Конодонты
Конодонты[1] (лат. Conodonta, от др.-греч. κώνος — конус и др.-греч. ὀδούς — зуб) — класс вымерших животных из типа хордовых, внешним обликом были похожи на современных миног и миксин.
| † Конодонты | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
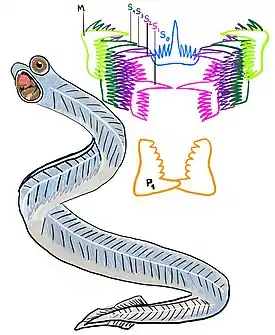 Реконструкция Hindeodus, показывающая расположение конодонтовых элементов | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Класс: † Конодонты |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Conodonta Eichenberg, 1930 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология 541—182,7 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
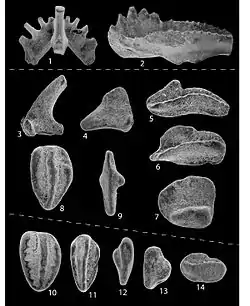
До начала 1980-х годов конодонты (в настоящее время их называют конодонтовыми элементами) были известны только по микроскопическим (0,1—1 мм) окаменелостям зубовидных скелетных остатков, обнаруженных в морских осадках различного геологического возраста (от позднего кембрия до позднего триаса включительно). При этом было установлено, что группа конодонтов объединяет ископаемые скелетные элементы, принадлежавшие различным группам животных: протоконодонты, параконодонты и эуконодонты. Самих животных теперь также называют конодонтоносителями (Conodontophora), чтобы избежать смешения понятий.
Благодаря быстрому эволюционному изменению морфологии зубных элементов конодонтов они широко используются для биостратиграфии и целей геологического картирования.
Описание
По одиннадцати известным полным окаменелым отпечаткам конодонтоносителей можно сделать вывод, что они по внешнему виду напоминали миног и миксин. Их ротовой аппарат состоял из 15 или, реже, 19 элементов и радикально отличался от челюстей современных животных. Форма элементов — зубовидная, гребенчатая, листообразная; состав — фосфат кальция.

Среди конодонтоносителей были как совсем крошечные (длиной около 1 см), так и гигантские (например, Promissum, длина которого достигала 40 см)[2]. В настоящее время палеонтологи сходятся во мнении, что для конодонтоносителей характерно наличие больших глаз, плавников с плавниковыми лучами, хорды и мощной поперечно расположенной мускулатуры.
Конодонтовые элементы состоят из минерала гидроксиапатита[3][4], который является основной минеральной составляющей костей и зубов позвоночных. Это свидетельствует в пользу того, что зубы были первыми твёрдыми структурами, содержащими гидроксиапатит[4].
Образ жизни
По мнению исследователей, «зубы» некоторых конодонтов представляли собой подобие фильтровальных аппаратов, при помощи которых планктон отфильтровывался из воды и отправлялся в глотку. Другие зубы, исходя из их строения, по их мнению, предназначались для того, чтобы «хватать и разрывать плоть»[2].
Сохранившиеся отпечатки мускулатуры позволяют предположить, что некоторые конодонты (во всяком случае, Promissum) были умелыми пловцами, неспособными, однако, к стремительным броскам[2].
Систематика
Конодонтов в настоящее время относят к хордовым благодаря наличию плавников с плавниковыми лучами, хорды и мощной поперечно расположенной мускулатуры[5], а также сходству химического состава зубов рыб и скелетных элементов конодонтов[6].
Некоторые исследователи относят конодонтов к подтипу позвоночных. Своим внешним обликом они похожи на современных миксин и миног[7].
Филогенетический анализ 2000 года показывает, что эуконодонты, вероятнее всего, являются позвоночными[8]. Тем не менее, этот же анализ вызывает противоречие: ранние формы конодонтов, протоконодонты, формируют отдельную филогенетическую ветвь от более поздних параконодонтов и эуконодонтов. Возможно, протоконодонты связаны с группой, включающей ныне существующих щетинкочелюстных, и не являются родственниками собственно конодонтов[9].
Филогенетический анализ 2020 года восстановил эуконодонтов в качестве сестринского таксона круглоротых[10]:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Окаменелости зубов конодонтов
Многие десятилетия (до начала 1980-х годов) конодонтоносители были известны только по микроскопическим (0,1—1 мм) окаменелостям зубовидных скелетных остатков, которые встречались повсеместно, но их не удавалось «привязать» к какой либо полной окаменелости. Эти микроокаменелости сейчас называют «конодонтовыми элементами», чтобы избежать смешения понятий.
Конодонтовые элементы находят в отложениях с кембрия по триас. Конодонтоносители на протяжении 250 миллионов лет относительно быстро эволюционировали и были широко распространены, что даёт возможность использовать их в биостратиграфии — для установления очень дробных стратиграфических подразделений и для корреляции отложений из разных районов. Кроме того, конодонтовые элементы (зубы) используют как палеотермометры, позволяющие выявлять температурные колебания, происходившие с породой, в которой они содержатся. Дело в том, что под влиянием повышенной температуры в фосфатах происходят предсказуемые и постоянные изменения окраски, которые можно измерить. Это позволяет использовать их в нефтеразведке.
До 1980-х годов конодонтовые элементы обнаруживались лишь отдельно, а поэтому невозможно было определить, частями какого организма они являются[11]. Крайняя редкость обнаружения полных окаменелостей связана с тем, что тело этих животных в основном состояло из мягких тканей.
Начиная с 1982 года было обнаружено несколько полных отпечатков конодонтоносителя в прослое ленточного известняка, находящегося в песчаниках Грантона нижнекаменноугольного возраста вблизи Эдинбурга (Шотландия). Это животное имело длинное (около 40 мм), узкое (около 2 мм) червеобразное тело с отчётливо обозначенной головой. Недалеко от переднего конца сохранился почти полный ротовой аппарат, состоящий из различных эуконодонтовых элементов. От передней части тела к задней отчётливо протягивается узкая, светлая полоса, которую интерпретировали как хорду, на основании чего само это животное было отнесено к примитивным позвоночным[12].
На территории России конодонтовые элементы впервые были обнаружены в 1971 году в разрезах Южного Приморья. К настоящему времени по конодонтам весьма детально проведена корреляция триасовых отложений Сихотэ-Алиня. Благодаря исследованию конодонтов с применением микропалеонтологического метода был установлен триасовый возраст многих кремнистых, вулканогенно-кремнистых и карбонатно-кремнистых толщ Сихотэ-Алиня, считавшихся палеозойскими или юрскими[13].
Примечания
- Нельсон Д. С. Рыбы мировой фауны / Пер. 4-го перераб. англ. изд. Н. Г. Богуцкой, науч. ред-ры А. М. Насека, А. С. Герд. — М.: Книжный дом «ЛИБРОКОМ», 2009. — С. 73. — ISBN 978-5-397-00675-0.
- Gabbott S. E., Aldridge R. J., Theron J. N. A giant conodont with preserved muscle tissue from the Upper Ordovician of South Africa (англ.) // Nature : journal. — 1995. — Vol. 374. — P. 800—803. — doi:10.1038/374800a0.
- Trotter J. A., Eggins S. M. Chemical systematics of conodont apatite determined by laser ablation ICPMS (англ.) // Chemical Geology : journal. — 2006. — Vol. 233, iss. 3. — P. 196—216. — ISSN 0009-2541. — doi:10.1016/j.chemgeo.2006.03.004. — .
- Шубин Н. Внутренняя рыба. История человеческого тела с древнейших времён до наших дней / пер. с англ. П. Петрова. — М.: Астрель : CORPUS, 2010. — С. 101—106. — 303 с. — (ЭЛЕМЕНТЫ). — ISBN 978-5-271-26000-1.
- Briggs D. Conodonts: a major extinct group added to the vertebrates (англ.) // Science : journal. — 1992. — Vol. 256. — P. 1285—1286. — doi:10.1126/science.1598571. — PMID 1598571.
- Касаткина А. П., Бурий Г. И. Chaetodonta — новый надтип животных и его положение в системе животного царства.
- Milsom C. & Rigby S. 2004. Fossils at a Glance. Victoria, Australia: Blackwell Publishing, 155 pp.
- Donoghue P. C. J., Forey P. L., Aldridge R. J. Conodont affinity and chordate phylogeny (англ.) // Biological Reviews. — 2000. — Vol. 75, no. 2. — P. 191—251. — doi:10.1017/S0006323199005472.
- Szaniawski H. New evidence for the protoconodont origin of chaetognaths (англ.) // Acta Palaeontologica Polonica : journal. — 2002. — Vol. 47, no. 3. — P. 405. — doi:10.1126/science.1137187. — PMID 17332408. Архивировано 25 сентября 2007 года.
- Miyashita T., Coates M. I., Farrar R., Larson P., Manning P. L., Wogelius R. A., Edwards N. P., Anné J., Bergmann U., Palmer A. R., Currie P. J. Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny (англ.) // PNAS : journal. — 2019. — Vol. 116, iss. 6. — P. 2146—2151. — ISSN 1091-6490. — doi:10.1073/pnas.1814794116. — .
- Briggs D. E. G., Clarkson E. N. K., Aldridge R. J. The conodont animal (неопр.) // Lethaia. — 1983. — Т. 16. — С. 1—14. — doi:10.1111/j.1502-3931.1983.tb01139.x.
- Касаткина А. П., Бурий Г. И. О связи конодонтов с хетогнатами. — Тихоокеанский океанологический институт ДВО РАН, 1997. — Архивная копия от 25 декабря 2007 на Wayback Machine.
- Бурий Г. И. Конодонты Сихотэ-Алиня. Геолого-минералогический музей Приморья. — Архивная копия от 3 июля 2007 на Wayback Machine.
Литература
- Aldridge R. J., Briggs D. E. G., Smith M. P., Clarkson E. N. K. & Clark N. D. L. (1993). The anatomy of conodonts. Philosophical Transactions of the Royal Society of London, Series B 340: 405—421.
- Aldridge R. J. & Purnell M. A. (1996). The conodont controversies. Trends in Ecology and Evolution 11: 463—468.
- Donoghue P. C. J., Forey P. L. and Aldridge R. J. (2000). Conodont affinity and chordate phylogeny. Biological Reviews 75: 191—251.
- Janvier P. Euconodonta. The tree of life web project (1997). Дата обращения: 5 сентября 2007. Архивировано 23 марта 2012 года.
- Sweet, Walter. The Conodonta.
- Sweet W. C. and Donoghue P. C. J. (2001). Conodonts: past, present and future. Journal of Paleontology 75: 1174—1184.
Ссылки
- Mark Purnell. An oblique anterior view of a model of the apparatus of the Pennsylvanian conodont Idiognathodus. Архивировано 23 марта 2012 года.
- "The Jaws That Catch": an Introduction to the Conodonta (недоступная ссылка). Palæos. Дата обращения: 5 сентября 2007. Архивировано 13 марта 2006 года.
- Jim Davison. Ordovician conodonts (недоступная ссылка) (15 октября 2002). Дата обращения: 5 сентября 2007. Архивировано 11 марта 2006 года.
- Конодонты прятали острые зубы за примитивными дёснами.
| Таксономия | |
|---|---|
| В библиографических каталогах |