Эндосома
Эндосома — мембранная внутриклеточная органелла, один из типов везикул, образующаяся при слиянии и созревании эндоцитозных пузырьков. Зрелые эндосомы представляют собой образования размером 300-400 нм.
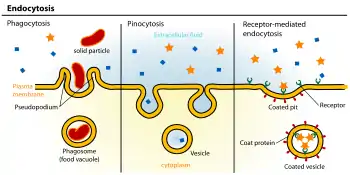
Функция
Большинство эндосом, образующихся в результате эндоцитоза из плазматической мембраны, транспортируются внутрь клетки, где сливаются с существующими эндосомами либо закисляются за счёт активности протонной АТФазы (H-АТФаза). В процессе созревания эндосома проходит несколько последовательных стадий, постепенно превращаясь в лизосому. При этом часть изначального материала плазматической мембраны может вернуться обратно для повторного использования (рециркуляция).
Роль в передаче сигналов в клетке
Многие мембранные рецепторы после связывания субстрата интернализуются в составе эндосом. Ранее это явление рассматривали как путь деградации или рециркуляции молекул рецепторов. Однако сегодня ясно, что локализация в эндосомах может играть особую роль в способности рецепторов к передаче сигнала. Так, например, известно, что рецептор эпидермального фактора роста способен к передаче сигнала сразу после связывания с субстратом на клеточной мембране, однако максимальной активности он достигает только в эндосомах. Кроме того, активированные рецепторы на мембране и в эндосомах могут запускать разные сигнальные пути. Например, толл-подобный рецептор 4, локализованный в клеточной мембране, передаёт сигнал по пути TIRAP-MyD88, а локализованный в эндосомах — по пути TRAM-TRIF[1].
Типы
Различают три типа эндосом: ранние, или первичные, эндосомы, поздние эндосомы (или мультивезикулярные тельца) и рециркулирующие эндосомы. [2] Они различаются по времени, требуемом для достижения эндоцитированного материала этих эндосом, и по функциональным маркёрам, таким как различные формы белков из группы Rab.[3] Кроме этого, они различаются по своей морфологии. После того, как эндоцитарные везикулы теряют оболочку, они сливаются с ранними эндосомами, которые в свою очередь в процессе созревания превращаются в поздние эндосомы перед тем, как слиться с лизосомами.[4][5]
Преобразование ранних эндосом, или процесс созревания, может происходить несколькими путями. Важную роль в этом процессе играют вакуолярные H-ATФазы, которые переносят протоны внутрь эндосомы и закисляют её.[6] Рециркулирующие молекулы (как правило рецепторы) собираются в трубчатых структурах эндосом. После рециркуляции и потери трубчатых структур образующиеся поздние эндосомы содержат только вакуолеобразные структуры. Они постепенно увеличиваются в размере по мере гомотипического слияния.[7] Молекулы также сортируются в мелкие везикулы, которые выпячиваются по периметру мембраны внутрь эндосомы, формируя люминальные везикулы. Это приводит к образованию мульти везикулярной организации поздних эндосом (мультивезикулярных телец). Одоновременно продолжается рециркуляция некоторых молекул, включая трансфериновый рецептор и маннозо-6-фосфатный рецептор. [4] Эндосомы теряют RAB5 и получают RAB7 и становятся компетентмыми для последующего слияния с лизосомами.[7]
Слияние поздних эндосом с лизосомами приводит первоначально к образованию гибридной структуры с промежуточными характеристиками.[8] Так, например, лизосомы обладают большей плотностью, чем эндосомы, в то время как такие гибридные структуры имеют промежуточную плотность.
Примечания
- Chaturvedi A., Martz R., Dorward D., Waisberg M., Pierce S. K. Endocytosed BCRs sequentially regulate MAPK and Akt signaling pathways from intracellular compartments // Nat Immunol. — 2011. — Т. 12, вып. 11. — С. 1119—1126. — doi:10.1038/ni.2116. — PMID 21964606.
- Mellman I. Endocytosis and molecular sorting (англ.) // Annual Review of Cell and Developmental Biology : journal. — 1996. — Vol. 12. — P. 575—625. — doi:10.1146/annurev.cellbio.12.1.575. — PMID 8970738.
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic (англ.) // Nat Rev Mol Cell Biol : journal. — 2009. — August (vol. 10, no. 8). — P. 513—525. — doi:10.1038/nrm2728. — PMID 19603039.
- Futter, CE.; Pearse, A.; Hewlett, LJ.; Hopkins, CR. Multivesicular endosomes containing internalized EGF-EGF receptor complexes mature and then fuse directly with lysosomes (англ.) // J Cell Biol : journal. — 1996. — March (vol. 132, no. 6). — P. 1011—1023. — doi:10.1083/jcb.132.6.1011. — PMID 8601581.
- Luzio J.P., Rous B.A., Bright N.A., Pryor P.R., Mullock B.M., Piper RC. Lysosome-endosome fusion and lysosome biogenesis (англ.) // Journal of Cell Science. — The Company of Biologists, 2000. — Vol. 113. — P. 1515—1524. — PMID 10751143.
- Lafourcade, C.; Sobo, K.; Kieffer-Jaquinod, S.; Garin, J.; van der Goot, FG. Regulation of the V-ATPase along the endocytic pathway occurs through reversible subunit association and membrane localization (англ.) // PLoS One : journal / Joly, Etienne. — 2008. — Vol. 3, no. 7. — P. e2758. — doi:10.1371/journal.pone.0002758. — PMID 18648502.
- Rink, J.; Ghigo, E.; Kalaidzidis, Y.; Zerial, M. Rab conversion as a mechanism of progression from early to late endosomes (англ.) // Cell : journal. — Cell Press, 2005. — September (vol. 122, no. 5). — P. 735—749. — doi:10.1016/j.cell.2005.06.043. — PMID 16143105.
- Mullock, BM.; Bright, NA.; Fearon, CW.; Gray, SR.; Luzio, JP. Fusion of lysosomes with late endosomes produces a hybrid organelle of intermediate density and is NSF dependent (англ.) // J Cell Biol : journal. — 1998. — February (vol. 140, no. 3). — P. 591—601. — doi:10.1083/jcb.140.3.591. — PMID 9456319.
Ссылки
- Alberts, Bruce; et al. Essential Cell Biology (англ.). — 2nd Edition. — New York, NY: Garland Science, 2004.(Cell and Molecular Biology)
| Эндомембранная система | |
|---|---|
| Цитоскелет | |
| Эндосимбионты | |
| Другие внутренние органеллы | |
| Внешние органеллы | |