Ядра таламуса
Ядра таламуса — это группы нейронов таламуса, выделяемые на основании особенностей их анатомо-гистологической структуры и цитоархитектоники, а также паттерна связей с другими частями головного мозга.[1]
| Ядра таламуса | |
|---|---|
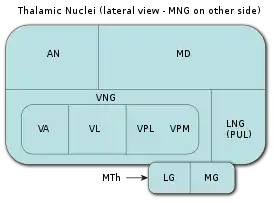 Список ядер таламуса: MNG = Срединная группа ядер AN = Передние ядра таламуса MD = Медиальное дорсальное ядро VNG = Вентральная группа ядер VA = Вентральное переднее ядро VL = Вентральное латеральное ядро VPL = Вентральное постеролатеральное ядро VPM = Вентральное постеромедиальное ядро LNG = Латеральная группа ядер PUL = Ядра подушки MTh = Метаталамус LG = Латеральное коленчатое тело MG = Медиальное коленчатое тело | |
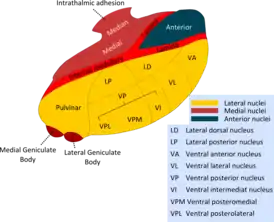 Ядра таламуса | |
| Каталоги | |
Все ядра таламуса можно подразделить на шесть групп: передние, медиальные, боковые (латеральные), ретикулярные, внутриламинарные ядра и ядра средней линии. Тонкий Y-образный слой миелинизированных нервных волокон, так называемая внутренняя медуллярная пластинка, разграничивает между собой переднюю, медиальную и боковую (латеральную) группы таламических ядер. У человека передняя и медиальная группы таламических ядер содержат всего по одному большому ядру, называемому соответственно передним дорсальным и медиальным дорсальным ядрами таламуса. Между тем боковая (латеральная) группа ядер таламуса человека подразделяется на дорсальный и вентральный уровни. Дорсальный уровень боковых (латеральных) ядер таламуса человека состоит из бокового дорсального, бокового заднего ядер и ядер подушки таламуса. Вентральный уровень боковых ядер таламуса человека, в свою очередь, состоит из вентрального переднего, вентрального бокового, вентрального задне-бокового и вентральных задних медиальных ядер.[1]
Боковая группа ядер таламуса покрыта другим тонким слоем миелинизированных нервных волокон — так называемой внешней медуллярной пластинкой. Между этим пучком нервных волокон и внутренней капсулой находится тонкий слой ГАМКергических нейронов, образующих ретикулярное ядро. Группа внутриламинарных (интраламинарных) ядер таламуса, среди которых самым крупным является так называемое центральное срединное ядро таламуса, как следует из её названия, располагается внутри Y-образной внутренней медуллярной пластинки.[1]
Наконец, последняя группа ядер таламуса, так называемые срединные ядра, или «ядра средней линии», покрывают собой часть медиальной поверхности обеих половинок таламуса, и примыкают к верхней боковой стенке третьего желудочка головного мозга. Эти ядра средней линии (срединные ядра) каждой из половинок таламуса могут соединяться между собой тонким и плоским серым пучком нервных волокон, так называемым межталамическим сращением. Выраженность межталамического сращения вариабельна не только у разных видов позвоночных, но даже у разных особей одного и того же вида (в том числе у разных людей), вплоть до её полного отсутствия. Отсутствие или слабая выраженность межталамического сращения практически не влияет на функциональность таламуса и является не патологией, а лишь анатомической особенностью (такой же, как наличие артерии Першерона).[1]
Функциональные различия ядер таламуса
Хотя анатомо-гистологическое подразделение ядер таламуса, безусловно, полезно, но большее значение для понимания его функционирования имеет их функциональная группировка, проводимая на основании типа их связей с другими частями мозга и типа передаваемой по этим связям и обрабатываемой этими ядрами информации. С этой точки зрения, среди ядер таламуса функционально выделяют первичные сенсорные и моторные ретрансляционные ядра, ядра ретрансляции ассоциативной информации, диффузно-проекционные ядра, и тормозное, ауторегулирующее активность таламуса, ретикулярное ядро, состоящее, в отличие от остальных ядер таламуса, из ингибирующих ГАМКергических нейронов, а не из возбуждающих глутаматергических.[1]
Первичные сенсорные и моторные ретрансляционные ядра
Первичные сенсорные и моторные ретрансляционные ядра таламуса передают информацию по своим проекциям в определённые специфические области сенсорной и моторной коры больших полушарий головного мозга, и получают от этих областей реципрокную обратную связь. Эта обратная связь, по-видимому, позволяет сенсорной и моторной коре больших полушарий мозга модулировать активность соответствующих таламических ядер и интенсивность получаемого ею входящего сигнала от этих ядер, а также работу соответствующих таламических фильтров.[1] Каждое конкретное первичное сенсорное или моторное ретрансляционное ядро таламуса получает и обрабатывает информацию только от одной определённой сенсорной системы или от одной определённой части двигательной системы. Так, например, латеральное коленчатое ядро таламуса получает зрительный сигнал от зрительного тракта, производит его простейшую первичную обработку, фильтрует от помех и затем передаёт обработанный и отфильтрованный зрительный сигнал первичной зрительной коре, расположенной в затылочной доле коры больших полушарий мозга.[1] Нейроны первичных сенсорных ретрансляционных ядер таламуса для ряда сенсорных систем формируют топографически организованные проекции в определённые специфические зоны коры больших полушарий (то есть, например, для первичной слуховой коры определённые частоты звуковых сигналов отображаются соответствующим ядром таламуса на определённые участки слуховой коры; то же самое верно для зрительной, соматосенсорной и висцеросенсорной систем, в которых отображается карта окружающего пространства или схема тела, соответственно). Аналогичную топографическую организацию проекций в моторную кору, отображающую схему тела, проявляют нейроны первичных моторных ретрансляционных ядер таламуса. Вместе с тем, некоторые первичные сенсорные и моторные зоны коры больших полушарий получают информацию более чем от одного первичного сенсорного или моторного ядра таламуса.[1]
Ядра ретрансляции ассоциативной информации
В отличие от первичных сенсорных и моторных ретрансляционных ядер таламуса, каждое из которых получает довольно простую и «сырую» (почти необработанную) информацию только от одного источника сенсорной или моторной информации, и передаёт её в довольно ограниченные по своим размерам области первичной сенсорной или моторной коры, ядра ретрансляции ассоциативной информации таламуса получают предварительно уже подвергшуюся высокой степени переработки информацию более чем из одного источника, ассоциируют одну информацию с другой, и передают её по своим проекциям в значительно более крупные по своим размерам области ассоциативной коры. Так, например, медиальное дорсальное ядро таламуса получает информацию одновременно от гипоталамуса и от миндалины, и связано реципрокными двусторонними связями с префронтальной корой, а также с определёнными областями премоторной и височной коры.[1]
Диффузно-проекционные ядра
В отличие от ретрансляционных ядер обоих типов (как первичных сенсорных и моторных ретрансляторов, так и ядер ретрансляции ассоциативной информации), диффузно-проекционные ядра получают менее специфическую входящую информацию, но зато одновременно от множества различных источников. Они также широковещательно, диффузно ретранслируют информацию по многообразным своим восходящим проекциям во многие различные области коры больших полушарий и в другие ядра таламуса, а по нисходящим проекциям — во множество различных подкорковых структур. Такой диффузный характер их проекций, широковещательный охват ретранслируемой по этим проекциям информации, а также неспецифичность получаемой ими входной информации, заставляет предполагать, что диффузно-проекционные ядра играют важную роль в регуляции общего уровня корковой и подкорковой возбудимости, уровня сознания, уровня активности и возбуждения ЦНС, концентрации внимания, и в регуляции смены состояний сна и бодрствования. И действительно, разрушение или повреждение этих ядер приводит в лёгких случаях к нарушениям концентрации внимания, к сонливости, а в более тяжёлых случаях — к летаргическому сну или перманентной коме. И наоборот, дегенеративные прионные изменения в них, такие, какие наблюдаются при фатальной семейной бессоннице — приводят к развитию упорной хронической, в пределе полной, бессонницы и в конечном итоге к смерти.[1]
Ауторегуляторное ретикулярное ядро
Ретикулярное ядро таламуса уникально тем, что оно, в отличие от всех других ядер таламуса, содержит не возбуждающие глутаматергические, а, наоборот, ингибирующие ГАМКергические нейроны. Ретикулярное ядро получает входящую информацию от коллатералей аксонов, которые взаимно (реципрокно) соединяют другие таламические ядра с корой больших полушарий головного мозга. Каждый нейрон ретикулярного ядра затем реципрокно проецирует свой единственный исходящий аксон именно в то ядро таламуса, от которого он получает входящую информацию. Такая схема связности нейронов ретикулярного ядра с остальными ядрами таламуса позволяет предполагать, что нейроны ретикулярного ядра таламуса осуществляют постоянный мониторинг и контроль уровня активности других таламических ядер и, косвенно, иннервируемых ими корковых областей, путём сэмплинга афферентной и эфферентной информации, поступающей от этих ядер таламуса в кору больших полушарий, и затем использует эту информацию для регулирования уровня активности соответствующих таламических ядер.[1]
Примечания
- Darlene Susan Melchitzky, David A. Lewis. 1.2 Functional Neuroanatomy // Kaplan and Sadock's Comprehensive Textbook of Psychiatry : [англ.] : in 2 vol. / под ред. Benjamin J. Sadock, Virginia A. Sadock, Pedro Ruiz. — 10-е изд. — Lippincott Williams & Wilkins, 2017. — Thalamus. — С. 158-170. — 12754 (эл.), 4997 (бум.) с. — ISBN 978-1451100471. — . — OCLC 949866139.