Деление прокариотических клеток
Деление прокариотических клеток — процесс образования дочерних прокариотических клеток из материнской. Ключевыми событиями клеточного цикла как прокариот, так и эукариот являются репликация ДНК и деление клетки. Отличительной чертой деления прокариотических клеток является непосредственное участие реплицированной ДНК в процессе деления[1]. В подавляющем большинстве случаев прокариотические клетки делятся с образованием двух одинаковых по размеру дочерних клеток, поэтому этот процесс ещё иногда называют бинарным делением. Так как чаще всего прокариотические клетки имеют клеточную стенку, бинарное деление сопровождается образованием септы — перегородки между дочерними клетками, которая затем расслаивается посередине. Процесс деления прокариотической клетки подробно изучен на примере Escherichia coli[2].

Деление грамотрицательных бактерий
Раскрытию механизма деления грамотрицательных бактерий способствовало изучение мутантных штаммов E. coli, у которых этот механизм нарушен. В результате мутаций, которые затрагивают гены, участвующие в делении клетки, могут формироваться следующие фенотипы:
- филаменты — длинные клетки, которые формируются, если септа по тем или иным причинам не образуется. Филаменты бывают нескольких типов:
- содержащие многочисленные нуклеоиды, равномерно распределённые по длине клетки. В таких штаммах сегрегация ДНК проходит нормально, но септа тем не менее не формируется; их называют Fts− (от англ. filamentation temperature-sensitive)[3];
- содержащие единственный нуклеоид примерно посередине клетки. В данном случае причиной образования филаментов являются нарушения в синтезе ДНК, соответственно штаммы называют Dna−[4];
- содержащие многочисленные нуклеоиды посередине клетки. В дальнейшем ближе к концам таких клеток могут формироваться септы, и вследствие этого образовываться безъядерные клетки (см. ниже). Эти события являются результатом нарушений в механизме сегрегации ДНК, соответствующие штаммы чаще всего называются Par− (от англ. partition)[4];
- миниклетки — маленькие, лишённые ДНК клетки. Миниклетки образуются, когда при делении формируется больше одной септы или она находится в неправильном месте. Штаммы с такими нарушениями называют Min− (от англ. miniature)[5];
- безъядерные клетки — клетки нормального размера, лишённые ДНК. Как было сказано выше, безъядерные клетки могут образовываться из филаментов типа Par−. В то же время при некоторых мутациях, например Muk−(от яп. mukaku — безъядерный), в популяции клеток могут обнаруживаться только безъядерные клетки при отсутствии филаментов. Тем не менее такой фенотип также связан с нарушением сегрегации ДНК[6].
Молекулярный механизм деления
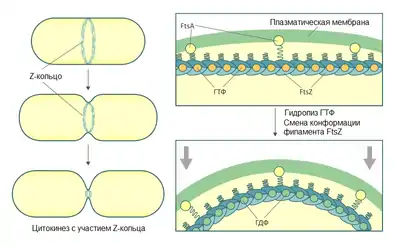
Центральную роль в делении клеток грамотрицательных бактерий играет септальное кольцо — кольцевая органелла, расположенная примерно посередине клетки и способная сокращаться, образуя перетяжку между двумя новыми дочерними клетками. Зрелое септальное кольцо представляет собой сложный белковый комплекс, состоящий более чем из дюжины разных белков. Десять из них (FtsA, B, I, K, L, N, Q, W, Z и ZipA) абсолютно необходимы для формирования септы, и нарушение в их работе приводит к формированию филаментов типа Fts−[2]. Остальные компоненты не являются строго необходимыми, их функции могут частично перекрываться. Формирование септального кольца происходит в несколько этапов, новые белки присоединяются по одному в таком порядке: FtsZ→FtsA/ZipA→FtsK→FtsQ→FtsL/FtsB→FtsW→FtsI→FtsN[7].
Белки, входящие в состав септального кольца, помимо FtsZ, можно разделить на несколько классов по функциям:
- модулирующие сборку филаментов FtsZ (FtsA, ZipA, ZapA, ZapB);
- связывающие Z-кольцо с мембраной (FtsA, ZipA);
- координирующие образование септы с сегрегацией ДНК (FtsK);
- синтезирующие (модулирующие) пептидогликан (FtsI, FtsW);
- гидролизующие пептидогликан для расхождения дочерних клеток (AmiC, EnvC).
Однако для многих белков септального кольца точная функция до сих пор не известна[8].
Формирование Z-кольца
Незрелую форму септального кольца называют Z-кольцом, по имени белка FtsZ, который играет ключевую роль в его формировании. Однако стоит отметить, что часто термины септальное кольцо и Z-кольцо используют как синонимы, поэтому в каждом отдельном случае это нужно оговаривать особо[2]. Белок FtsZ имеет тенденцию формировать длинные фибриллярные структуры. После деления FtsZ формирует прилегающую ко внутренней мембране спираль, закрученную вдоль оси клетки. Эта спираль постоянно меняет своё положение и быстро осциллирует от одного полюса клетки к другому[9][10]. Примерно ко времени завершения репликации ДНК спираль FtsZ схлопывается, в результате чего формируется Z-кольцо посередине клетки[11]. Есть все основания предполагать, что Z-кольцо на самом деле также представляет собой короткую плотную спираль[10].
Белок FtsZ — прокариотический гомолог тубулина с похожей третичной структурой[1]. Это позволяет предполагать, что ассоциация FtsZ в Z-кольцо может напоминать сборку микротрубочек эукариот. FtsZ, как и тубулин, обладает ГТФазной активностью, гидролиз ГТФ обеспечивает полимеризацию FtsZ с образованием линейных протофиламентов. Z-кольцо — динамичная структура: молекулы FtsZ в составе кольца постоянно заменяются молекулами из цитоплазматического пула[12][13].
FtsZ сам по себе не имеет сродства к мембране, формирование кольцевой структуры из протофиламентов, их закрепление во внутренней мембране и стабилизацию Z-кольца обеспечивают белки FtsA и ZipA, которые взаимодействуют с FtsZ прямо и независимо. ZipA — интегральный белок внутренней мембраны, FtsA — цитоплазматический белок, который тем не менее может связываться с мембраной за счёт особой аминокислотной последовательности на C-конце. ZipA, по-видимому, характерен только для γ-протеобактерий, в то время как FtsA более универсален[2]. Z-кольцо у E. coli может формироваться при отсутствии одного из этих белков, но не двух сразу, что указывает на их перекрывающиеся функции[14][15].
Ещё два белка — ZapA и ZapB — включаются в состав Z-кольца на ранней стадии, однако их присутствие не строго обязательно для его формирования[2][7][16]. ZapA — универсальный для многих прокариот белок, а вот ZapB, по всей вероятности, есть только у γ-протеобактерий. ZapA связывается с FtsZ непосредственно, а ZapB связывается с ZapA. Интересно, что ZapB формирует ещё одну кольцевую структуру, которая находится дальше от мембраны, чем Z-кольцо. Функции этих белков ещё до конца не установлены, однако предполагается, что они принимают участие в превращении спирали FtsZ в Z-кольцо, а также в последующей стабилизации Z-кольца[7].
Созревание септального кольца
Z-кольцо существует в описанном виде 14—21 минуту (в зависимости от скорости деления), и только после этого к нему присоединяются все остальные ключевые белки, начиная с FtsQ[17]. В какое время присоединяется FtsK, пока точно не установлено. Оставшиеся белки включаются в состав септального кольца практически одновременно в течение 1—3 минут. До того момента, как начинает собираться септальное кольцо, Z-кольцо стимулирует синтез пептидогликана в центре клетки таким образом, что клетка начинает удлиняться. Молекулярный механизм этого процесса, однако, до сих пор не установлен[2][17].
Одними из последних в септальное кольцо включаются белки, ответственные за синтез полярного пептидогликана (FtsW, FtsI), и белки, обеспечивающие частичный гидролиз пептидогликана на границе раздела между двумя клетками (AmiA, B, C, EnvC, NlpD)[2].
Формирование перетяжки
Завершающим этапом деления прокариотической клетки является формирование перетяжки и конечное разделение двух новых клеток. Образование перетяжки затрагивает все компоненты клеточной оболочки (внутреннюю мембрану, слой пептидогликана и внешнюю мембрану). Есть основания полагать, что за инвагинацию внутренней мембраны отвечает Z-кольцо, однако как именно оно передаёт напряжение на мембрану, пока не известно. Параллельно с этим процессом ферменты септального кольца синтезируют (или модифицируют особым образом предсуществующий) пептидогликан септы[2][17]. После формирования септы в работу вступают пептидогликангидролазы, которые отделяют будущие клетки друг от друга. Завершается процесс деления инвагинацией и обособлением внешних мембран клеток.
Примечания
- Benjamin Lewin. Chapter 13: The replicon // Genes VIII. — Upper Saddle River, NJ: Pearson Prentice Hall, 2004. — ISBN 0131439812.
- de Boer PA. Advances in understanding E. coli cell fission (неопр.) // Curr Opin Microbiol.. — 2010. — Т. 13. — С. 730—737. — doi:10.1016/j.mib.2010.09.015. — PMID 20943430.
- Adler H. I., Hardigree AA. Growth and Division of Filamentous Forms of Escherichia coli (англ.) // Journal of Bacteriology : journal. — 1965. — Vol. 90. — P. 223—226. — PMID 16562021.
- Hirota Y., Ryter A., Jacob F. Thermosensitive mutants of E. coli affected in the processes of DNA synthesis and cellular division (англ.) // Cold Spring Harb Symp Quant Biol. : journal. — 1968. — Vol. 33. — P. 677—693. — PMID 4892005.
- Adler H. I., Fisher W. D., Cohen A., Hardigree AA. MINIATURE escherichia coli CELLS DEFICIENT IN DNA (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1967. — Vol. 57. — P. 321—326. — PMID 16591472.
- Hiraga S., Niki H., Ogura T., Ichinose C., Mori H., Ezaki B., Jaffé A. Chromosome partitioning in Escherichia coli: novel mutants producing anucleate cells (англ.) // Journal of Bacteriology : journal. — 1989. — Vol. 171. — P. 1496—1505. — PMID 2646284.
- Galli E., Gerdes K. Spatial resolution of two bacterial cell division proteins: ZapA recruits ZapB to the inner face of the Z-ring. (англ.) // Microbiology : journal. — Microbiology Society, 2010. — Vol. 76. — P. 1514—1526. — doi:10.1111/j.1365-2958.2010.07183.x. — PMID 20487275.
- Weiss DS. Bacterial cell division and the septal ring. (англ.) // Microbiology : journal. — Microbiology Society, 2004. — Vol. 54. — P. 588—597. — doi:10.1111/j.1365-2958.2004.04283.x. — PMID 15491352.
- Thanedar S., Margolin W. FtsZ exhibits rapid movement and oscillation waves in helix-like patterns in Escherichia coli. (англ.) // Curr Biol. : journal. — 2004. — Vol. 14. — P. 1167—1173. — doi:10.1016/j.cub.2004.06.048. — PMID 15242613.
- Erickson H. P., Anderson D. E., Osawa M. FtsZ in bacterial cytokinesis: cytoskeleton and force generator all in one. (англ.) // Microbiology and Molecular Biology Reviews : journal. — American Society for Microbiology, 2010. — Vol. 74. — P. 504—528. — doi:10.1128/MMBR.00021-10. — PMID 21119015.
- Den Blaauwen T., Buddelmeijer N., Aarsman M. E., Hameete C. M., Nanninga N. Timing of FtsZ assembly in Escherichia coli. (англ.) // Curr Biol. : journal. — 1999. — Vol. 181. — P. 5167—5175. — PMID 10464184.
- Stricker J., Maddox P., Salmon E. D., Erickson HP. Rapid assembly dynamics of the Escherichia coli FtsZ-ring demonstrated by fluorescence recovery after photobleaching. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2002. — Vol. 99. — P. 3171—3175. — doi:10.1073/pnas.052595099. — PMID 11854462.
- Romberg L., Levin PA. Assembly dynamics of the bacterial cell division protein FTSZ: poised at the edge of stability. (англ.) // Annu Rev Microbiol. : journal. — 2003. — Vol. 57. — P. 125—154. — doi:10.1146/annurev.micro.57.012903.074300. — PMID 14527275.
- Hale C. A., de Boer PA. Direct binding of FtsZ to ZipA, an essential component of the septal ring structure that mediates cell division in E. coli. (англ.) // Cell : journal. — Cell Press, 1997. — Vol. 88. — P. 175—185. — doi:10.1016/S0092-8674(00)81838-3. — PMID 9008158.
- Pichoff S., Lutkenhaus J. Unique and overlapping roles for ZipA and FtsA in septal ring assembly in Escherichia coli. (англ.) // EMBO J. : journal. — 2002. — Vol. 21. — P. 685—693. — doi:10.1093/emboj/21.4.685. — PMID 11847116.
- Ebersbach G., Galli E., Møller-Jensen J., Löwe J., Gerdes K. Novel coiled-coil cell division factor ZapB stimulates Z ring assembly and cell division. (англ.) // Microbiology : journal. — Microbiology Society, 2008. — Vol. 68. — P. 720—735. — doi:10.1111/j.1365-2958.2008.06190.x. — PMID 18394147.
- Aarsman M. E., Piette A., Fraipont C., Vinkenvleugel T. M., Nguyen-Distèche M., den Blaauwen T. Maturation of the Escherichia coli divisome occurs in two steps. (англ.) // Microbiology : journal. — Microbiology Society, 2005. — Vol. 55. — P. 1631—1645. — doi:10.1111/j.1365-2958.2005.04502.x. — PMID 15752189.