53]
Влияние Wnt на клеточный цикл и пролиферацию клеток
Появляется все больше доказательств сложного взаимодействия канонического пути Wnt-сигнализации и клеточного цикла. Сигнализация Wnt сильно активируется в митозе, свидетельствуя о том, что «митотическая Wnt-сигнализация» играет важную роль в организации программы клеточного деления и таким образом способствует клеточной пролиферации[54][55][56]. Сигнал Wnt может действовать на пролиферацию клеток, активируя транскрипцию циклина D1, c-myc и CDC20-опосредованного контроля деградации белка conductin, которые контролируют переход G1 / S клеточного цикла, а также с помощью комплекса, содержащего Cdk14 (PFTK1) и циклин Y[57]. Компоненты сигнального каскада Wnt действуют непосредственно на формирование митотического веретена. Так например, у излюбленного модельного организма исследователей — червячка C. elegans — сигнализация Wnt вызывает асимметрию митотического веретена, приводящую к асимметричному распределению β-катенина[58].
Роль Wnt в регенерации
В молодости у млекопитающих область кончика пальцев может регенерировать после ампутации подобно тому, как это происходит у амфибий. В этой регенерации принимают участие стволовые клетки, базирующиеся в матриксе кончиков пальца. В этом процессе важнейшая роль отведена Wnt, который необходим для дифференцировки этих стволовых клеток, а также для направленного подсоединения к ним нервов, без которых невозможен рост мезенхимальной бластемы и дальнейшая регенерация.[59]
Люди, пережившие инфаркт миокарда, нередко становятся инвалидами из-за прогрессирующей сердечной недостаточности, вызванной заменой мышечной ткани на рубцовую. Обнаружено, что химическое ингибирование Wnt-ацилтрансферазы эндоплазматического ретикулума — поркупина, ингибируя секрецию Wnt, приводит к существенному улучшению функции сердца после инфаркта миокарда у мышей[60][61]. Воздействие ингибитора Поркупина (WNT974/LGK‑974) на пострадавшие от инфаркта ткани сердца приводило к ослаблению процессов рубцевания и повышению способности сердца перекачивать кровь. Поэтому ингибиторы Поркупина потенциально могут быть использованы для предотвращения сердечной недостаточности после инфаркта миокарда.
Примечания
- Yang Y. Wnt signaling in development and disease. (англ.) // Cell & Bioscience. — 2012. — 20 April (vol. 2, no. 1). — P. 14—14. — doi:10.1186/2045-3701-2-14. — PMID 22520685.
- Lie D. C., Colamarino S. A., Song H. J., Désiré L., Mira H., Consiglio A., Lein E. S., Jessberger S., Lansford H., Dearie A. R., Gage F. H. Wnt signalling regulates adult hippocampal neurogenesis. (англ.) // Nature. — 2005. — 27 October (vol. 437, no. 7063). — P. 1370—1375. — doi:10.1038/nature04108. — PMID 16251967.
- Rijsewijk F., Schuermann M., Wagenaar E., Parren P., Weigel D., Nusse R. The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. (англ.) // Cell. — 1987. — 14 August (vol. 50, no. 4). — P. 649—657. — PMID 3111720.
- Sharma R. P., Chopra V. L. Effect of the Wingless (wg1) mutation on wing and haltere development in Drosophila melanogaster. (англ.) // Developmental Biology. — 1976. — February (vol. 48, no. 2). — P. 461—465. — PMID 815114.
- Nüsslein-Volhard C., Wieschaus E. Mutations affecting segment number and polarity in Drosophila. (англ.) // Nature. — 1980. — 30 October (vol. 287, no. 5785). — P. 795—801. — PMID 6776413.
- Wu J., Cohen S. M. Repression of Teashirt marks the initiation of wing development. (англ.) // Development (Cambridge, England). — 2002. — May (vol. 129, no. 10). — P. 2411—2418. — PMID 11973273.
- Nusse R., van Ooyen A., Cox D., Fung Y. K., Varmus H. Mode of proviral activation of a putative mammary oncogene (int-1) on mouse chromosome 15. (англ.) // Nature. — 1984. — 12 January (vol. 307, no. 5947). — P. 131—136. — PMID 6318122.
- Willert K., Nusse R. Wnt proteins. (англ.) // Cold Spring Harbor Perspectives In Biology. — 2012. — 1 September (vol. 4, no. 9). — P. 007864—007864. — doi:10.1101/cshperspect.a007864. — PMID 22952392.
- Zhang X., Abreu J. G., Yokota C., MacDonald B. T., Singh S., Coburn K. L., Cheong S. M., Zhang M. M., Ye Q. Z., Hang H. C., Steen H., He X. Tiki1 is required for head formation via Wnt cleavage-oxidation and inactivation. (англ.) // Cell. — 2012. — 22 June (vol. 149, no. 7). — P. 1565—1577. — doi:10.1016/j.cell.2012.04.039. — PMID 22726442.
- Barrott J. J., Cash G. M., Smith A. P., Barrow J. R., Murtaugh L. C. Deletion of mouse Porcn blocks Wnt ligand secretion and reveals an ectodermal etiology of human focal dermal hypoplasia/Goltz syndrome. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2011. — 2 August (vol. 108, no. 31). — P. 12752—12757. — doi:10.1073/pnas.1006437108. — PMID 21768372.
- Джагаров Д. (2012) Важнейшие стрелочники клеток организма: белки Wnt
- Herr P., Basler K. Porcupine-mediated lipidation is required for Wnt recognition by Wls. (англ.) // Developmental Biology. — 2012. — 15 January (vol. 361, no. 2). — P. 392—402. — doi:10.1016/j.ydbio.2011.11.003. — PMID 22108505.
- Buechling T., Chaudhary V., Spirohn K., Weiss M., Boutros M. p24 proteins are required for secretion of Wnt ligands. (англ.) // EMBO Reports. — 2011. — 1 December (vol. 12, no. 12). — P. 1265—1272. — doi:10.1038/embor.2011.212. — PMID 22094269.
- Mulligan K. A., Fuerer C., Ching W., Fish M., Willert K., Nusse R. Secreted Wingless-interacting molecule (Swim) promotes long-range signaling by maintaining Wingless solubility. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2012. — 10 January (vol. 109, no. 2). — P. 370—377. — doi:10.1073/pnas.1119197109. — PMID 22203956.
- Berendsen A. D., Fisher L. W., Kilts T. M., Owens R. T., Robey P. G., Gutkind J. S., Young M. F. Modulation of canonical Wnt signaling by the extracellular matrix component biglycan. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2011. — 11 October (vol. 108, no. 41). — P. 17022—17027. — doi:10.1073/pnas.1110629108. — PMID 21969569.
- Kakugawa S., Langton P.F., Zebisch M., et al., (2015). Notum deacylates Wnt proteins to suppress signalling activity. Nature. 519(7542), 187–192. doi:10.1038/nature14259 PMC 4376489
- Suciu, R. M., Cognetta III, A. B., Potter, Z. E., & Cravatt, B. F. (2018). Selective irreversible inhibitors of the Wnt-deacylating enzyme NOTUM developed by activity-based protein profiling. ACS medicinal chemistry letters, 9(6), 563-568. doi:10.1021/acsmedchemlett.8b00191 PMC 6004566
- Pentinmikko, N., Iqbal, S., Mana, M., et al., & Smolander, O. P. (2019). Notum produced by Paneth cells attenuates regeneration of aged intestinal epithelium. Nature, 571(7765), 398-402. doi:10.1038/s41586-019-1383-0.
- Randall T. Moon (2013) Canonical Wnt/-catenin Signaling
- van Amerongen R., Nusse R. Towards an integrated view of Wnt signaling in development. (англ.) // Development (Cambridge, England). — 2009. — October (vol. 136, no. 19). — P. 3205—3214. — doi:10.1242/dev.033910. — PMID 19736321.
- Goodrich L. V., Strutt D. Principles of planar polarity in animal development. (англ.) // Development (Cambridge, England). — 2011. — May (vol. 138, no. 10). — P. 1877—1892. — doi:10.1242/dev.054080. — PMID 21521735.
- Kohn A. D., Moon R. T. Wnt and calcium signaling: beta-catenin-independent pathways. (англ.) // Cell Calcium. — 2005. — September (vol. 38, no. 3-4). — P. 439—446. — doi:10.1016/j.ceca.2005.06.022. — PMID 16099039.
- Merkel C. E., Karner C. M., Carroll T. J. Molecular regulation of kidney development: is the answer blowing in the Wnt? (англ.) // Pediatric Nephrology (Berlin, Germany). — 2007. — November (vol. 22, no. 11). — P. 1825—1838. — doi:10.1007/s00467-007-0504-4. — PMID 17554566.
- Taelman V. F., Dobrowolski R., Plouhinec J. L., Fuentealba L. C., Vorwald P. P., Gumper I., Sabatini D. D., De Robertis E. M. Wnt signaling requires sequestration of glycogen synthase kinase 3 inside multivesicular endosomes. (англ.) // Cell. — 2010. — 23 December (vol. 143, no. 7). — P. 1136—1148. — doi:10.1016/j.cell.2010.11.034. — PMID 21183076.
- Li V. S., Ng S. S., Boersema P. J., Low T. Y., Karthaus W. R., Gerlach J. P., Mohammed S., Heck A. J., Maurice M. M., Mahmoudi T., Clevers H. Wnt signaling through inhibition of β-catenin degradation in an intact Axin1 complex. (англ.) // Cell. — 2012. — 8 June (vol. 149, no. 6). — P. 1245—1256. — doi:10.1016/j.cell.2012.05.002. — PMID 22682247.
- Wehrli M., Dougan S. T., Caldwell K., O'Keefe L., Schwartz S., Vaizel-Ohayon D., Schejter E., Tomlinson A., DiNardo S. arrow encodes an LDL-receptor-related protein essential for Wingless signalling. (англ.) // Nature. — 2000. — 28 September (vol. 407, no. 6803). — P. 527—530. — doi:10.1038/35035110. — PMID 11029006.
- Cadigan K. M. TCFs and Wnt/β-catenin signaling: more than one way to throw the switch. (англ.) // Current Topics In Developmental Biology. — 2012. — Vol. 98. — P. 1—34. — doi:10.1016/B978-0-12-386499-4.00001-X. — PMID 22305157.
- Palsgaard J., Emanuelli B., Winnay J. N., Sumara G., Karsenty G., Kahn C. R. Cross-talk between insulin and Wnt signaling in preadipocytes: role of Wnt co-receptor low density lipoprotein receptor-related protein-5 (LRP5). (англ.) // The Journal Of Biological Chemistry. — 2012. — 6 April (vol. 287, no. 15). — P. 12016—12026. — doi:10.1074/jbc.M111.337048. — PMID 22337886.
- Muñoz-Descalzo S., de Navascues J., Arias A. M. Wnt-Notch signalling: an integrated mechanism regulating transitions between cell states. (англ.) // BioEssays : news and reviews in molecular, cellular and developmental biology. — 2012. — Vol. 34, no. 2. — P. 110—118. — doi:10.1002/bies.201100102. — PMID 22215536.
- Azzolin L., Zanconato F., Bresolin S., Forcato M., Basso G., Bicciato S., Cordenonsi M., Piccolo S. Role of TAZ as mediator of Wnt signaling. (англ.) // Cell. — 2012. — 21 December (vol. 151, no. 7). — P. 1443—1456. — doi:10.1016/j.cell.2012.11.027. — PMID 23245942.
- Konsavage Jr. W. M., Yochum G. S. Intersection of Hippo/YAP and Wnt/β-catenin signaling pathways. (англ.) // Acta Biochimica Et Biophysica Sinica. — 2013. — February (vol. 45, no. 2). — P. 71—79. — doi:10.1093/abbs/gms084. — PMID 23027379.
- Imajo M., Miyatake K., Iimura A., Miyamoto A., Nishida E. A molecular mechanism that links Hippo signalling to the inhibition of Wnt/β-catenin signalling. (англ.) // The EMBO Journal. — 2012. — 7 March (vol. 31, no. 5). — P. 1109—1122. — doi:10.1038/emboj.2011.487. — PMID 22234184.
- Katanaev V. L., Ponzielli R., Sémériva M., Tomlinson A. Trimeric G protein-dependent frizzled signaling in Drosophila. (англ.) // Cell. — 2005. — 14 January (vol. 120, no. 1). — P. 111—122. — doi:10.1016/j.cell.2004.11.014. — PMID 15652486.
- Liu X., Rubin J. S., Kimmel A. R. Rapid, Wnt-induced changes in GSK3beta associations that regulate beta-catenin stabilization are mediated by Galpha proteins. (англ.) // Current Biology : CB. — 2005. — 22 November (vol. 15, no. 22). — P. 1989—1997. — doi:10.1016/j.cub.2005.10.050. — PMID 16303557.
- Ho H. Y., Susman M. W., Bikoff J. B., Ryu Y. K., Jonas A. M., Hu L., Kuruvilla R., Greenberg M. E. Wnt5a-Ror-Dishevelled signaling constitutes a core developmental pathway that controls tissue morphogenesis. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2012. — 13 March (vol. 109, no. 11). — P. 4044—4051. — doi:10.1073/pnas.1200421109. — PMID 22343533.
- Macheda M. L., Sun W. W., Kugathasan K., Hogan B. M., Bower N. I., Halford M. M., Zhang Y. F., Jacques B. E., Lieschke G. J., Dabdoub A., Stacker S. A. The Wnt receptor Ryk plays a role in mammalian planar cell polarity signaling. (англ.) // The Journal Of Biological Chemistry. — 2012. — 24 August (vol. 287, no. 35). — P. 29312—29323. — doi:10.1074/jbc.M112.362681. — PMID 22773843.
- May-Simera H. L., Kelley M. W. Cilia, Wnt signaling, and the cytoskeleton. (англ.) // Cilia. — 2012. — 2 May (vol. 1, no. 1). — P. 7—7. — doi:10.1186/2046-2530-1-7. — PMID 23351924.
- Liu, Y., Song, Y., Ye, M., Hu, X., Wang, Z. P., & Zhu, X. (2019). The emerging role of WISP proteins in tumorigenesis and cancer therapy. Journal of translational medicine, 17(1), 28. PMC 6335850
- Ono, M., Masaki, A., Maeda, A., Kilts, T. M., Hara, E. S., Komori, T., ... & Young, M. F. (2018). CCN4/WISP1 controls cutaneous wound healing by modulating proliferation, migration and ECM expression in dermal fibroblasts via α5β1 and TNFα. Matrix Biology. PMC 6015535
- Gómez-Orte E., Sáenz-Narciso B., Moreno S., Cabello J. Multiple functions of the noncanonical Wnt pathway. (англ.) // Trends In Genetics : TIG. — 2013. — September (vol. 29, no. 9). — P. 545—553. — doi:10.1016/j.tig.2013.06.003. — PMID 23846023.
- Green J., Nusse R., van Amerongen R. The role of Ryk and Ror receptor tyrosine kinases in Wnt signal transduction. (англ.) // Cold Spring Harbor perspectives in biology. — 2014. — Vol. 6, no. 2. — doi:10.1101/cshperspect.a009175. — PMID 24370848.
- Cadigan K. M., Waterman M. L. TCF/LEFs and Wnt signaling in the nucleus. (англ.) // Cold Spring Harbor perspectives in biology. — 2012. — Vol. 4, no. 11. — doi:10.1101/cshperspect.a007906. — PMID 23024173.
- Chen H. J., Hsu L. S., Shia Y. T., Lin M. W., Lin C. M. The β-catenin/TCF complex as a novel target of resveratrol in the Wnt/β-catenin signaling pathway. (англ.) // Biochemical pharmacology. — 2012. — Vol. 84, no. 9. — P. 1143—1153. — doi:10.1016/j.bcp.2012.08.011. — PMID 22935447.
- Hoffmeyer K., Raggioli A., Rudloff S., Anton R., Hierholzer A., Del Valle I., Hein K., Vogt R., Kemler R. Wnt/β-catenin signaling regulates telomerase in stem cells and cancer cells. (англ.) // Science (New York, N.Y.). — 2012. — Vol. 336, no. 6088. — P. 1549—1554. — doi:10.1126/science.1218370. — PMID 22723415.
- Fernandez A., Huggins I. J., Perna L., Brafman D., Lu D., Yao S., Gaasterland T., Carson D. A., Willert K. The WNT receptor FZD7 is required for maintenance of the pluripotent state in human embryonic stem cells. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2014. — Vol. 111, no. 4. — P. 1409—1414. — doi:10.1073/pnas.1323697111. — PMID 24474766.
- Zhang Z., Rankin S. A., Zorn A. M. Different thresholds of Wnt-Frizzled 7 signaling coordinate proliferation, morphogenesis and fate of endoderm progenitor cells. (англ.) // Developmental biology. — 2013. — Vol. 378, no. 1. — P. 1—12. — doi:10.1016/j.ydbio.2013.02.024. — PMID 23562607.
- Engert S., Burtscher I., Liao W. P., Dulev S., Schotta G., Lickert H. Wnt/β-catenin signalling regulates Sox17 expression and is essential for organizer and endoderm formation in the mouse. (англ.) // Development (Cambridge, England). — 2013. — Vol. 140, no. 15. — P. 3128—3138. — doi:10.1242/dev.088765. — PMID 23824574.
- Blauwkamp T. A., Nigam S., Ardehali R., Weissman I. L., Nusse R. Endogenous Wnt signalling in human embryonic stem cells generates an equilibrium of distinct lineage-specified progenitors. (англ.) // Nature communications. — 2012. — Vol. 3. — P. 1070. — doi:10.1038/ncomms2064. — PMID 22990866.
- Lian X., Hsiao C., Wilson G., Zhu K., Hazeltine L. B., Azarin S. M., Raval K. K., Zhang J., Kamp T. J., Palecek S. P. Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2012. — Vol. 109, no. 27. — P. 1848—1857. — doi:10.1073/pnas.1200250109. — PMID 22645348.
- Florian M. C., Nattamai K. J., Dörr K., Marka G., Uberle B., Vas V., Eckl C., Andrä I., Schiemann M., Oostendorp R. A., Scharffetter-Kochanek K., Kestler H. A., Zheng Y., Geiger H. A canonical to non-canonical Wnt signalling switch in haematopoietic stem-cell ageing. (англ.) // Nature. — 2013. — Vol. 503, no. 7476. — P. 392—396. — doi:10.1038/nature12631. — PMID 24141946.
- Schreck C., Istvánffy R., Ziegenhain C., Sippenauer T., Ruf F., Henkel L., Gärtner F., Vieth B., Florian M. C., Mende N., Taubenberger A., Wagner A., Pagel C., Grziwok S., Götze K. S., Guck J., Dean D. C., Massberg S., Essers M., Waskow C., Geiger H., Schiemann M., Peschel C., Enard W., Oostendorp R. A. Niche WNT5A regulates the actin cytoskeleton during regeneration of hematopoietic stem cells. (англ.) // The Journal Of Experimental Medicine. — 2017. — January (vol. 214, no. 1). — P. 165—181. — doi:10.1084/jem.20151414. — PMID 27998927.
- Schreck, C., Istvánffy, R., Ziegenhain, C., Sippenauer, T., Ruf, F., Henkel, L., ... & Oostendorp, R. A. (2017). Niche WNT5A regulates the actin cytoskeleton during regeneration of hematopoietic stem cells. Journal of Experimental Medicine, 214(1), 165-181. PMID 27998927 PMC 5206491 doi:10.1084/jem.20151414
- Harada, S., Mabuchi, Y., Kohyama, J., Shimojo, D., Suzuki, S., Kawamura, Y., ... & Matsuzaki, Y. (2020). FZD5 regulates cellular senescence in human mesenchymal stem/stromal cells. Stem Cells. PMID 33338299 doi:10.1002/stem.3317
- Niehrs C., Acebron S. P. Mitotic and mitogenic Wnt signalling. (англ.) // The EMBO journal. — 2012. — Vol. 31, no. 12. — P. 2705—2713. — doi:10.1038/emboj.2012.124. — PMID 22617425.
- Gougelet A., Colnot S. A Complex Interplay between Wnt/β-Catenin Signalling and the Cell Cycle in the Adult Liver. (англ.) // International journal of hepatology. — 2012. — Vol. 2012. — P. 816125. — doi:10.1155/2012/816125. — PMID 22973520.
- Hadjihannas M. V., Bernkopf D. B., Brückner M., Behrens J. Cell cycle control of Wnt/β-catenin signalling by conductin/axin2 through CDC20. (англ.) // EMBO reports. — 2012. — Vol. 13, no. 4. — P. 347—354. — doi:10.1038/embor.2012.12. — PMID 22322943.
- Kaldis P., Pagano M. Wnt signaling in mitosis. (англ.) // Developmental cell. — 2009. — Vol. 17, no. 6. — P. 749—750. — doi:10.1016/j.devcel.2009.12.001. — PMID 20059944.
- Sugioka K., Mizumoto K., Sawa H. Wnt regulates spindle asymmetry to generate asymmetric nuclear β-catenin in C. elegans. (англ.) // Cell. — 2011. — Vol. 146, no. 6. — P. 942—954. — doi:10.1016/j.cell.2011.07.043. — PMID 21925317.
- Takeo M., Chou W. C., Sun Q., Lee W., Rabbani P., Loomis C., Taketo M. M., Ito M. Wnt activation in nail epithelium couples nail growth to digit regeneration. (англ.) // Nature. — 2013. — Vol. 499, no. 7457. — P. 228—232. — doi:10.1038/nature12214. — PMID 23760480.
- Cancer drug could promote regeneration of heart tissue. ScienceDaily, 3 February 2017
- Moon J., Zhou H., Zhang L. S., Tan W., Liu Y., Zhang S., Morlock L. K., Bao X., Palecek S. P., Feng J. Q., Williams N. S., Amatruda J. F., Olson E. N., Bassel-Duby R., Lum L. Blockade to pathological remodeling of infarcted heart tissue using a porcupine antagonist. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2017. — Vol. 114, no. 7. — P. 1649—1654. — doi:10.1073/pnas.1621346114. — PMID 28143939.
Литература
- J. Behrens (2013) Everything You Would Like to Know About Wnt Signaling. Sci. Signal. 6(275), pe17 DOI: 10.1126/scisignal.2004167
- Wnt Signaling. Cold Spring Harbor Perspectives in Biology. CSHL Press 2013, ISBN 978-1-936113-23-1
- Van Camp JK, Beckers S, Zegers D, Van Hul W. (2014 Apr.). Wnt signaling and the control of human stem cell fate. Stem Cell Rev.; 10(2), 207—229. doi:10.1007/s12015-013-9486-8 PMID 24323281
- Xiao, Y. F., Yong, X., Tang, B., Qin, Y., Zhang, J. W., Zhang, D., … & Yang, S. M. (Feb 2016). Notch and Wnt signaling pathway in cancer: Crucial role and potential therapeutic targets (Review). International journal of oncology, 48(2), 437—449 doi:10.3892/ijo.2015.3280
- Путем ингибирования сигналов Wnt в эмбриональных стволовых клетках, с помощью ингибитора сигнального пути Wnt DKK1, можно заставить эмбриональные стволовые клетки мыши, дифференцироваться в энтодерму и, что особенно важно, подавить онкогенность эмбриональных стволовых клеток. Ou, L., Fang, L., Tang, H., Qiao, H., Zhang, X., & Wang, Z. (2016). Dickkopf Wnt signaling pathway inhibitor 1 regulates the differentiation of mouse embryonic stem cells in vitro and in vivo. Molecular medicine reports, 13(1), 720—730. doi:10.3892/mmr.2015.4586 PMC 4686056
- Lehoczky, J. A., & Tabin, C. J. (2018). Rethinking WNT signalling. The identification of genetic mutations that can hinder the development of human limbs has led to the discovery of an unanticipated mode of regulation for the WNT signalling pathway during limb development. Nature 557, 495-496 doi:10.1038/d41586-018-04820-y
- Szenker-Ravi, E., Altunoglu, U., Leushacke, M., Bosso-Lefèvre, C., Khatoo, M., Tran, H., ... & Sousa, S. B. (2018). RSPO2 inhibition of RNF43 and ZNRF3 governs limb development independently of LGR4/5/6. Nature 557, 564–569 doi:10.1038/s41586-018-0118-y
- Matthias C. Vogg, Leonardo Beccari, Laura Iglesias Ollé, Christine Rampon, Sophie Vriz, Chrystelle Perruchoud, Yvan Wenger & Brigitte Galliot (2019). An evolutionarily-conserved Wnt3/β-catenin/Sp5 feedback loop restricts head organizer activity in Hydra. Nature Communicationsvolume 10, Article number: 312(2019) https://doi.org/10.1038/s41467-018-08242-2
- Kakugawa S, Langton PF, Zebisch M, Howell S, Chang TH, Liu Y, Feizi T, Bineva G, O'Reilly N, Snijders AP, Jones EY, Vincent JP (2015). Notum deacylates Wnt proteins to suppress signalling activity. Nature. 519(7542), 187–192. doi:10.1038/nature14259. PMC 4376489.
- Suciu, R. M., Cognetta III, A. B., Potter, Z. E., & Cravatt, B. F. (2018). Selective irreversible inhibitors of the Wnt-deacylating enzyme NOTUM developed by activity-based protein profiling. ACS medicinal chemistry letters, 9(6), 563-568. doi:10.1021/acsmedchemlett.8b00191 PMC 6004566
- Pentinmikko, N., Iqbal, S., Mana, M., Andersson, S., Cognetta, A. B., Suciu, R. M., ... & Smolander, O. P. (2019). Notum produced by Paneth cells attenuates regeneration of aged intestinal epithelium. Nature, 571, 398–402 https://doi.org/10.1038/s41586-019-1383-0
- Grainger, S., & Willert, K. (2018). Mechanisms of Wnt signaling and control. Wiley Interdisciplinary Reviews: Systems Biology and Medicine, 10(5), e1422. doi:10.1002/wsbm.1422 PMC 6165711