Грамотрицательные бактерии
Грамотрица́тельные бакте́рии (обозначаются Грам (-)) — бактерии, которые не окрашиваются кристаллическим фиолетовым при использовании окраски микроорганизмов по методу окрашивания по Граму[1]. В отличие от грамположительных бактерий, которые сохраняют фиолетовую окраску даже после промывания обесцвечивающим растворителем (спирт), грамотрицательные полностью обесцвечиваются. После промывания растворителем при окрашивании по Граму добавляется контрастный краситель (обычно сафранин), который окрашивает все грамотрицательные бактерии в красный или розовый цвет. Это происходит из-за наличия внешней мембраны, препятствующей проникновению красителя внутрь клетки. Сам по себе тест полезен при классификации бактерий и разделении их на две группы относительно строения их клеточной стенки. Из-за своей более мощной и непроницаемой клеточной стенки грамотрицательные бактерии более устойчивы к антителам, чем грамположительные.

Обычно патогенность грамотрицательных бактерий связывают с определёнными компонентами их клеточных стенок, а именно, с липополисахаридным слоем (ЛПС или эндотоксический слой)[1]. В человеческом организме ЛПС вызывает иммунный ответ, который характеризуется синтезом цитокинов и активацией иммунной системы. Обычной реакцией на синтез цитокинов является воспаление, что также может привести к увеличению количества токсичных веществ в организме хозяина.
Характеристика
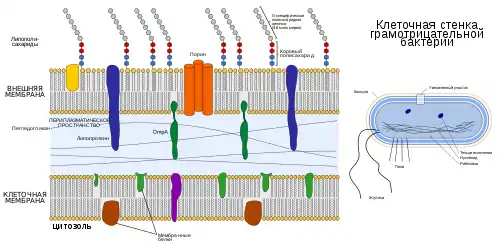
Общие признаки, свойственные большинству грамотрицательных бактерий:
- Наличие двух мембран, между которыми находится клеточная стенка и периплазматическое пространство.
- Более тонкий, по сравнению с грамположительными бактериями, пептидогликановый слой.
- Наружная мембрана содержит липополисахариды (состоит из липида А, полисахаридного ядра и антигена О снаружи и из фосфолипидов изнутри).
- В наружной мембране присутствуют порины, функционирующие подобно порам для определённых молекул.
- S-слой прикреплен к наружной мембране, а не к пептидогликановому слою.
- Если есть жгутик, он имеет четыре поддерживающих кольца, а не два.
- Отсутствуют тейхоевая и липотейхоевая кислоты.
- Обычно не образуют спор (примечательным исключением является Coxiella burnetii, образующая спороподобные структуры).
- Липопротеины прикреплены непосредственно к полисахаридной основе.
- Большинство содержат липопротеин Брауна, который связывает наружную мембрану и цепочки пептидоглюканов ковалентной связью.
Химический состав и структура клеточных покровов
Далее рассмотрены слои клеточных покровов грамотрицательных прокариотов, начиная с самого внутреннего:
Клеточная стенка
У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных. В её состав входит гораздо большее число макромолекул разного химического типа. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями.
Некоторые скользящие бактерии (миксобактерии, флексибактерии) способны в процессе перемещения по твердому субстрату периодически менять форму клеток, например путём изгибания, что говорит об эластичности их клеточной стенки, и в первую очередь её пептидогликанового слоя. Электронно-микроскопическое изучение, однако, обнаружило у них клеточную стенку, типичную для грамотрицательных эубактерий. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий — чрезвычайно низкая сшитость её пептидогликанового компонента[2].
Периплазматическое пространство
Появление у грамотрицательных эубактерий дополнительной мембраны в составе клеточной стенки фактически привело к созданию обособленной полости (периплазматического пространства), отграниченной от цитоплазмы и внешней среды специфическими мембранами и несущей важную функциональную нагрузку[2].
Периплазматическое пространство, куда погружен пептидогликановый слой, заполнено раствором, в состав которого входят специфические белки, олигосахариды и неорганические молекулы. Периплазматические белки представлены двумя типами: транспортными белками и гидролитическими ферментами.
Было обнаружено также, что многие бактерии способны в больших количествах вырабатывать ферменты (гликозидазы, протеазы, липазы и проч.), гидролизующие все типы полимерных молекул. Последними могут быть как молекулы, синтезируемые самой клеткой, так и чужеродные, попавшие в клетку извне. Отрицательные последствия гидролиза собственных молекул (самопереваривание) очевидны. В то же время прокариоты нуждаются в гидролитических ферментах, так как это расширяет круг используемых ими веществ, включая в него полимеры разного типа. Становится понятна необходимость изолирования этих ферментов от цитоплазматического содержимого. Грамположительные эубактерии выделяют гидролитические ферменты во внешнюю среду, у грамотрицательных они локализованы в периплазматическом пространстве.
Внешняя мембрана
Снаружи от пептидогликана располагается дополнительный слой клеточной стенки — наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида. Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30—40 % её поверхности и локализованный во внешнем слое.
Белки наружной мембраны можно разделить на основные и минорные. Основные белки представлены небольшим числом различных видов, но составляют почти 80 % всех белков наружной мембраны. Одна из функций этих белков — формирование в мембране гидрофильных пор диаметром примерно 1—15 нм и длиной 50—70 нм, наклонённых к поверхности клеточной стенки по углом 30-40°. Через них осуществляется неспецифическая диффузия молекул с массой до 600—900 Да. Это означает, что через такие поры могут проходить сахара, аминокислоты, небольшие олигосахариды и пептиды. Белки, пронизывающие наружную мембрану насквозь и образующие гидрофильные поры, называют поринами. Минорные белки наружной мембраны представлены гораздо бо́льшим числом видов. Их основная функция — транспортная и рецепторная. Примером минорных белков могут служить белки, ответственные за специфический транспорт в клетку железосодержащих соединений.
Разнообразные функции выполняют макромолекулы, локализованные частично или полностью на внешней стороне клеточной стенки, контактирующей с окружающей средой; это специфические рецепторы для фагов и колицинов; антигены; макромолекулы, обеспечивающие межклеточные взаимодействия при конъюгации, а также между патогенными бактериями и тканями высших организмов.
S-слой и осцилиновые фибриллы
S-слой или зубчатый слой — слой из плотноупакованных белков, выстилающих наружную поверхность клетки и обеспечивающих защиту от резких изменений pH или концентраций каких-либо ионов. Поверх этого слоя расположен слой из волоскоподобных фибрилл, находится за пределами наружной мембраны цианобактерий, способных к скольжению. Похожие на волоски, фибриллы верхнего слоя состоят из стержнеподобного гликопротеина, называемого осциллин.
Скольжение происходит посредством секреции слизи через поры на внешнюю сторону клеточных покровов[3]. Слизь проходит вдоль поверхности из осциллиновых фибрилл наружного слоя клетки и по расположенному рядом субстрату, продвигая фибриллы вперёд. Совокупность организованных волоскоподобных фибрилл, таким образом, действует как пассивный винт, в то время как слизь проходит по их поверхности в процессе скольжения[4].
Классификация
Наряду с анализом формы клетки, окрашивание по Граму является быстрым диагностическим методом, который ранее использовался для группирования видов бактерий в подотделы.

Основываясь на окраске по Граму, ныне ликвидированное царство Monera было разделено на четыре дивизиона: Firmacutes (+), Gracillicutes (-), Mollicutes (0) и Mendocutes (var.)[5]. Но начиная уже с 1987 года монофилия грамотрицательных бактерий была поставлена под сомнение, а позже и полностью опровергнута бимолекулярными исследованиями[6]. Однако некоторые авторы, такие как Т. Кавалир-Смит, всё ещё считают их монофилитической кладой и относят к подцарству Negibacteria[7].
Строение внешней клеточной мембраны и классификация бактерий
Важно указать на то, что хотя бактерии традиционно делятся на две основные группы, грамположительные и грамотрицательные, такая классификация является двусмысленной и условной, поскольку может относиться к трём совершенно разным аспектам (результат окрашивания, организация клеточных покровов, таксономическая группа), которые необязательно совпадают для всех видов бактерий[8][9][10][11]. Реакция на окрашивание грамположительных и грамотрицательных бактерий не является надёжной характеристикой также и потому, что эти два вида бактерий не формирует филогенетически единую группу[8]. Как бы то ни было, хотя окрашивание по Граму и является эмпирическим критерием, в его основе лежат отчётливые различия в ультраструктуре и химическом составе двух главных видов прокариотических клеточных стенок, встречающихся в природе. Оба этих вида клеточных стенок отличаются друг от друга по наличию или отсутствию внешней липидной мембраны, которая является более надёжной и фундаментальной характеристикой бактериальных клеток[8][12]. Все грамположительные бактерии окружены единственным слоем фосфолипидной мембраны и обычно имеют толстый слой (20—80 нм) из пептидогликанов (муреин и проч.), сохраняющий на себе краситель Грама. Ряд других бактерий, окружённых единственной мембраной, но окрашивающихся грамотрицательно из-за отсутствия пептидогликанового слоя (см. микоплазмы) или же своей неспособности удерживать краситель Грама вследствие специфического состава клеточной стенки, также находится в близком родстве с грамположительными бактериями. Для бактериальных клеток, окружённых одной клеточной мембраной, был предложен термин монодермные бактерии или монодермные прокариоты[8][8][12]. В противоположность грамположительным бактериям, все архетипичные грамотрицательные бактерии, помимо цитоплазматической мембраны, окружены ещё и внешней клеточной мембраной и содержат между ними крайне тонкий слой пептидогликанов (2—3 нм). Наличие внутренней и наружной мембран порождает новый клеточный компартмент — периплазматическое пространство. Такие бактерии/прокариоты были обозначены как дидермные бактерии[8][8][12].
Ещё одно важное различие между монодермными и дидермными прокариотами — консервативный набор делеций в ряде важных белков (см. DnaK, GroEL)[8][9][12][13]. Из двух структурно различных групп прокариотических организмов, монодермные бактерии считаются предковой группой по отношению к дидермным. Основываясь на ряде наблюдений, включая тот факт, что грамположительные бактерии в большинстве своём продуценты антибиотиков, а грамотрицательные бактерии обычно к таковым устойчивы, было высказано предположение, что у наружной клеточной мембраны грамотрицательных бактерий существует защитный механизм против селективного действия антибиотиков[8][9][12][13]. Некоторые бактерии, как например Deinococcus, окрашиваются грамположительно из-за толстого пептидогликанового слоя, но обладают наружной клеточной мембраной и считаются промежуточным звеном между монодермами (грамположительные) и дидермами (грамотрицательные)[8][13]. Дидермные бактерии в свою очередь могут быть подразделены на простых дидерм, у которых нет слоя липолисахаридов, архетипичных дидерм, чья наружная клеточная мембрана содержит липолисахариды, и собственно дидерм, наружная мембрана которых состоит из миколовой кислоты[10][11][13][14].
Вдобавок многие бактериальные таксоны (см. Negativicutes, Fusobacteria, Synergistetes и Elusimicrobia), которые являются частью типа Firmicutes или его близкородственной ветвью, также обладают дидермной структурой клетки[11][13][14]. Однако консервативная делеция (CSI) в белке HSP60 (GroEL) служит отличительным знаком всех традиционных типов грамотрицательных бактерий (то есть Proteobacteria, Aquificae, Chlamydiae, Bacteroidetes, Chlorobi, Cyanobacteria, Fibrobacteres, Verrucomicrobia, Planctomycetes, Spirochetes, Acidobacteria и т. д.) от атипичных дидерм и других типов монодермных бактерий (таких как Actinobacteria, Firmicutes, Thermotogae, Chloroflexi и т. д.)[13]. Наличие CSI во всех секвенированых видах из обычных ЛПС-содержащих типов грамотрицательных бактерий доказывает, что этот тип бактерий формирует монофилитическую кладу, и говорит в пользу того, что в этой группе ни у одного вида не происходила потеря внешней мембраны. Эти данные говорят против гипотезы о происхождении монодермных прокариот от дидермных путём потери внешней мембраны[13].
Примеры видов
Протеобактерии — большая группа грамотрицательных бактерий, включающая Escherichia coli (E. coli), Salmonella, Shigella и прочих Enterobacteriaceae, Pseudomonas, Moraxella, Helicobacter, Stenotrophomonas, Bdellovibrio, уксуснокислых бактерий, Legionella и множество других. Другие примечательные группы грамотрицательных бактерий включают в себя цианобактерий, спирохет, зелёных серо- и не-серобактерий.
Связанные с медициной грамотрицательные кокки включают в себя три организма, вызывающих заболевания, передающиеся половым путём (Neisseria gonorrhoeae), менингит (Neisseria meningitidis) и респираторные симптомы (Moraxella catarrhalis).
Среди бацилл есть большое количество важных для медицины видов. Некоторые из них в первую очередь вызывают болезни органов дыхания, как например (Hemophilus influenzae, Klebsiella pneumoniae, Legionella pneumophila, Pseudomonas aeruginosa), выделения (Escherichia coli , Proteus mirabilis, Enterobacter cloacae, Serratia marcescens) и пищеварения (Helicobacter pylori, Salmonella enteritidis, Salmonella typhi).
Ряд грамотрицательных бактерий связан с внутрибольничными инфекциями. В их числе Acinetobacter baumannii, вызывающий бактериемии, вторичный менингит и вентиляторную пневмонию у больных в отделениях реанимации.
Значение для медицины
Одна из нескольких уникальных характеристик грамотрицательных бактерий — это структура внешней клеточной мембраны. Внешний лист мембраны включает в себя комплекс липополисахаридов, липидные части которых выполняют роль эндотоксинов. Если эндотоксины попадают в транспортную систему, происходит интоксикация, с последующим увеличением температуры, повышением частоты дыхания и понижением кровяного давления. Всё это может привести к токсическому шоку и последующему летальному исходу.
Эта внешняя мембрана защищает бактерию от некоторых антибиотиков, красителей и детергентов, которые в норме повреждают внутреннюю мембрану или клеточную стенку. Такая защита делает бактерию устойчивой к лизоциму и пенициллину. Однако были разработаны альтернативные методы лечения, такие как лизоцим с ЭДТА и антибиотик ампициллин, способные справиться с защитной внешней мембраной некоторых патогенных грамотрицательных организмов. Для этой же цели могут быть использованы и другие лекарства, наиболее эффективные из которых хлорамфеникол, стрептомицин и налидиксовая кислота.
См. также
Источники
- 'Salton M. R. J., Kim K. S.. Structure. in: Baron's Medical Microbiology (Baron S. et al., eds.). — 4th. — Univ of Texas Medical Branch, 1996. — ISBN 0-9631172-1-1.
- Гусев М. В., Минеева Л. А. Микробиология: Учебник. — 2003. − 464 с.
- (Hoiczyk и Baumeister, 1998; Hoiczyk, 2000)
- Lee R. E. Phycology. — 4th Ed. — Colorado State University, USA.
- Gibbons, N. E.; Murray, R. G. E. Proposals Concerning the Higher Taxa of Bacteria (англ.) // International Journal of Systematic Bacteriology : journal. — 1978. — Vol. 28, no. 1. — P. 1—6. — doi:10.1099/00207713-28-1-1.
- Woese C. R. Bacterial evolution. (англ.) // Microbiological reviews. — 1987. — Vol. 51, no. 2. — P. 221—271. — PMID 2439888.
- Cavalier-Smith T. Rooting the tree of life by transition analyses (англ.) // Biol. Direct : journal. — 2006. — Vol. 1. — P. 19. — doi:10.1186/1745-6150-1-19. — PMID 16834776.
- Gupta R. S. (1998) Protein phylogenies and signature sequences: A reappraisal of evolutionary relationships among archaebacteria, eubacteria and eukaryotes. Microbiol. Mol. Biol. Rev. 62: 1435—1491.
- Gupta R. S. (2000) The natural evolutionary relationships among prokaryotes. Crit. Rev. Microbiol. 26: 111—131.
- Desvaux Mickaël, Hébraud Michel, Talon Régine, Henderson Ian R. Secretion and subcellular localizations of bacterial proteins: a semantic awareness issue // Trends in Microbiology. — 2009. — Апрель (т. 17, № 4). — С. 139—145. — ISSN 0966-842X. — doi:10.1016/j.tim.2009.01.004.
- Sutcliffe Iain C. A phylum level perspective on bacterial cell envelope architecture // Trends in Microbiology. — 2010. — Октябрь (т. 18, № 10). — С. 464—470. — ISSN 0966-842X. — doi:10.1016/j.tim.2010.06.005.
- Gupta R. S. (1998). What are archaebacteria: life’s third domain or monoderm prokaryotes related to Gram-positive bacteria? A new proposal for the classification of prokaryotic organisms. Molecular Microbiology. 29(3):695-707.
- Gupta R. S. (2011). Origin of diderm (Gram-negative) bacteria: antibiotic selection pressure rather than endosymbiosis likely led to the evolution of bacterial cells with two membranes. Antonie van Leeuwenhoek. 100:171—182.
- Marchandin H., Teyssier C., Campos J., Jean-Pierre H., Roger F., Gay B., Carlier J. P., Jumas-Bilak E. Negativicoccus succinicivorans gen. nov., sp. nov., isolated from human clinical samples, emended description of the family Veillonellaceae and description of Negativicutes classis nov., Selenomonadales ord. nov. and Acidaminococcaceae fam. nov. in the bacterial phylum Firmicutes. (англ.) // International journal of systematic and evolutionary microbiology. — 2010. — Vol. 60, no. Pt 6. — P. 1271—1279. — doi:10.1099/ijs.0.013102-0. — PMID 19667386.
| В библиографических каталогах |
|---|