Клетка
Кле́тка (лат. cellula) — структурно-функциональная элементарная единица строения и жизнедеятельности всех организмов. Обладает собственным обменом веществ, способна к самовоспроизведению. Организм, состоящий из одной клетки, называется одноклеточным (многие простейшие и бактерии). Раздел биологии, занимающийся изучением строения и жизнедеятельности клеток, называется цитологией. Также принято говорить о биологии клетки, или клеточной биологии.
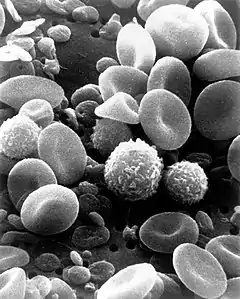
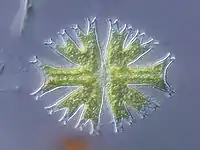
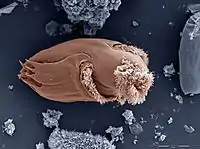
История открытия
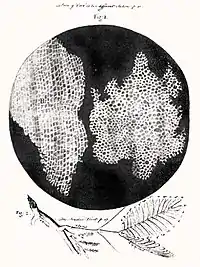
Первым человеком, увидевшим клетки, был английский учёный Роберт Гук (известный открытием закона Гука). В 1665 году, пытаясь понять, почему пробковое дерево хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему соты в ульях медоносных пчёл, которые он так и назвал — англ. cell («ячея»).
В 1675 году итальянский врач Марчелло Мальпиги подтвердил клеточное строение растений, а в 1681 году — английский ботаник Неемия Грю. О клетке стали говорить как о «пузырьке, наполненном питательным соком». В 1674 году голландский мастер Антоний ван Левенгук с помощью микроскопа впервые увидел в капле воды «зверьков» — движущихся живых организмов (инфузории, амёбы, бактерии). Также Левенгук впервые наблюдал животные клетки — эритроциты и сперматозоиды. Таким образом, к началу XVIII века учёные знали, что под большим увеличением растения имеют ячеистое строение, и видели некоторые организмы, которые позже получили название одноклеточных. В 1802—1808 годах французский исследователь Шарль-Франсуа Мирбель установил, что растения состоят из тканей, образованных клетками. Ж. Б. Ламарк в 1809 году распространил идею Мирбеля о клеточном строении и на животные организмы. В 1825 году чешский учёный Я. Пуркине открыл ядро яйцеклетки птиц, а в 1839 ввёл термин «протоплазма». В 1831 году английский ботаник Р. Броун впервые описал ядро растительной клетки, а в 1833 году установил, что ядро является обязательным органоидом клетки растения. С тех пор главным в организации клеток считается не оболочка, а содержимое.
Клеточная теория
Клеточная теория строения организмов была сформирована в 1839 году немецкими учёными, зоологом Теодором Шванном и ботаником М. Шлейденом, и включала в себя три положения. В 1858 году Рудольф Вирхов дополнил её ещё одним положением, однако в его идеях присутствовал ряд ошибок: так, он предполагал, что клетки слабо связаны друг с другом и существуют каждая «сама по себе». Лишь позднее удалось доказать целостность клеточной системы.
В 1878 году русским учёным И. Д. Чистяковым открыт митоз в растительных клетках; в 1878 году В. Флемминг и П. И. Перемежко обнаруживают митоз у животных. В 1882 году В. Флемминг наблюдает мейоз у животных клеток, а в 1888 году Э. Страсбургер — у растительных.
Клеточная теория является одной из основополагающих идей современной биологии, она стала неопровержимым доказательством единства всего живого и фундаментом для развития таких дисциплин, как эмбриология, гистология и физиология. На сегодняшний день теория содержит такие утверждения:
- Клетка — элементарная единица строения, функционирования, размножения и развития всех живых организмов. Вне клетки нет жизни.
- Клетка — целостная система, содержащая большое количество связанных друг с другом элементов — органелл.
- Клетки различных организмов похожи (гомологичны) по строению и основным свойствам и имеют общее происхождение.
- Увеличение количества клеток происходит путём их деления, после репликации их ДНК: клетка — от клетки.
- Многоклеточный организм — система из большого количества клеток, объединённых в системы тканей и органов, связанных между собой гуморальной и нервной регуляциями.
- Клетки многоклеточных организмов обладают одинаковым полным фондом генетического материала этого организма, всеми возможными потенциями для проявления этого материала, — но отличаются по уровню экспрессии (работы) отдельных генов, что приводит к их морфологическому и функциональному разнообразию — дифференцировке[2].
Количество и формулировки отдельных положений современной клеточной теории в разных источниках могут отличаться.
Методы исследования клеток
Впервые клетки удалось увидеть только после создания оптических (световых) микроскопов. С того времени микроскопия остается одним из важнейших методов исследования клеток. Световая микроскопия, несмотря на небольшое разрешение, позволяла наблюдать за живыми клетками. В XX веке была изобретена электронная микроскопия, которая позволила изучить ультраструктуру клеток.
Для изучения функций клеток и их частей используют разнообразные биохимические методы — как препаративные, например фракционирование методом дифференциального центрифугирования, так и аналитические. Для экспериментальных и практических целей используют методы клеточной инженерии. Все упомянутые методические подходы могут использоваться в сочетании с методами культуры клеток.
Оптическая микроскопия
В оптическом микроскопе увеличение объекта достигается благодаря серии линз, через которые проходит свет. Максимальное увеличение составляет более 1000 раз. Также важной характеристикой является разрешение — расстояние между двумя точками, которые ещё распознаются отдельно. Разрешение характеризует чёткость изображения. Эта величина ограничивается длиной световой волны, и даже при использовании самого коротковолнового света — ультрафиолетового — можно достичь разрешения только около 200 нм; такое разрешение было получено ещё в конце XIX века. Малейшие структуры, которые можно наблюдать под оптическим микроскопом, это митохондрии и бактерии. Их линейный размер составляет примерно 500 нм. Однако объекты размером меньше 200 нм видны в световом микроскопе, если они сами излучают свет. Эта особенность используется в флуоресцентной микроскопии, когда клеточные структуры или отдельные белки связываются со специальными флуоресцентными белками или антителами с флуоресцентными метками. На качество изображения, полученного с помощью оптического микроскопа, влияет также контрастность — её можно увеличить, используя различные методы окраски клеток. Для изучения живых клеток используют фазово-контрастную, дифференциальную интерференционно-контрастную и темнопольную микроскопию. Конфокальные микроскопы позволяют улучшить качество флуоресцентных изображений[3][4].
 Paramecium aurelia (светлопольная микроскопия) |
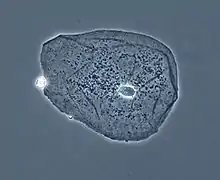 Клетки эпителия щеки (фазово-контрастная микроскопия) |
Nuclearia thermophila (дифференциальная интерференционно-контрастная микроскопия) |

Электронная микроскопия
 |
 | |
Эпителий трахеи (РЭМ) |
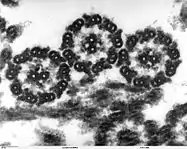
Поперечное сечение жгутиков хламидомонады (ПЭМ) |
В 30-х годах XX века был сконструирован электронный микроскоп, в котором вместо света через объект пропускается пучок электронов. Теоретический предел разрешения для современных электронных микроскопов составляет около 0,002 нм, однако из практических причин для биологических объектов достигается разрешение только около 2 нм. С помощью электронного микроскопа можно изучать ультраструктуру клеток. Различают два основных типа электронной микроскопии: сканирующую и трансмиссионную. Сканирующая (растровая) электронная микроскопия (РЭМ) используется для изучения поверхности объекта. Образцы зачастую покрывают тонкой плёнкой золота. РЭМ позволяет получать объёмные изображения. Трансмиссионная (просвечивающая) электронная микроскопия (ПЭМ) — используется для изучения внутреннего строения клетки. Пучок электронов пропускается через объект, предварительно обработанный тяжёлыми металлами, которые накапливаются в определённых структурах, увеличивая их электронную плотность. Электроны рассеиваются на участках клетки с большей электронной плотностью, в результате чего на изображениях эти области выглядят темнее[3][4].
Фракционирование клеток
Для установления функций отдельных компонентов клетки важно выделить их в чистом виде, чаще всего это делается с помощью метода дифференциального центрифугирования. Разработаны методики, позволяющие получить чистые фракции любых клеточных органелл. Получение фракций начинается с разрушения плазмалеммы и образования гомогената клеток. Гомогенат последовательно центрифугируется при различных скоростях, на первом этапе можно получить четыре фракции: (1) ядер и крупных обломков клеток, (2) митохондрий, пластид, лизосом и пероксисом, (3) микросом — пузырьков аппарата Гольджи и эндоплазматического ретикулума, (4) рибосом, в супернатанте останутся белки и более мелкие молекулы. Дальнейшее дифференциальное центрифугирование каждой из смешанных фракций позволяет получить чистые препараты органелл, к которым можно применять разнообразные биохимические и микроскопические методы[2].
Строение клеток
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
- прокариоты (доядерные) — более простые по строению, возникли в процессе эволюции раньше;
- эукариоты (ядерные) — более сложные, возникли позже. Клетки всех нынешних растений, животных, грибов и протистов - эукариотические.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам.
Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органеллы и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждая из органелл клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
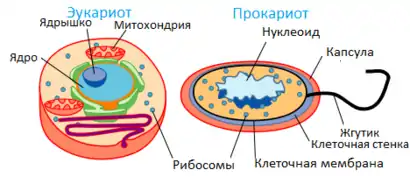
Прокариотическая клетка
Прокариоты (от др.-греч. πρό «перед, до» + κάρῠον «орех; ядро») — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
Эукариотическая клетка
Эукариоты, также эвкариоты (от др.-греч. εὖ «хорошо; полностью» + κάρῠον «орех; ядро») — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот есть система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.
Строение прокариотической клетки
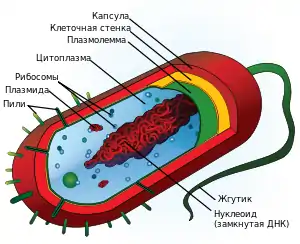
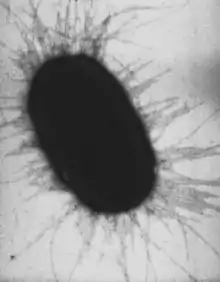
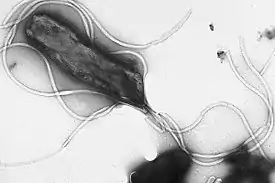
Клетки двух основных групп прокариот — бактерий и архей — похожи по структуре, характерными их признаками являются отсутствие ядра и мембранных органелл.
Основными компонентами прокариотической клетки являются:
- Клеточная стенка, которая окружает клетку извне, защищает её, придаёт устойчивую форму, предотвращающую от осмотического разрушения. У бактерий клеточная стенка состоит из пептидогликана (муреина), построенного из длинных полисахаридных цепей, соединённых между собой короткими пептидными перемычками. По строению клеточной стенки различают две группы бактерий:
- Грамположительные бактерии (например роды Staphylococcus, Bacillus, Lactobacillus[5]) — имеют более простую структуру клеточной стенки, состоящую почти исключительно из муреина;
- Грамотрицательные бактерии (например роды Salmonella, Escherichia, Azotobacter[5]) — клеточная стенка содержит меньше пептидогликана и имеет дополнительную внешнюю мембрану, которая состоит из фосфолипидов.
- Клеточная стенка архей не содержит муреина, а построена в основном из разнообразных белков и полисахаридов[4].
- Капсула — имеющаяся у некоторых бактерий слизистая оболочка, расположенная снаружи от клеточной стенки. Состоит в основном из разнообразных белков, углеводов и уроновых кислот. Капсулы защищают клетки от высыхания, могут помогать бактериям в колониях удерживаться вместе, а индивидуальным бактериям — прикрепляться к различным субстратам. Кроме этого, капсулы предоставляют клетке дополнительную защиту: например, капсулированные штаммы пневмококков свободно размножаются в организме и вызывают воспаление лёгких, тогда как некапсулированные быстро уничтожаются иммунной системой и являются абсолютно безвредными[5].
- Пили или ворсинки — тонкие волоскоподобные выросты, которые присутствуют на поверхности бактериальных клеток. Существуют различные типы пилей, из которых наиболее распространёнными являются:
- Фимбрии — пили, которые служат для прикрепления. Например, возбудитель гонореи — Neisseria gonorrhoeae — использует фимбрии для удержания на слизистой оболочке хозяина.
- Половые пили (F-пили) — задействованы в процессе конъюгации у бактерий[4].
- Жгутики — органеллы движения некоторых бактерий. Бактериальный жгутик построен значительно проще эукариотического, и он в 10 раз тоньше, не покрыт плазматической мембраной и состоит из одинаковых молекул белков, которые образуют цилиндр. В мембране жгутик закреплён при помощи базального тела[4].
- Плазматическая и внутренние мембраны. Общий принцип устройства клеточных мембран не отличается от эукариот, однако в химическом составе мембраны есть немало различий, в частности, в мембранах прокариот отсутствуют молекулы холестерина и некоторых липидов, присущих мембранам эукариот. Большинство прокариотических клеток (в отличие от эукариотических) не имеют внутренних мембран, которые разделяют цитоплазму на отдельные компартменты. Только у некоторых фотосинтетических и аэробных бактерий плазмалемма образует вгибание внутрь клетки, что выполняет соответствующие метаболические функции[4].
- Нуклеоид — не ограниченный мембранами участок цитоплазмы, в котором расположена кольцевая молекула ДНК — «бактериальная хромосома», где хранится весь генетический материал клетки[4].
- Плазмиды — небольшие дополнительные кольцевые молекулы ДНК, несущие обычно всего несколько генов. Плазмиды, в отличие от бактериальной хромосомы, не являются обязательным компонентом клетки. Обычно они придают бактерии определённые полезные для неё свойства, такие как устойчивость к антибиотикам, способность усваивать из среды определённые энергетические субстраты, способность инициировать половой процесс и т. д.[4][5]
- Рибосомы прокариот, как и у всех других живых организмов, отвечают за осуществление процесса трансляции (одного из этапов биосинтеза белка). Однако бактериальные рибосомы несколько меньше, чем эукариотические (коэффициенты седиментации 70S и 80S соответственно), и имеют другой состав белков и РНК. Из-за этого бактерии, в отличие от эукариот, чувствительны к таким антибиотикам, как эритромицин и тетрациклин, которые избирательно действуют на 70S-рибосомы[5].
- Эндоспоры — окружённые плотной оболочкой структуры, содержащие ДНК бактерии и обеспечивающее выживание в неблагоприятных условиях. К образованию эндоспор способны лишь некоторые виды прокариот, например представители родов Clostridium (C. tetani — возбудитель столбняка, C. botulinum — возбудитель ботулизма, C. perfringens — возбудитель газовой гангрены и т. п.) и Bacillus (в частности B. anthracis — возбудитель сибирской язвы). Для образования эндоспоры клетка реплицирует свою ДНК и окружает копию плотной оболочкой, из созданной структуры удаляется избыток воды, и в ней замедляется метаболизм[4]. Споры бактерий могут выдерживать довольно жёсткие условия среды, такие как длительное высушивание, кипячение, коротковолновое облучение и др.[5]
Строение эукариотической клетки

Поверхностный комплекс животной клетки

Состоит из гликокаликса, плазмалеммы и расположенного под ней кортикального слоя цитоплазмы. Плазматическая мембрана называется также плазмалеммой, наружной клеточной мембраной, цитолеммой и т. д. Её толщина — около 10 нанометров. Обеспечивает разграничение клетки и внешней среды, а также пропускание внутрь и наружу некоторых веществ.
На сохранение целостности своей мембраны клетка не тратит энергии: молекулы удерживаются по тому же принципу, по которому удерживаются вместе молекулы жира — гидрофобным частям молекул термодинамически выгоднее располагаться в непосредственной близости друг к другу. Гликокаликс представляет собой «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов, гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции.
Плазматическая мембрана животных клеток в основном состоит из фосфолипидов и липопротеидов со вкрапленными в неё молекулами белков, в частности, поверхностных антигенов и рецепторов.
В кортикальном (прилегающем к плазматической мембране) слое цитоплазмы находятся специфические элементы цитоскелета — упорядоченные определённым образом актиновые микрофиламенты. Основной и самой важной функцией кортикального слоя (кортекса) являются псевдоподиальные реакции: выбрасывание, прикрепление и сокращение псевдоподий. При этом микрофиламенты перестраиваются, удлиняются или укорачиваются. От структуры цитоскелета кортикального слоя зависит также форма клетки (например, наличие микроворсинок).
Структура цитоплазмы
Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органеллы. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами», и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
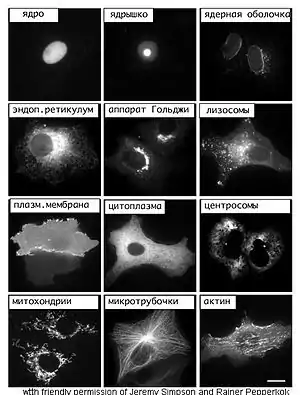
Рибосомы
Рибосомы — органоиды, необходимые клетке для синтеза белка. Их размер составляет примерно 20—30 нм. В клетке их насчитывается несколько миллионов. Рибосомы образованы из двух субъединиц: большой и малой, состоящих из четырёх молекул РНК и нескольких молекул белков. У эукариотических клеток рибосомы встречаются не только в цитоплазме, но и в митохондриях и хлоропластах. Рибосомы формируются в области ядрышек, а затем через ядерные поры выходят в цитоплазму.
Эндоплазматический ретикулум
В эукариотической клетке существует система переходящих друг в друга мембранных отсеков (трубочек, мешочков и плоских цистерн разных размеров), которая называется эндоплазматическим ретикулумом (или эндоплазматическая сеть, ЭПР или ЭПС). Ту часть ЭПР, к мембранам которого прикреплены рибосомы, относят к гранулярному (или шероховатому) эндоплазматическому ретикулуму, на его мембранах происходит синтез белков. Те компартменты, на стенках которых нет рибосом, относят к агранулярному (или гладкому) ЭПР, принимающему участие в синтезе липидов. Внутренние пространства гладкого и гранулярного ЭПС не изолированы, а переходят друг в друга и сообщаются с просветом ядерной оболочки.
Аппарат Гольджи
Аппарат Гольджи представляет собой стопку плоских мембранных цистерн, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. Аппарат Гольджи асимметричен — цистерны, располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от эндоплазматического ретикулума. По-видимому, при помощи таких же пузырьков происходит дальнейшее перемещение созревающих белков от одной цистерны к другой. В конце концов от противоположного конца органеллы (транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
Ядро
Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки — интроны), после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками.
Компартмент для ядра — кариотека — образован за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов ядерной оболочки. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой.
Лизосомы
Лизосома — небольшое тельце, ограниченное от цитоплазмы одинарной мембраной и имеющее вид пузырька. В ней находятся литические ферменты, способные расщепить практически любые природные органические соединения. Основная функция — автолиз — то есть расщепление отдельных органоидов, участков цитоплазмы клетки.
Цитоскелет
К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные филаменты. Микротрубочки принимают участие в транспорте органелл, входят в состав органоидов движения — ресничек и жгутиков, характерных для некоторых клеток (например, инфузорий, сперматозоидов), из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, по-видимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка.
Центриоли
Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет, за исключением низших водорослей). Центриоль представляет собой цилиндр, боковая поверхность которого образована девятью наборами микротрубочек. Количество микротрубочек в наборе может колебаться для разных организмов от 1 до 3.
Вокруг центриолей находится так называемый центр организации цитоскелета, район, в котором группируются минус-концы микротрубочек клетки.
Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путём синтеза новой структуры, перпендикулярной существующей.
Центриоли, по-видимому, гомологичны базальным телам жгутиков и ресничек.
Митохондрии
Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. Дыхание (поглощение кислорода и выделение углекислого газа) происходит также за счёт энзиматических систем митохондрий.
Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами, наружной и внутренней, между которыми располагается межмембранное пространство. Внутренняя мембрана митохондрии образует складки — кристы, на которых размещаются ферменты, ускоряющие реакции окисления жиров и углеводов. В матриксе содержатся различные ферменты, принимающие участие в дыхании и синтезе АТФ. Центральное значение для синтеза АТФ имеет водородный потенциал внутренней мембраны митохондрии.
Митохондрии имеют свой собственный ДНК-геном и прокариотические рибосомы, что, безусловно, указывает на симбиотическое происхождение этих органелл. В ДНК митохондрий закодированы совсем не все митохондриальные белки, большая часть генов митохондриальных белков находятся в ядерном геноме, а соответствующие им продукты синтезируются в цитоплазме, а затем транспортируются в митохондрии. Геномы митохондрий отличаются по размерам: например геном человеческих митохондрий содержит всего 13 генов. Самое большое число митохондриальных генов (97) из изученных организмов имеет простейшее Reclinomonas americana.
Сравнение прокариотической и эукариотической клеток
Наиболее важным отличием эукариот от прокариот долгое время считалось наличие оформленного ядра и мембранных органоидов. Однако к 1970—1980-м гг. стало ясно, что это лишь следствие более глубинных различий в организации цитоскелета. Некоторое время считалось, что цитоскелет свойственен только эукариотам, но в середине 1990-х гг. белки, гомологичные основным белкам цитоскелета эукариот, были обнаружены и у бактерий.
Именно наличие специфическим образом устроенного цитоскелета позволяет эукариотам создать систему подвижных внутренних мембранных органоидов. Кроме того, цитоскелет позволяет осуществлять эндо- и экзоцитоз (как предполагается, именно благодаря эндоцитозу в эукариотных клетках появились внутриклеточные симбионты, в том числе митохондрии и пластиды). Другая важнейшая функция цитоскелета эукариот — обеспечение деления ядра (митоз и мейоз) и тела (цитотомия) эукариотной клетки (деление прокариотических клеток организовано проще). Различия в строении цитоскелета объясняют и другие отличия про- и эукариот — например, постоянство и простоту форм прокариотических клеток и значительное разнообразие формы и способность к её изменению у эукариотических, а также относительно большие размеры последних. Так, размеры прокариотических клеток составляют в среднем 0,5—5 мкм, размеры эукариотических — в среднем от 10 до 50 мкм. Кроме того, только среди эукариот попадаются поистине гигантские клетки, такие как массивные яйцеклетки акул или страусов (в птичьем яйце весь желток — это одна огромная яйцеклетка), нейроны крупных млекопитающих, отростки которых, укреплённые цитоскелетом, могут достигать десятков сантиметров в длину.
Сравнительная характеристика клеток эукариот и прокариот[5] | ||
| Признак | Прокариоты | Эукариоты |
| Размеры клеток | Средний диаметр 0,5—10 мкм | Средний диаметр 10—100 мкм |
Организация генетического материала | ||
| Форма, количество и расположение молекул ДНК | Обычно имеется одна кольцевая молекула ДНК, размещённая в цитоплазме | Обычно есть несколько линейных молекул ДНК — хромосом, локализованных в ядре |
| Компактизация ДНК | У бактерий ДНК компактизируется без участия гистонов[6]. У архей ДНК ассоциирована с белками-гистонами[7] | Имеется хроматин: ДНК компактизируется в комплексе с белками-гистонами[6]. |
| Организация генома | У бактерий экономный геном: отсутствуют интроны и большие некодирующие участки[8]. Гены объединены в опероны[6]. У архей имеются интронные участки особой структуры[9]. | Большей частью геном не экономный: имеется экзон-интронная организация генов, большие участки некодирующей ДНК[8]. Гены не объединены в опероны[6]. |
Деление | ||
| Тип деления | Простое бинарное деление | Мейоз или митоз |
| Образование веретена деления | Веретено деления не образуется | Веретено деления образуется |
Органеллы | ||
| Тип рибосом | 70S-рибосомы | 80S-рибосомы |
| Наличие мембранных органелл | Окружённые мембранами органеллы отсутствуют, иногда плазмалемма образует выпячивание внутрь клетки | Имеется большое количество одномембранных и двумембранных органелл |
| Тип жгутика | Жгутик простой, не содержит микротрубочки, не окружён мембраной, диаметр около 20 нм | Жгутики состоят из микротрубочек, расположенных по принципу «9+2», окружены плазматической мембраной, диаметр около 200 нм |
Анаплазия
Разрушение клеточной структуры (например, при злокачественных опухолях) носит название анаплазии.
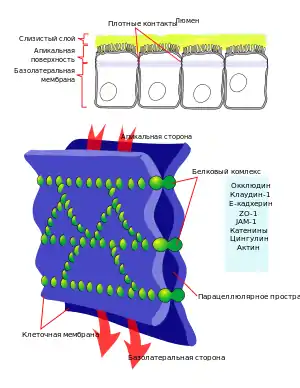
Межклеточные контакты
У высших животных и растений клетки объединены в ткани и органы, в составе которых они взаимодействуют между собой, в частности, благодаря прямым физическим контактам. В растительных тканях отдельные клетки соединяются между собой с помощью плазмодесм, а животные образуют различные типы клеточных контактов.
Плазмодесмы растений — это тонкие цитоплазматические каналы, которые проходят через клеточные стенки соседних клеток, соединяя их между собой. Полость плазмодесм устлана плазмалеммой. Совокупность всех клеток, объединённых плазмодесмами, называется симпластом, между ними возможен регулируемый транспорт веществ.
Межклеточные контакты позвоночных животных на основе строения и функций разделяют на три основных типа: якорные (англ. anchoring junctions), включающие адгезионные контакты и десмосомы, плотные или изоляционные (англ. tight junction) и щелевые или коммуникационные (англ. gap junction). Кроме того, некоторые особые виды соединений между клетками, такие как химические синапсы нервной системы и иммунологические синапсы (между T-лимфоцитами и антигенпрезентирующими клетками), объединяют по функциональному признаку в отдельную группу: контакты, которые передают сигналы, (англ. signal-relaying junction). Однако в межклеточном сигнализировании могут участвовать и якорные, щелевые и плотные контакты[3].
| Основные характеристики межклеточных контактов позвоночных животных[3] | ||
|---|---|---|
| Якорные контакты | Плотные контакты | Щелевые контакты |
 Адгезионный контакт |
 Плотный контакт |
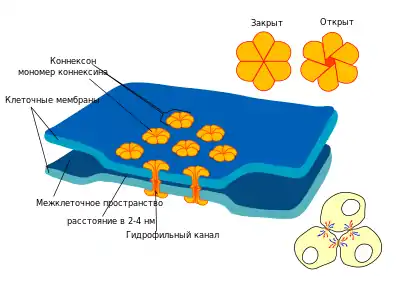 Щелевой контакт |
| Якорные контакты физически соединяют клетки между собой, обеспечивают целостность и прочность тканей, в частности эпителиальных и мышечных. При образовании контактов этого типа элементы цитоскелета соседних клеток как бы объединяются в единую структуру: с помощью специальных якорных белков они прикрепляются к внутриклеточной части белков кадгенринов, проходящих через плазматическую мембрану, и в межклеточном пространстве прикрепляются к кадгеринам соседних клеток. Различают два основных типа якорных контактов: адгезионные, объединяющие микрофиламенты соседних клеток; и десмосомы, в образовании которых принимают участие промежуточные филаменты. | Плотные (изоляционные) контакты обеспечивают максимальное сближение мембран соседних клеток, между которыми остается промежуток в 2-3 нм. Этот тип контактов чаще всего возникает в эпителии. Плотные контакты образуют непрерывные пояса вокруг каждой клетки, крепко прижимая их друг к другу и предотвращая протекание межклеточной жидкости между ними. Такие контакты необходимы, в частности, для обеспечения водонепроницаемости кожи. В формировании тесных контактов принимают участие белки окклюдины, клаудины и другие. | Щелевые (коммуникационные) контакты — это небольшие участки, на которых плазмалеммы соседних клеток приближены друг к другу на расстояние 2-4 нм и пронизаны белковыми комплексами — коннексонами. Каждый коннексон состоит из шести трансмембранных белков коннексинов, которые окружают небольшие гидрофильные поры диаметром в 1,5 нм. Через эти каналы от одной клетки к другой могут проходить ионы и другие небольшие гидрофильные молекулы. Таким образом происходит общение между соседними клетками. Щелевые контакты характерны для большинства тканей животного организма: в частности, эпителиальной, соединительной, сердечной мышцы, нервной (где формируют электрические синапсы) и др. |
Клеточный цикл
Деление клетки

Деление эукариотических клеток
Амито́з — прямое деление клетки, происходит в соматических клетках эукариот реже, чем митоз. В большинстве случаев амитоз наблюдается в клетках со сниженной митотической активностью: это стареющие или патологически изменённые клетки, часто обречённые на гибель (клетки зародышевых оболочек млекопитающих, опухолевые клетки и другие). При амитозе морфологически сохраняется интерфазное состояние ядра, хорошо видны ядрышко и ядерная оболочка. Репликация ДНК отсутствует. Спирализация хроматина не происходит, хромосомы не выявляются. Клетка сохраняет свойственную ей функциональную активность, которая почти полностью исчезает при митозе. Таково, например, деление макронуклеусов многих инфузорий, где без образования веретена происходит сегрегация коротких фрагментов хромосом. При амитозе делится только ядро, причём без образования веретена деления, поэтому наследственный материал распределяется случайным образом. Отсутствие цитокинеза приводит к образованию двуядерных клеток, которые в дальнейшем не способны вступать в нормальный митотический цикл. При повторных амитозах могут образовываться многоядерные клетки.
Мито́з (от греч. μιτος — нить) — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических клеток, один из фундаментальных процессов онтогенеза. Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяции тканевых клеток. Биологическое значение митоза заключается в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений[10]. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений[11]. На основании морфологических особенностей митоз условно подразделяется на:
Продолжительность митоза в среднем составляет 1—2 часа[10][12]. В клетках животных митоз, как правило, длится 30—60 минут, а в растительных — 2—3 часа[13]. Клетки человека за 70 лет суммарно претерпевают порядка 1014 делений[14].
Мейоз (от др.-греч. μείωσις — «уменьшение») или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток или гамет из недифференцированных стволовых. Уменьшение числа хромосом в результате мейоза в жизненном цикле ведёт к переходу от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса. В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма. Этот же механизм лежит в основе стерильности межвидовых гибридов. Определённые ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации).
Деление прокариотических клеток
Прокариоти́ческие клетки делятся надвое. Сначала клетка удлиняется, в ней образуется поперечная перегородка. На завершающем этапе дочерние клетки расходятся. Отличительной чертой деления прокариотических клеток является непосредственное участие реплицированной ДНК в процессе деления[15]. Обычно прокариотические клетки делятся с образованием двух одинаковых по размеру дочерних клеток, поэтому этот процесс ещё иногда называют бинарным делением. В связи с тем, что в подавляющем большинстве случаев прокариотические клетки имеют клеточную стенку, бинарное деление сопровождается образованием септы — перегородки между дочерними клетками, которая затем расслаивается посередине. Процесс деления прокариотической клетки подробно изучен на примере Escherichia coli[16].
Дифференцировка клеток многоклеточного организма
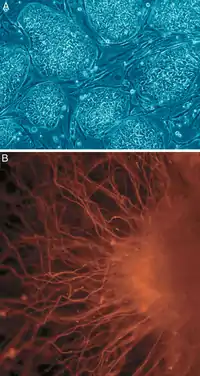
Многоклеточные организмы состоят из клеток, которые в той или иной степени отличаются по строению и функциям, например у взрослого человека около 230 различных типов клеток[17]. Все они являются потомками одной клетки — зиготы (в случае полового размножения) — и приобретают различия в результате процесса дифференцировки. Дифференцировка в подавляющем большинстве случаев не сопровождается изменением наследственной информации клетки, а обеспечивается лишь путём регуляции активности генов, специфический характер экспрессии генов наследуется во время деления материнской клетки обычно благодаря эпигенетическим механизмам. Однако есть исключения: например, при образовании клеток специфической иммунной системы позвоночных происходит перестройка некоторых генов, эритроциты млекопитающих теряют всю наследственную информацию, а половые клетки — её половину.
Различия между клетками на первых этапах эмбрионального развития появляются, во-первых, вследствие неоднородности цитоплазмы оплодотворённой яйцеклетки, из-за чего во время процесса дробления образуются клетки, различающиеся по содержанию определённых белков и РНК; во-вторых, важную роль играет микроокружение клетки — её контакты с другими клетками и средой.
Подвергаясь дифференцировке, клетки теряют свои потенции, то есть способность давать начало клеткам других типов. Из тотипотентных клеток, к которым относится, в частности, зигота, может образоваться целостный организм. Плюрипотентные клетки (например, клетки бластоцисты) имеют возможность дифференцироваться в любой тип клеток организма, но из них не могут развиться внезародышевые ткани, а значит и новая особь. Клетки, которые способны дать начало только ограниченному количеству других тканей, называются мультипотентными (стволовые клетки взрослого человека), а те, которые могут воспроизводить только себе подобных — унипотентными. Многие из окончательно дифференцированных клеток (например нейроны, эритроциты) полностью теряют способность к делению и выходят из клеточного цикла[3].
В некоторых случаях дифференцировка может быть обратной, противоположный ей процесс называется дедифференцировкой. Он характерен для процессов регенерации. С некоторыми оговорками к явлению дедифференцировки можно отнести опухолевую трансформацию клеток[18].
Клеточная смерть
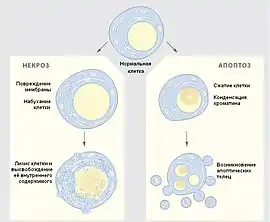
Одноклеточные организмы в некотором смысле можно считать «бессмертными», поскольку, за исключением случаев повреждения или голодания, они не умирают, а делятся, в результате чего образуется два новых организма. Зато все клетки многоклеточных организмов (кроме гамет) обречены на гибель, но умирают они не только в случае смерти всей особи — этот процесс происходит постоянно.
Смерть некоторых клеток необходима во время эмбрионального развития, клетки продолжают умирать и у взрослых организмов, например, в костном мозге и кишечнике человека ежечасно гибнут миллиарды клеток. Из-за физиологических условий происходит «запрограммированная клеточная смерть», другими словами клетки «совершают суицид». Наиболее распространённым, однако не единственным, путём клеточного самоуничтожения является апоптоз. Основными признаками апоптоза является фрагментация ДНК, распад клетки на апоптические тельца — везикулы, окружённые мембранами. На их поверхности расположены особые молекулы, которые побуждают соседние клетки и макрофаги фагоцитовать их таким образом, что процесс не сопровождается воспалением. Апоптоз является энергозависимым процессом и требует использования АТФ. Этот путь клеточной смерти важен не только для развития организма, нормального функционирования иммунной системы, но также и для защиты особи от повреждённых клеток, которые могут стать на путь злокачественной трансформации, и от вирусных инфекций[19].
Физическое или химическое повреждение клеток, а также недостаток источников энергии и кислорода, может привести к другой смерти — некротической. Некроз, в отличие от апоптоза, — пассивный процесс, он часто сопровождается разрывом плазмалеммы и утечкой цитоплазмы. Некроз почти всегда вызывает воспаление окружающих тканей. В последнее время исследуется механизм запрограммированного некроза как возможной противовирусной и противоопухолевой защиты[19].
При условии длительного недостатка АТФ в клетке она не сразу погибает путём некроза, а во многих случаях становится на путь аутофагии — процесса, который позволяет ей ещё некоторое время оставаться жизнеспособной. При аутофагии (буквально «самопоедание») обмен веществ переключается в сторону активного катаболизма, при этом отдельные органеллы окружаются двойными мембранами, образуются так называемые аутофагосомы, сливающиеся с лизосомами, где происходит переваривание органических веществ. Если голодовка продолжается и после того, как большинство органелл уже «съедено», клетка погибает путём некроза. Некоторые авторы считают, что при определённых условиях автофагия может быть отдельным типом клеточной смерти[19].
Отдельным типом клеточной смерти является нетоз — процесс гибели нейтрофилов, одного из типов иммунных клеток. Нейтрофилы при чрезмерном количестве вокруг патогенов могут выбрасывать сети нуклеиновых кислот из своего ядра. Патогены запутываются в этих сетях и нейтрализуются. Таким образом, нетоз можно считать "клеточным самопожертвованием". В последние годы, однако, нетоз был разделен на "витальный", при котором нейтрофил не погибает, и "суицидальный", при котором он погибает. Также описаны негативные эффекты нетоза на организм и его роль в патогенезе инфекционных, воспалительных и тромботических заболеваний.
Эволюция клеток
Доподлинно неизвестно, когда на Земле появилась первая клетка и каким путём она возникла. Наиболее ранние вероятные ископаемые остатки клеток, приблизительный возраст которых оценен в 3,49 млрд лет, найдены на востоке Пилбары (Австралия), хотя биогенность их происхождения была поставлена под сомнение. О существовании жизни в раннем архее свидетельствуют также строматолиты того же периода[20][21].
Возникновению первых клеток должно было предшествовать накопление органических веществ в среде и появление определённой формы пребиотического метаболизма. Протоклетки содержали как минимум два обязательных элемента: наследственную информацию в виде молекул, способных к саморепликации, и определённого рода оболочку, которая ограждала внутреннее содержимое первых клеток от окружающей среды. Наиболее вероятным кандидатом на роль саморепликативных молекул является РНК, поскольку она может одновременно выступать и носителем наследственной информации, и катализатором; кроме того, РНК, в отличие от ДНК, самодостаточна для осуществления биосинтеза белков[21][22].
Неизвестно также, из каких веществ были построены мембраны первых клеток, однако, вполне вероятно, это могли быть простые амфифильные соединения, такие как соли жирных кислот, способные самоорганизовываться в липосомы, которые могут проходить циклы роста и деления. Жирные кислоты были синтезированы во многих экспериментах по воспроизведению пребиотических условий, также они были найдены в метеоритах[22][23]. Считается, что первые живые клетки были гетеротрофными.
Возникновение эукариотических клеток
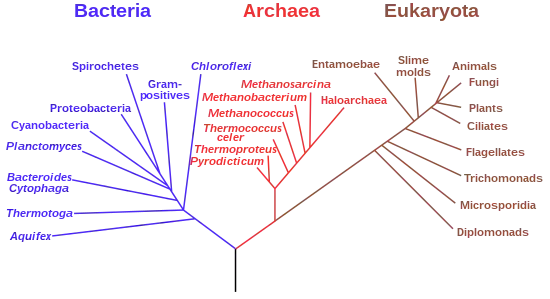
Данные секвенирования рРНК позволили построить универсальное дерево жизни, в котором последний универсальный общий предок дал начало двум ветвям эволюции: эубактериям и кладе Neomura, последняя из которых в свою очередь разделилась на две ветви: археи и эукариоты[24]. В эволюции эукариот, вероятно, большую роль сыграл эндосимбиоз — считается, что именно таким методом клетки ядерных получили митохондрии, а позже — и хлоропласты[25].
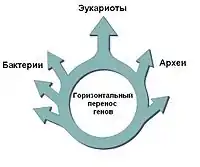
Эукариоты имеют много общих генов как с эубактериями, так с археями; некоторые учёные считают, что они возникли в результате слияния геномов этих двух групп организмов, что могло произойти в результате эндосимбиоза. Из-за этого, вместо «дерева жизни», предлагается использовать «круг жизни»[26]. Другие же исследователи, отмечая важность интенсивного горизонтального переноса между предками эукариот, бактерий и археобактерий, предлагают отображать филогенетические связи между ними с помощью «сетки жизни»[27].
Химическая организация клетки
См. также
Примечания
- Hooke, Robert (1635—1703). Micrographia, Observation XVIII.
- Ченцов Ю. С. Введение в клеточною биологию: Учебник для вузов. — 4-е изд. перераб. и доп./ Ю. С. Ченцов. — М.: ИКЦ «Академкнига», 2004. — 495 с.: ил. ISBN 5-94628-105-4
- Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. Molecular Biology of the Cell (англ.). — 5th. — Garland Science, 2007. — ISBN 978-0-8153-4105-5.
- Campbell N. A., Reece J. B. Biology (англ.). — 8th. — Benjamin Cammings, 2008. — ISBN 978-0321543257.
- Тейлор Д., Грин Н., Стаут У. Биология: В 3-х т. Т.1: Пер. с англ./Под. ред. Р. Сопера — 3-е изд., — М.: Мир, 2004. —454с., ил. ISBN 5-03-003685-7
- Tamarin R. H. Principles of Genetics (англ.). — 7th. — McGraw-Hill Education, 2001. — ISBN 0072334193.
- Robinson R et al. Genetics (Volume 1 A-D) (англ.). — MacMillan Reference USA, 2003. — ISBN 0-02-865607-5..
- Сингер М., Берг П. Гены и геномы: В 2-ох т. Т.2 Пер. с англ.— М.: Мир, 1998. — 391 с., ил. ISBN 5-03-002850-1
- Lykke-Andersen J., Aagaard C., Semionenkov M., Garrett R. A. Archaeal introns: splicing, intercellular mobility and evolution (англ.) // Trends in Biochemical Sciences. — 1997. — Vol. 22. — P. 326—331. — doi:10.1016/S0968-0004(97)01113-4. — PMID 9301331.
- Клетка // Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М. : Сов. энциклопедия, 1986. — С. 261—262. — 831 с. — 100 000 экз.
- Гилберт С. Биология развития: в 3-х томах. — М.: «Мир», 1995. — Т. 3. — 352 с. C. 202. — 5000 экз. — ISBN 5-03-001833-6
- Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки: В 3-х т.. — 2-ое, переработанное. — М.: «Мир», 1993. — Т. 2. — 539 с. С. 396. — ISBN 5-03-001987-1
- Митоз — статья из Большой советской энциклопедии. Алов И. А..
- Булдаков Л. А., Калистратова В. С. Радиоактивное излучение и здоровье. — М.: Информ-Атом, 2003. — 165 с. С. 39
- Benjamin Lewin. Chapter 13: The replicon // Genes VIII. — Upper Saddle River, NJ: Pearson Prentice Hall, 2004. — ISBN 0131439812.
- de Boer PA. Advances in understanding E. coli cell fission (англ.) // Current Opinion in Microbiology. — 2010. — Vol. 13. — P. 730—737. — doi:10.1016/j.mib.2010.09.015. — PMID 20943430.
- Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. Cells of the Adult Human Body — A Catalog Архивная копия от 4 марта 2016 на Wayback Machine (недоступная ссылка с 23-05-2013 [3203 дня]). — Garland Science, 2007
- Sell S. Cellular origin of cancer: dedifferentiation or stem cell maturation arrest? (англ.) // Environ. Health Perspect.. — 1993. — December (vol. 101, no. Suppl 5). — P. 15—26. — doi:10.2307/3431838. — PMID 7516873. — .
- Edinger A. L., Thompson C. B. Death by design: apoptosis, necrosis and autophagy (англ.) // Curr Opin Cell Biol.. — 2004. — Vol. 16, no. 6. — P. 663—669. — doi:10.1016/j.ceb.2004.09.011. — PMID 15530778.
- Altermann W., Kazmierczak J. Archean microfossils: a reappraisal of early life on Earth (англ.) // Research in Microbiology. — 2003. — Vol. 154, no. 9. — P. 611—617. — doi:10.1016/j.resmic.2003.08.006. — PMID 14596897.
- Oró J., Miller S. L., Lazcano A. The Origin and Early Evolution of Life on Earth (англ.) // Annual Review of Earth and Planetary Sciences. — Annual Reviews, 1990. — Vol. 18. — P. 317—356. — doi:10.1146/annurev.ea.18.050190.001533. — PMID 11538678.
- Chen I. et al. The Emergence of Cells During the Origin of Life (англ.) // Science. — 2006. — Vol. 5805, no. 314. — P. 1558—1559. — doi:10.1126/science.1137541. — PMID 11538678.
- Deamer D. W. The first living systems: a bioenergetic perspective (англ.) // Microbiology and Molecular Biology Reviews. — American Society for Microbiology, 1997. — Vol. 61, no. 2. — P. 239—261. — PMID 9184012.
- Cavalier-Smith T. Cell evolution and Earth history: stasis and revolution (англ.) // Philosophical Transactions of the Royal Society B: Biological Sciences. — 2006. — Vol. 1470, no. 361. — P. 623—630. — doi:10.1098/rstb.2006.1842. — PMID 16572163.
- Embley T. M., Martin W. Eukaryotic evolution, changes and challenges (англ.) // Nature. — 2006. — Vol. 7084, no. 440. — P. 969—1006. — doi:10.1038/nature04546. — PMID 16754610.
- Rivera M. C., Lake J. A. The ring of life provides evidence for a genome fusion origin of eukaryotes (англ.) // Nature. — 2004. — Vol. 7005, no. 431. — P. 152—155. — doi:10.1038/nature02848. — PMID 15356622.
- Gogarten J. P., Townsend JP. Horizontal gene transfer, genome innovation and evolution (англ.) // Nature Reviews Microbiology. — 2005. — Vol. 9, no. 3. — P. 679—687. — doi:10.1038/nrmicro1204. — PMID 16138096.
Литература
- Клетка // Биологический энциклопедический словарь / Гл. редактор Гиляров М. С.. — М.: «Советская энциклопедия», 1986. — С. 261—262. — 831 с. — 100 000 экз.
- Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки: В 3-х т. — 2-ое, переработанное. — М.: «Мир», 1993. — Т. 2. — 539 с. — ISBN 5-03-001987-1.
- Булдаков Л. А., Калистратова В. С. Радиоактивное излучение и здоровье. — М.: Информ-Атом, 2003. — 165 с. Архивная копия от 2 декабря 2003 на Wayback Machine
- Гилберт С. Биология развития: в 3-х томах. — М.: «Мир», 1995. — Т. 3. — 352 с. — 5000 экз. — ISBN 5-03-001833-6.
- Билич Г. Л., Крыжановский В. А. Биология. Полный курс: В 4 т. — издание 5-е, дополненное и переработанное. — М.: Издательство Оникс, 2009. — Т. 1. — 864 с. — ISBN 978-5-488-02311-6
- Вермель Е.М. История учения о клетке. — М.: «Наука», 1970. — 257 с. — 9600 экз.
- Иост Х. Физиология клетки. — М.: «Мир», 1975. — 864 с.
Периодические издания
- Цитология и генетика (ISSN 0564-3783) публикует статьи на русском, украинском и английском языках по выбору автора, переводится на английский язык (ISSN 0095-4527)
- BMC Cell Biology ISSN: 1471—2121 Журнал в открытом доступе (Импакт-фактор (2010) — 2,46)
- International Journal of Cell Biology Print ISSN: 1687-8876 Online ISSN: 1687-8884 Журнал в открытом доступе
- Cell (Импакт-фактор (2010) — 32.401)
- Nature Cell Biology ISSN: 1465-7392 eISSN: 1476-4679 (Импакт-фактор (2010) — 19.407)
- Journal of Molecular Cell Biology Print ISSN 1674—2788 Online ISSN 1759-4685 (Импакт-фактор (2010) — 13.400)
- Journal of cell biology Print ISSN: 0021-9525 Online ISSN: 1540-8140 (Импакт-фактор (2010) — 6.290)
Ссылки
- Наука о клетке // Лекция Е. Шеваля в проекте «ПостНаука» (15.04.2013)
- Alberts B., Johnson A. Lewis J. et al. Molecular Biology of the Cell. — 4th edition. — New York, 2002. — ISBN 0-8153-4072-9. — учебник по молекулярной биологии клетки на английском языке
- Библиотека изображений и видео внутреннего строения клетки Сell image library (англ.)
.svg.png.webp) Сквозь виртуальную клетку. Through the Virtual Cell (англ.)
Сквозь виртуальную клетку. Through the Virtual Cell (англ.)- Cell Structure
- Интерактивная анимация: строение прокариотической, животной и растительной клетки (англ.)
- Сборник материалов: AMAZING CELLS на сайте Learn Genetics (англ.)
- Интерактивная анимация: строение животной клетки (англ.)
| В библиографических каталогах |
|---|
| Эндомембранная система | |
|---|---|
| Цитоскелет | |
| Эндосимбионты | |
| Другие внутренние органеллы | |
| Внешние органеллы | |
