Верхнее двухолмие
Верхнее двухолмие (лат. Superior colliculus) — одна из основных структур среднего мозга хордовых. Оно обычно является частью его верхней дорсальной поверхности и вместе с нижним двухолмием (если оно имеется у данного вида) образует, соответственно, четверохолмие среднего мозга, выглядящее снаружи как четыре симметричных выступа («холмика») на задней (дорсальной) поверхности среднего мозга (см. рис.). Данное образование имеет несколько названий, при этом термины «тектум» (лат. «крыша») или «оптический тектум» употребляются обычно для немлекопитающих видов, а термин «верхнее двухолмие» — для млекопитающих, хотя и в этом случае прилагательное «тектальный» используется в литературе повсеместно. Структурно верхнее двухолмие состоит из нескольких слоёв, число которых различно у разных видов животных. При этом так называемые «верхние» слои связаны с обработкой информации от глаз и других сенсорных систем[2], а «глубинные» слои более связаны с моторными действиями, например, активацией движений глаз и другими движениями. Кроме этого имеются ещё и «средние» слои, содержащие мультимодальные сенсорные клетки, также обладающие моторными свойствами.
| Верхнее двухолмие, или верхние холмики четверохолмия | |
|---|---|
 Разрез сквозь верхние холмики четверохолмия, показывающий путь глазодвигательного нерва | |
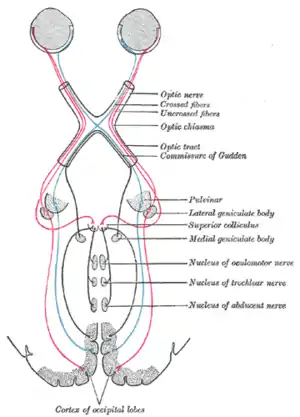 Схема, показывающая внутримозговые соединения зрительного нерва, а также зрительный тракт. Верхние холмики четверохолмия видны вблизи центра. | |
| Часть | Четверохолмия, которое, в свою очередь, является частью крыши мозга, а та — частью среднего мозга. |
| Каталоги | |
В общем и целом считается, что основная функция тектальной системы — непосредственные реакции, ориентированные на обусловленные точки в персональном эгоцентрическом пространстве (координаты которого задаются положением тела животного). Каждый из слоёв тектума содержит топологическую карту окружающего пространства в ретинотопических координатах, и активация нейронов в точке такой карты вызывает соответствующую реакцию, направленную на соответствующую точку пространства. Например, для приматов верхнее двухолмие изучалось преимущественно в связи с его ролью в управлении движениями глаз. Визуальное возбуждение от сетчатки, либо «команда» от неокортекса, создают определённый «очаг активности» на картах верхнего двухолмия, который, при достаточной его интенсивности, вызывает саккадное движение глаз. Но даже у приматов верхнее двухолмие также участвует в движениях общего целенаправленного поворота головы и протягивания рук[3], а также в сдвиге внимания, которое не связано ни с каким видимым движением[4]. У других животных тектум участвует в широком спектре двигательных реакций, включая поворот всего тела у крыс, рыб или птиц; выстреливание языком в сторону добычи у лягушек; целенаправленная атака (бросок) змей и т. д.
Для некоторых немлекопитающих видов, включая рыб и птиц, тектум является одной из самых крупных компонент их мозга. У млекопитающих, особенно у приматов, массивная экспансия церебральной коры мозга оставила тектуму (верхнему двухолмию) значительно меньшую долю в общем объёме мозга. Но даже у них он остаётся важной частью с функциями первичной интеграции информации, обеспечивающей движения глаз.
Эволюция и сравнительная анатомия

Оптический тектум — фундаментальная компонента мозга всех хордовых от миксинообразных до человека[5]. При этом некоторые аспекты его строения остаются постоянными, включая его общую структуру, состоящую из нескольких слоёв с массивным входом от оптического тракта к верхним слоям и мощным входом от других соматосенсорных систем к более глубоким слоям. Прочие аспекты строения тектума варьируют весьма значительно. Например общее число слоёв тектума изменяется от 3 у двоякодышащей рыбы протоптера до 15 у золотой рыбки [5]. Число типов клеток меняется от двух у двоякодышащей рыбы до 27 у домашнего воробья[6]. У миксин, миног и акул тектум является относительно маленькой структурой в мозге, однако у костистых рыб он значительно больше и иногда становится самой крупной частью мозга (см. прилагаемый рисунок мозга трески). Для амфибий, рептилий и особенно птиц это также очень существенная компонента мозга, но у млекопитающих его относительный объём эволюционно значительно уменьшился из-за массивного расширения коры головного мозга[6].
Миноги
Нервной системе миног было посвящено много исследований, поскольку они имеют относительно простой мозг и считается, что его структура во многом отражает структуру мозга наших древнейших позвоночных предков. Начиная с 1970-х годов Стен Гриллинер вместе с коллегами в Королевском Институте Стокгольма использовал миног в качестве модели для понимания основ моторного контроля для позвоночных. Они начали со спинного мозга и продвигались далее к мозгу[7]. В серии своих исследований они обнаружили, что нейронные цепи в спинном мозге способны генерировать ритмические моторные паттерны, которые обеспечивают плавательные движения, и что эти цепи контролируются специфическими локомоторными зонами в стволе и среднем мозге, а эти области, в свою очередь, контролируются более высокими структурами мозга, включая базальные ганглии и тектум. В работе, посвящённой тектуму миног, опубликованной в 2007 году[8], они обнаружили, что его электрическая стимуляция может вызвать движения глаз, боковое сворачивающиеся движение либо плавательные движения, и что тип, амплитуда и направление этих движений явно зависит от того места в тектуме, которое было простимулировано. Эти наблюдения были проинтерпретированы как согласующиеся с общим представлением о тектуме как о генераторе направленных локомоций для миног, как и для других видов позвоночных.
Летучие мыши
Хотя летучие мыши вовсе не слепы, они гораздо больше зависят от своей звуковой эхолокации, чем от зрения, во время своих ночных полётов и охоты на добычу. Поэтому весь их мозг очень чётко специализирован для этого процесса и часть таких специализаций наблюдается и в верхнем двухолмии[9]. У них нервные проекции от сетчатки глаз приходят только в тонкий слой сразу у поверхности двухолмия, а проекции от аудиальных зон гораздо более развиты, как и выходы в моторные области отвечающие за ориентацию ушей, головы или всего тела летучей мыши. Эхо, приходящее с различных направлений, активирует нейроны в разных местах уровней двухолмия[10], а стимуляция нейронов двухолмия влияет на звуковые импульсы, испускаемые летучей мышью. То есть имеется чёткое указание, что верхнее двухолмие выполняет в данном случае функции того же рода для аудиально направляемого поведения летучих мышей, что и для визуально направляемого поведения у других видов. При этом для особого вида крупных летучих мышей, называемых летучими лисицами, которые более полагаются на своё развитое зрение для навигации, их визуальные рецепторные поля в верхнем двухолмии формируют точно такие же ретинотопические карты, очень сходные с полями находимыми у кошек и приматов.
Внутреннее устройство
Внутренняя структура оптического тектума/верхнего двухолмия (ОТ/ВД) существенно различается у позвоночных разных видов. Но как общее правило можно указать на чёткое различие между верхними уровнями, которые получают проекции от визуальной системы и реагируют в основном на визуальные стимулы, и более глубокими уровнями, которые получают нейронные проекции различных типов и сами проецируются в различные моторные и связанные с ними структуры мозга. Различие между этими двумя областями всегда настолько чёткое и постоянное, что некоторые анатомы предлагают их рассматривать как отдельные структуры мозга.
Для млекопитающих большинство нейроанатомов выделяют семь слоёв или нейронных пластинок[11]. Первые три из них называются верхними слоями (superficial layers):
- Пластинка I или SZ (от stratum zonale (поверхностный слой)), это очень тонкий слой состоящий из небольших миелинированных аксонов вместе с остаточными и горизонтальными клетками.
- Пластинка II или SGS (от stratum griseum superficiale (верхний серый слой)), содержит множество нейронов различной формы и типов.
- Пластинка III или SO (от stratum opticum (оптический слой), состоит в основном из аксонов приходящих из оптического тракта.
Далее идут два промежуточных или средних слоя (intermediale layers):
- Пластинка IV или SGI (от stratum griseum intermediale (промежуточный серый слой) — самый толстый слой состоящий также из нейронов различного типа и размеров. Этот уровень по толщине часто равен всем остальным вместе взятым, поэтому в нём иногда ещё выделяют «верхние» («upper») и «нижние» («lower») слои (обозначены на рисунке как uSGI и lSGI соответственно).
- Пластинка V или SAI (от stratum album intermediale (промежуточный белый слой)), состоит в основном из аксонов от самых разных источников.
И наконец идут два нижних слоя (deep layers):
- Пластинка VI или SGP (от stratum griseum profundum (нижний серый слой)) состоит из плотно упакованных нейронов и аксонов между ними.
- Пластинка VII или SAP (от stratum album profundum (нижний белый слой)) находится точно над центральным серым телом (periaqueductal gray) и состоит только из аксонов.
Верхние слои ВД получают импульсы в основном от сетчатки глаз, визуальных зон коры и ещё двух связанных с тектумом структур, называемых «претектальная область», «претектум» (pretectum) и «парабигемиальные ядра» (parabigeminal nucleus). Проекции от сетчатки охватывают всю область верхних слоёв и являются формально билатеральными (от каждого глаза в обе половинки ВД), хотя контрлатеральные проекции (от противоположного глаза) значительно превосходят ипсилатеральные в приблизительном соотношении 95 % к 5 %. Кортикальные проекции приходят преимущественно от первичной визуальной коры (зона 17), вторичной визуальной коры (зоны 18 и 19) и фронтальных глазных полей (frontal eye fields — FEF). Ядра parabigeminal nucleus (претектум) также играют весьма значительную роль для выполнения тектальных функций, что будет описано далее.
В отличие от исключительно визуального доминирования в проекциях на верхние уровни тектума, его средние и нижние уровни имеют связи с очень разнообразным набором сенсорных и моторных структур мозга. Большинство остальных зон коры мозга шлют свои проекции именно к этим уровням, хотя связи от «ассоциативных» зон выглядят более значительно, чем связи от первичных сенсорных и моторных областей (нет ссылки на цитату). Количество связей от различных зон коры к этим уровням значительно варьируется для различных видов животных[12]. Ещё одна очень важная проекция приходит в средние уровни ВД от ретикулярного чёрного тела (substantia nigra pars reticulata) компоненты базальных ганглий. В ней используется тормозящий (подавляющий) нейротрансмиттер GABA и предполагается, что таким образом контролируется общий пропускной («gating») эффект для информации через верхнее двухолмие (от верхних слоёв к моторным средним и нижним). Кроме того промежуточные и нижние слои ОТ получают проекции от спинных ядер тройничного нерва (spinal trigeminal nucleus), которые передают соматосенсорную информацию от лица, а также проекции от гипоталамуса (hypothalamus), от субталамической неопределённой зоны (zona incerta), таламуса (thalamus), и нижнего двухолмия (inferior colliculus).
Кроме отличий во входящих связях верхние и нижние слои тектума также сильно различаются и в своих выходных проекциях. Для верхних слоёв один из важнейших выходов идёт в подушкообразное ядро (pulvinar) и латеральную среднюю зону таламуса, которая в свою очередь проецируется в зоны коры, связанные с контролем движений глаз. Кроме того, есть проекции от верхних уровней к претектальным ядрам (подробнее описанные далее) и латеральному коленчатому телу таламуса. Проекции от нижних слоёв тектума гораздо более разнообразны (см. например схему в начале статьи). Среди них есть два крупных нисходящих пути, идущих в ствол мозга и спинной мозг, а также множество восходящих проекций к различным сенсорным и моторным структурам, включая некоторые участвующие в инициации движений глаз.
Мозаичное строение
При более подробном изучении становится видно, что уровни двухолмия не являются однородными слоями, но организованы в отдельные колонки расположенные сотовым образом[13][14]. Наиболее явно колоночная структура проявляется при маркировании холинергических проекций от претектальных ядер, чьи терминали образуют сходные по размерам кластеры, простирающиеся от верхних до нижних слоёв тектума[15]. Однако и другие нейрохимические маркеры, включая рецепторы калретинина (calretinin), парвальбумина (parvalbumin), GAP-43, NMDA, а также карты связей с различными структурами в стволе и промежуточном мозге, показывают сходную неоднородность[16]. Общее число колонок в тектуме оценивается приблизительно в 100 штук[13]. Функциональное значение такой колоночной организации тектума до сих пор до конца не ясно, однако в недавних исследованиях появились свидетельства, что указанные выше холинергические связи являются частью рекуррентной цепочки связей, которые динамически выявляют абсолютных «победителей» в активации колонок тектума. Более подробно этот механизм описан далее.
Хотя у всех исследованных видов позвоночных, включая млекопитающих и прочих, была обнаружена описанная модуляризация тектума, тем не менее имеется несколько систематических различий в деталях их строения[15]. Для видов с равномерной сетчаткой (в основном для видов с боковым расположением глаз, таких как зайцы или лоси) разбиение на модули распространялось на все верхнее двухолмие. Однако для видов с центральным расположением зрительной ямки подразделение на модули не соблюдалось полностью в ростральной области ВД, которая как раз связана с этой частью сетчатки. Эти области ВД содержат множество нейронов «фиксации взгляда», которые постоянно посылают импульсы, когда глаза животного остаются зафиксированными в одном положении.
Связи с претектальными ядрами
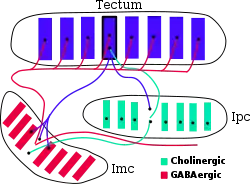
Оптический тектум очень тесно связан с близкой к нему структурой, называемой претектальные ядра (nucleus isthmii для немлекопитающих), которая недавно привлекла к себе пристальное внимание, поскольку появились новые данные о значительном вкладе этой структуры в исполнение «выделяющих» функций тектума[17]. Для млекопитающих, где оптический тектум называется верхним двухолмием, данная структура также имеет другое название — парабигемиальные ядра (parabigeminal nucleus). Тем не менее, это одна и та же структура, только с разными названиями. Эти ядра, в свою очередь, разделяют на две подструктуры — с крупными клетками (pars magnocellularis, Imc) и с «мелкими» клетками (pars parvocellularis — Ipc). Первую подструктуру (Imc) также ещё называют полумесячным ядром (pars semilunaris), поскольку оно выглядит как полумесяц на поперечных срезах среднего мозга.
Как показано на прилагаемой диаграмме, соединения между тектумом и претектальными ядрами Imc и Ipc имеют чёткий топографически согласованный порядок. Нейроны из верхних слоёв тектума проецируются в соответствующие точки Imc и Ipc. При этом их проекции в Ipc чётко сфокусированы, а проекции в Imc гораздо более «размыты». Ipc в свою очередь посылает чётко сфокусированные холинергические (возбуждающие) проекции и в Imc и обратно в тектум, где они образуют терминали по всей колонке тектума, от верха до низа. Напротив, Imc посылает GABA-проекции (тормозные) в Ipc и тектум, которые распространяются гораздо более широко по площади, захватывая большую часть их «карты». Таким образом, описанная здесь цепь нейронных связей тектум-Ipc-Imc обеспечивает рекуррентную обратную связь для импульсной активности в тектуме, что даёт в результате чётко сфокусированное возбуждение в колонке нейронов тектума вместе с обширным подавлением удалённых тектальных нейронов.
Функции
В истории исследований оптического тектума было несколько серьёзных сдвигов или этапов в понимании общей роли этой структуры. Примерно до 1970 года большинство исследований тектума проводилось на немлекопитающих (рыбах, лягушках, птицах) у которых тектум является доминирующей структурой получающей и обрабатывающей информацию от глаз. Тогда общим мнением было, что тектум является основным визуальным центром для мозга немлекопитающих и, соответственно, участвует в очень широком спектре поведения для подобных видов. В период с 1970-х до 1990-х исследование нейронных импульсов у млекопитающих, большей частью обезьян, было сосредоточено в основном на роли верхнего двухолмия в контроле движений глаз. И это направление исследований стало настолько доминировать в научной литературе, что сформировало мнение о верхнем двухолмии, как будто контроль движений глаз является его единственной основной функцией для млекопитающих. Этот взгляд до сих пор находит отражение даже во множестве современных учебников.
Однако в конце 1990-х годов в экспериментах, где голова животных не была жёстко закреплена, было ясно показано, что ВД на самом деле инициирует поворот взгляда, состоящий как правило и из поворота головы, и из собственно движения глаз (а не только последнего). Это открытие пробудило интерес к повторному исследованию всего спектра функций верхнего двухолмия и привело к изучению его мультисенсорной интеграции в самых различных ситуациях, для разных видов животных. Тем не менее, до сих пор роль ВД в контроле поворота глаз изучена и понимается гораздо глубже всех его остальных функций.
Поведенческие эксперименты показали, что ВД хотя и не является необходимой структурой для распознавания образов, но играет критическую роль в направлении поведения на определённый объект, и может обеспечивать эту функцию даже при отсутствии коры мозга.[18] Так кошки с обширным повреждением визуальной коры, хотя и не могли распознать объекты, тем не менее все равно могли следовать и ориентироваться на движущийся стимул, но гораздо медленнее чем обычно. Если при этом у них была удалена одна половинка ВД, то они постоянно кружили в сторону противоположную удалённой части ВД и компульсивно ориентировались на объекты расположенные в этой области. Однако им это не удавалось сделать с объектами в другой стороне от них. Этот дефицит функций постепенно уменьшался со временем, но никогда не исчезал полностью.
Движения глаз
Для приматов среди всех движений глаз можно выделить несколько типов: фиксация, при которой глаза направлены на неподвижный объект и движения глаз совершаются только для компенсации движений головы; плавное следование, при котором глаза постоянно следуют за движущимся предметом, саккады, при которых глаза очень быстро перепрыгивают с одного направления на другое; и наведение, когда глаза сдвигаются одновременно в противоположных направлениях осуществляя единое бинокулярное зрение объектов на определённом удалении. Верхнее двухолмие участвует во всех этих движениях, однако его роль в осуществлении саккадных движений изучена наиболее полно.
Каждая латеральная «половинка» верхнего двухолмия содержит в своих слоях двумерную карту, представляющую половину зрительного поля. При этом зрительная ямка сетчатки (fovea) — регион максимальной чувствительности глаза — представлена на ростральной части карты, а периферия зрения — на её каудальной части. Движения глаз вызываются активностью в глубоких слоях ВД. Во время фиксации взгляда нейроны в ростральной части ВД — зоне зрительной ямки — все время тонически активны. Во время плавного следования возбуждаются нейроны на небольшом расстоянии от ростральной части карты ВД, что приводит к небольшим движениям глаз. При саккадах возбуждаются нейроны в регионах представляющих точку, куда будет направлена саккада. И прямо перед её осуществлением активность этих нейронов быстро нарастает и уменьшается во всех остальных частях ВД. При этом, точка цели саккады представлена довольно широким «холмом» активности нейронов, который захватывает существенную часть карты двухолмия, а вершина этого холма как раз и обозначает целевую точку саккады.
Также ВД представляет на своих картах цель для поворота части или всего тела, но похоже, что при этом оно не задаёт, каким именно способом будет осуществлено это движение[19]. Последующая декомпозиция такого движения на поворот головы и на поворот глаз, а также конкретная траектория движения глаз во время саккады зависит от обработки сигналов от ВД в последующих моторных центрах, механизм которой сейчас не совсем понятен. Однако, независимо от того как инициируется или выполняется конкретное движение, ВД всегда кодирует его конечную цель в ретинотопических координатах. То есть паттерн активации ВД задаёт требуемый сдвиг от текущего направления глаз, независимо от их положения[20].
Существуют разные мнения насчёт того, участвует ли ВД непосредственно в управлении движениями глаз, либо же полностью оставляет это другим структурам, либо активно участвует только в исполнении саккад. В 1991 году Муньос (Munoz) с коллегами получили данные, что вышеописанный «холм» активности нейронов ВД постепенно продвигается во время исполнения саккады, постоянно отображая изменяющийся угол до её цели[21]. Однако сейчас доминирует точка зрения, что хотя «холм» и сдвигается немного во время саккады, однако это не происходит плавно и пропорционально, как предсказывала первоначальная гипотеза «движущегося холма»[22].
Исходящие от моторных слоёв ВД сигналы идут ко множеству ядер среднего и ствола мозга, где трансформируются из кодов «целевого отклонения», используемых в ВД, в коды частоты, используемые в моторных нейронах глаз. Дальнейшее движение глаз осуществляется шестью их мускулами, организованными в три ортогональные пары. Таким образом получается, что в самом конце движения глаз кодируются в обычных Декартовых координатах.
Хотя ВД получает очень мощные нейронные входы от сетчатки глаз, у приматов оно находится по большей части под контролем коры мозга, которая содержит в себе несколько областей связанных с заданием и коррекцией конкретных движений глаз[23]. Например, фронтальные глазные поля (frontal eye fields — FEF) — часть моторной коры, также участвует в генерации намеренных саккад глаз, а прилегающая к ним область, дополнительные глазные поля (additional eye fields — AEF) участвуют в организации групп саккадных движений в целостные последовательности. Теменные глазные поля (pariental eye fields — PEF), расположенные ещё ближе к затылку, участвуют в основном в рефлексивных саккадах, совершаемых в ответ на изменения в зрительном поле.
Визуальные сигналы приходят только в верхние слои ВД, тогда как его средние слои получают также аудиальные и соматосенсорные сигналы и подключены, в свою очередь, ко многим соматомоторным центрам и областям мозга. В настоящий момент полагают, что двухолмие в целом предназначено для ориентации головы и глаз на что-то, что было увидено или услышано.[4][24][25][26]
Различия
Летучие мыши
Как показали эксперименты со стимуляцией точек тектума летучих мышей, они могут вызывать не только целенаправленный поворот головы и глаз в заданном направлении, но и поворот ушей, а также, в некоторых случаях, целенаправленный эхолокационный импульс в этом направлении.
Змеи
Для змей, которые могут воспринимать инфракрасное (тепловое) излучение, таких как питоны и гремучие змеи, указанные нейронные сигналы передаются в средние слои их тектума естественно не через оптический тракт, а через тройничный нерв. Вся остальная обработка сигналов полностью аналогична случаю визуальной информации включая инициацию поворота головы и атаки (броска) [27].
См. также
Дополнительные рисунки
 Поверхностный срез ствола мозга. Вид сбоку.
Поверхностный срез ствола мозга. Вид сбоку. Срез ствола мозга. Вид сбоку.
Срез ствола мозга. Вид сбоку. Глубокий срез ствола мозга. Вид сбоку.
Глубокий срез ствола мозга. Вид сбоку.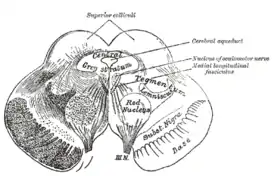 Поперечный разрез среднего мозга на уровне верхнего двухолмия.
Поперечный разрез среднего мозга на уровне верхнего двухолмия.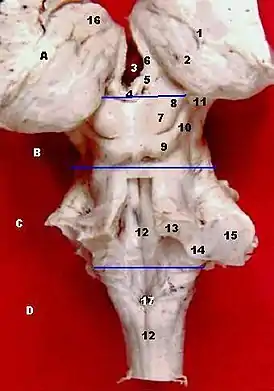 Ствол, средний мозг и таламус человека. Вид сзади.
Ствол, средний мозг и таламус человека. Вид сзади.
Примечания
- Superior colliculus // Foundational Model of Anatomy
- Wallace et al., 2005
- Lunenburger et al., 2001
- Kustov & Robinson, 1996
- Maximino, 2008
- Northcutt, 2002
- Grillner, 2003
- Saitoh et al., 2007
- Ulanovsky & Moss, 2008
- Valentine & Moss, 1997
- Huerta & Harting, 1984
- Clemo & Stein, 1984
- Chavalier & Mana, 2000
- Краснощекова 2007
- Illing, 1996
- Mana & Chevalier, 2001
- Li Xiao Wang, 2007
- Sprague, 1996
- Sparks & Gandhi, 2003
- Klier et al., 2001
- Munoz et al., 1991
- Soetedjo et al., 2002
- Pierrot-Deseilligny et al., 2003
- Klier et al., 2003
- Krauzlis et al., 2004
- Sparks, 1999
Ссылки
- Краснощекова Е.И. Модульная организация нервных центров. — 2-е изд. — СПб.: Изд-во СПбГУ, 2007. — С. 87 —102. — 130 с.
- Clemo, HR; Stein B. E. Topographic organization of somatosensory corticotectal influences in cat (англ.) // Journal of Neurophysiology : journal. — 1984. — Vol. 51, no. 5. — P. 843—858. — PMID 6726314.
- Chevalier, G; Mana S. Honeycomb-like structure of the intermediate layers of the rat superior colliculus, with additional observations in several other mammals: AChE patterning (англ.) // J Comp Neurol : journal. — 2000. — Vol. 419, no. 2. — P. 137—153. — doi:10.1002/(SICI)1096-9861(20000403)419:2<137::AID-CNE1>3.0.CO;2-6. — PMID 10722995.
- Dean, P; Redgrave P., Westby G. W. Event or emergency? Two response systems in the mammalian superior colliculus (англ.) // Trends Neurosci : journal. — 1989. — Vol. 12, no. 4. — P. 137—147. — doi:10.1016/0166-2236(89)90052-0. — PMID 2470171.
- Grillner, S. The motor infrastructure: from ion channels to neuronal networks (англ.) // Nat Rev Neurosci : journal. — 2003. — Vol. 4, no. 7. — P. 573—586. — doi:10.1038/nrn1137. — PMID 12838332.
- Hartline, PH; Kass L., Loop M. S. Merging of modalities in the optic tectum: infrared and visual integration in rattlesnakes (англ.) // Science : journal. — 1978. — Vol. 199, no. 4334. — P. 1225—1229. — doi:10.1126/science.628839. — PMID 628839.
- Huerta, MF; Harting J. K. Comparative Neurology of the Optic Tectum (англ.) / Vanegas H.. — New York: Plenum Press, 1984. — P. 687—773. — ISBN 9780306412363.
- Illing, R-B. The mosaic architecture of the superior colliculus (неопр.) // Prog Brain Res. — 1996. — Т. 112. — С. 17—34. — doi:10.1016/S0079-6123(08)63318-X. — PMID 8979818.
- King, AJ; Schnupp JWH, Carlile S., Smith A. L., Thompson I. D. The development of topographically-aligned maps of visual an auditory space in the superior colliculus (англ.) // Prog Brain Res : journal. — 1996. — Vol. 112. — P. 335—350. — doi:10.1016/S0079-6123(08)63340-3. — PMID 8979840.
- Klier, EM; Wang H., Crawford J. D. The superior colliculus encodes gaze commands in retinal coordinates (англ.) // Nat Neurosci : journal. — 2001. — Vol. 4, no. 6. — P. 627—632. — doi:10.1038/88450. — PMID 11369944.
- Klier, E; Wang H., Crawford D. Three-dimensional eye-head coordination is implemented downstream from the superior colliculus (англ.) // J Neurophysiol : journal. — 2003. — Vol. 89, no. 5. — P. 2839—2853. — doi:10.1152/jn.00763.2002. — PMID 12740415.
- Krauzlis, R; Liston D., Carello C. Target selection and the superior colliculus: goals, choices and hypotheses (англ.) // Vision Res : journal. — 2004. — Vol. 44, no. 12. — P. 1445—1451. — doi:10.1016/j.visres.2004.01.005. — PMID 15066403.
- Kustov, A; Robinson D. Shared neural control of attentional shifts and eye movements (англ.) // Nature : journal. — 1996. — Vol. 384, no. 6604. — P. 74—77. — doi:10.1038/384074a0. — PMID 8900281.
- Lane, RH; Allman J. M., Kaas J. H., Miezin F. M. The visuotopic organization of the superior colliculus of the owl monkey (Aotus trivirgatus) and the bush baby (Galago senegalensis) (англ.) // Brain Res : journal. — 1973. — Vol. 60, no. 2. — P. 335—349. — doi:10.1016/0006-8993(73)90794-4. — PMID 4202853.
- Lunenburger, L; Kleiser R., Stuphorn V., Miller L. E., Hoffmann K. P. A possible role of the superior colliculus in eye–hand coordination (англ.) // Prog Brain Res : journal. — 2001. — Vol. 134. — P. 109—125. — doi:10.1016/S0079-6123(01)34009-8. — PMID 11702538.
- Mana, S; Chevalier G. Honeycomb-like structure of the intermediate layers of the rat superior colliculus: afferent and efferent connections (англ.) // Neuroscience : journal. — Elsevier, 2001. — Vol. 103, no. 3. — P. 673—693. — doi:10.1016/S0306-4522(01)00026-4. — PMID 11274787.
- Maximino, C; Soares, Daphne. Evolutionary changes in the complexity of the tectum of nontetrapods: a cladistic approach (англ.) // PLOS One : journal / Soares, Daphne. — 2008. — Vol. 3, no. 10. — P. e385. — doi:10.1371/journal.pone.0003582. — PMID 18974789.
- Munoz, DP; Pélisson D., Guitton D. Movement of activity on the superior colliculus motor map during gaze shifts (англ.) // Science : journal. — 1991. — Vol. 251, no. 4999. — P. 1358—1360. — doi:10.1126/science.2003221. — PMID 2003221.
- Northcutt, R. G. Understanding vertebrate brain evolution (англ.) // Integrative and Comparative Biology : journal. — Oxford University Press, 2002. — Vol. 42, no. 4. — P. 743—746. — doi:10.1093/icb/42.4.743. — PMID 21708771.
- Pettigrew, J. D. Flying primates? Megabats have the advanced pathway from eye to midbrain (англ.) // Science : journal. — 1986. — Vol. 231, no. 4743. — P. 1304—1306. — doi:10.1126/science.3945827. — PMID 3945827.
- Pierrot-Deseilligny, C; Müri R. M., Ploner C. J., Gaymard B., Rivaud-Péchoux S. Cortical control of ocular saccades in humans: a model for motricity (англ.) // Prog Brain Res : journal. — 2003. — Vol. 142. — P. 3—17. — doi:10.1016/S0079-6123(03)42003-7. — PMID 12693251.
- Saitoh, K; Ménard A., Grillner S. Tectal control of locomotion, steering, and eye movements in lamprey (англ.) // J Neurophysiol : journal. — 2007. — Vol. 97, no. 4. — P. 3093—3108. — doi:10.1152/jn.00639.2006. — PMID 17303814.
- Soetedjo, R; Kaneko C. R., Fuchs A. F. Evidence against a moving hill in the superior colliculus during saccadic eye movements in the monkey (англ.) // J Neurophysiol : journal. — 2002. — Vol. 87, no. 6. — P. 2778—2789. — PMID 12037180.
- Sparks, D. L. Conceptual issues related to the role of the superior colliculus in the control of gaze (англ.) // Curr Op Neurobiol : journal. — 1999. — Vol. 6, no. 6. — P. 698—707. — doi:10.1016/S0959-4388(99)00039-2. — PMID 10607648.
- Sparks, DL; Gandhi N. J. Single-cell signals: an oculomotor perspective (неопр.) // Prog Brain Res. — 2003. — Т. 142. — С. 35—53. — doi:10.1016/S0079-6123(03)42005-0. — PMID 12693253.
- Sprague, J. M. Neural mechanisms of visual orienting responses (неопр.) // Prog Brain Res. — 1996. — Т. 112. — С. 1—15. — doi:10.1016/S0079-6123(08)63317-8. — PMID 8979817.
- Stein, BE; Clamman H. P. Control of pinna movements and sensorimotor register in cat superior colliculus (англ.) // Brain Behav Evol : journal. — 1981. — Vol. 19, no. 3—4. — P. 180—192. — doi:10.1159/000121641. — PMID 7326575.
- Ulanovsky, N; Moss C. F. What the bat's voice tells the bat's brain (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2008. — Vol. 105, no. 25. — P. 8491—8498. — doi:10.1073/pnas.0703550105. — PMID 18562301.
- Valentine, D; Moss C. F. Spatially selective auditory responses in the superior colliculus of the echolocating bat (англ.) // J Neurosci : journal. — 1997. — Vol. 17, no. 5. — P. 1720—1733. — PMID 9030631.
- Li, Da-Peng; Qian Xiao, Shu-Rong Wang. Feedforward Construction of the Receptive Field and Orientation Selectivity of Visual Neurons in the Pigeon (англ.) // Cerebral Cortex : journal. — 2007. — Vol. 17, no. 4. — P. 885—893. — doi:10.1093/cercor/bhk043.
- Valentine, Doreen E.; Shiva R. Sinha, Cynthia F. Moss. Orienting responses and vocalizations produced by microstimulation in the superior colliculus of the echolocating bat, Eptesicus fuscus (англ.) // J Comp Physiol : journal. — 2002. — Vol. 188, no. 2. — P. 89—108. — PMID 11919691. (недоступная ссылка)
- Wallace, MT; Meredith M. A., Stein B. E. Multisensory integration in the superior colliculus of the alert cat (англ.) // J Neurophysiol : journal. — 1998. — Vol. 80, no. 2. — P. 1006—1010. — PMID 9705489.