Нуклеопротеиды
Нуклеопротеиды — комплексы нуклеиновых кислот с белками.
К нуклеопротеидам относятся устойчивые комплексы нуклеиновых кислот с белками, длительное время существующие в клетке в составе органелл или структурных элементов клетки в отличие от разнообразных короткоживущих промежуточных комплексов «белок — нуклеиновая кислота» (комплексы нуклеиновых кислот с ферментами — синтетазами и гидролазами — при синтезе и деградации нуклеиновых кислот, комплексы нуклеиновых кислот с регуляторными белками и т.п.).
Структура и устойчивость
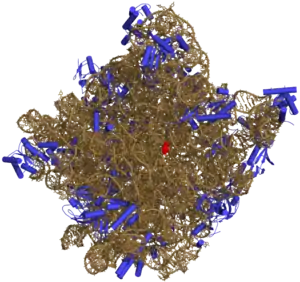
Жёлтым показана рРНК, синим — белки.
В зависимости от типа входящих в состав нуклеопротеидных комплексов нуклеиновых кислот различают рибонуклеопротеиды и дезоксирибонуклеопротеиды.
Устойчивость нуклеопротеидных комплексов обеспечивается нековалентным взаимодействием. У различных нуклеопротеидов в обеспечение стабильности комплекса вносят вклад различные типы взаимодействий, при этом нуклеиново-белковые взаимодействия могут быть специфичными и неспецифичными. В случае специфичного взаимодействия определённый участок белка связан со специфичной (комплементарной участку) нуклеотидной последовательностью, в этом случае вклад водородных связей, образующихся между нуклеотидными и аминокислотными остатками благодаря пространственному взаимному соответствию фрагментов, максимален. В случае неспецифичного взаимодействия основной вклад в стабильность комплекса вносит электростатическое взаимодействие отрицательно заряженных фосфатных групп полианиона нуклеиновой кислоты с положительно заряженными аминокислотными остатками белка.
Примером специфичного взаимодействия могут служить нуклеопротеидные комплексы рРНК — субъединицы рибосом; неспецифичное электростатическое взаимодействие характерно для хромосомных комплексов ДНК — хроматина и комплексов ДНК—протамины головок сперматозоидов некоторых животных.
Нуклеопротеиды диссоциируют на белки и нуклеиновые кислоты при воздействии агентов, разрушающих или ослабляющих нековалентные связи:
- повышенные концентрации солей или мочевины, увеличивающих ионную силу раствора,
- ионогенные поверхностно-активные вещества,
- некоторые полярные органические соединения (формамид и диметилформамид, фенол и т.п.).
Некоторые нуклеопротеиды (рибосомные субчастицы, нуклеокапсиды вирусов) обладают способностью к самосборке, то есть к образованию, при соответствующих условиях, нуклеопротеидов in vitro без участия клеточных структур или агентов; такая самосборка возможна в случае специфичных нуклеиново-белковых взаимодействий (нуклеиново-белкового узнавания). В любом случае, при образовании нуклеопротеидов происходят существенные конформационные изменения нуклеиновых кислот и, в некоторых случаях, белков, образующих нуклеопротеидный комплекс.
Распространённость и биологическая роль
Наиболее сильные конформационные изменения при образовании нуклеопротеидов претерпевают нуклеиновые кислоты, и эти изменения наиболее существенны в случае образования дезоксирибонуклеопротеидов. В отличие от одноцепочечной РНК, способной образовывать вторичные и третичные структуры за счёт антипараллельного комплементарного спаривания смежных отрезков цепи, двухцепочечная ДНК такой возможности не имеет и существует в растворах в виде значительно более «рыхлых», по сравнению с компактными глобулами РНК, клубков. Однако связывание ДНК с сильноосновными белками (гистонами и протаминами) за счёт электростатического взаимодействия приводит к значительно более плотно упакованным нуклеопротеидным комплексам — хроматинам, обеспечивающим компактное хранение ДНК и, соответственно, наследственной информации в составе хромосом эукариот. С другой стороны, большая конформационная подвижность РНК и её каталитические свойства приводят к большому разнообразию рибонуклеопротеидов, выполняющих различные функции.
Дезоксирибонуклеопротеиды
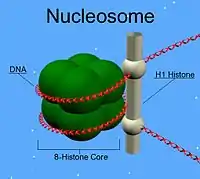
- Хроматин — комплекс ДНК с гистонами в соматических клетках эукариот. За счёт электростатического взаимодействия нить ДНК совершает двойной оборот вокруг октамера гистонного комплекса H2a, H2b, H3 и H4, образуя нуклеосомы, соединённые нитью ДНК. При присоединении к комплексу гистона H1 шесть нуклеосом образуют кольцеобразный комплекс, в результате происходит конденсация хроматина с образованием фибриллярной структуры, которая далее при присоединении топоизомеразы II и ряда вспомогательных белков способна конденсироваться в гетерохроматин. ДНК, связанная в таком нуклеопротеидном комплексе, не транскрибируется.
- Нуклеопротамины — комплекс ДНК с протаминами в сперматозоидах позвоночных, моллюсков и некоторых насекомых. За счет высокой основности протаминов и их меньшего, по сравнению с гистонами, размера, ДНК в нуклеопротаминах упакована значительно плотнее, чем в соматических клетках — в результате образования нейтрального комплекса ДНК с протаминами нуклеопротамины образуют плотные тороидальные структуры, содержащие ~50,000 пар оснований, соединенные короткими участками ДНК, связывающимися с ядерным скелетом. В свою очередь, тороиды при конденсации нуклеопротамина образуют фибриллы, в которых они упакованы «стопкой». Такая плотнейшая организация нуклеопротаминов обеспечивает защиту ДНК сперматозоидов от внешних факторов.
- Отдельным важным классом дезоксирибонуклеопротеидов являются вирусные нуклеопротеиды. Для репликации генетического материала ДНК-содержащих вирусов необходим перенос вирусной ДНК в ядро клетки, и такой транспорт и проникновение в ядро осуществляются в виде нуклеопротеидных комплексов, белки которых несут специфичные участки — сигналы ядерной локализации (nuclear localization signal, NLS), обеспечивающие транспорт через ядерные поры.
Рибонуклеопротеиды
В клетках в наибольших количествах содержатся два класса рибонуклеопротеидов:
- Нуклеопротеидные комплексы рибосомальных РНК (рРНП) — субъединицы рибосом — органелл, на которых происходит трансляция мРНК и синтез белков. Рибосомы представляют собой агрегаты из двух различных рРНП-субъединиц.
- Малые ядерные рибонуклеопротеиды (мяРНП) — нуклеопротеидные комплексы малых ядерных РНК, являющиеся субъединицами сплайсосом (участников сплайсинга ядерных транскриптов — предшественников зрелых РНК).
- Нуклеопротеидные комплексы мРНК — матричные рибонуклеопротеиды (мРНП), в русскоязычной литературе часто называемые информосомами (название предложено по аналогии с синонимическим названием мРНК — информационная РНК А. С. Спириным, идентифицировавшим такие комплексы в 1964 году в цитоплазме зародышей рыб). Биологическая роль мРНП весьма разнообразна: они, предположительно, участвуют в транспорте мРНК, стабилизации (защите от деградации при транспорте) и трансляционной регуляции. мРНП также являются химически наиболее разнообразным классом нуклеопротеидов, и их разнообразие определяется транскриптомом, то есть совокупностью мРНК, синтезируемых в клетке.
Нуклеокапсиды вирусов

1 — РНК, 2 — белковая субъединица, 3 — упакованный нуклеокапсид.
Нуклеокапсиды вирусов представляют собой достаточно плотно упакованные комплексы белков с нуклеиновой кислотой (ДНК или РНК у ретровирусов) и как функционально, так и структурно близки хроматину, представляя собой компактную форму вирусного генома.
Существует два основных типа нуклеокапсидных структур: палочковидная (нитевидная), либо сферическая («изометрическая»).
В первом случае связанные белковые субъединицы периодически располагаются вдоль нити нуклеиновой кислоты таким образом, что она сворачивается в спираль, образуя своего рода «инвертированную нуклеосому», в которой, в отличие от нуклеосом эукариот, белковая часть расположена не внутри, а снаружи структуры. Такая структура нуклеокапсидов типична для вирусов растений (в частности, вируса табачной мозаики) и миксо-, парамиксо- и рабдовирусов, нуклеокапсиды которых имеют спиральную форму.
В изометрических структурах упаковка нуклеиновой кислоты вирусного генома более сложна: белки оболочки нуклеокапсида относительно слабо связаны с нуклеиновой кислотой или нуклеопротеидами, что налагает минимум ограничений на способ упаковки нуклеиновой кислоты. Нуклеопротеиды «сердцевины» при этом могут быть весьма сложно организованы: так, в паповавирусах двухцепочечная кольцевая ДНК, связываясь с гистонами, образует структуры, очень похожие на нуклеосомы.
Литература
- А. С. Воронина. Трансляционная регуляция в раннем развитии. Успехи биологической химии, т. 42, 2002, с. 139—160
- А. С. Спирин. Рибонуклеиновые кислоты как центральное звено живой материи. Вестник РАН, том 73, № 2, с. 117—127 (2003)
- H. Hieronimus, P. Silver. System view of mRNP biology // Genes & Development
- Вирусология (под ред. Б. Филдса и Д. Найпа). М.: Мир, 1989.
| В библиографических каталогах |
|---|
| Эндомембранная система | |
|---|---|
| Цитоскелет | |
| Эндосимбионты | |
| Другие внутренние органеллы | |
| Внешние органеллы | |