Нуклеоид
Нуклео́ид (англ. Nucleoid) — неправильной формы зона в цитоплазме прокариотической клетки, в которой находится геномная ДНК и ассоциированные с ней белки. На долю ДНК приходится около 60 % массы нуклеоида; помимо ДНК, нуклеоид содержит РНК и белки[1]. Белки нуклеоида, которые обеспечивают пространственную организацию геномной ДНК, называют нуклеоидными белками или нуклеоид-ассоциированными белками; они не имеют ничего общего с гистонами, упаковывающими ДНК у эукариот. В отличие от гистонов, ДНК-связывающие белки нуклеоида не формируют нуклеосомы и обеспечивают компактизацию геномной ДНК другим способом[2]. Несмотря на аморфную форму, отдельные гены располагаются в нём упорядоченно[3].
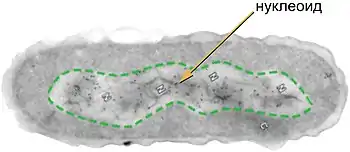
Бактерии
Бактериальные хромосомы
В большинстве случаев геном бактерии представлен кольцевой молекулой ДНК, которую также называют хромосомой. Как правило, бактериальная хромосома имеет длину около 1 мм, она занимает до 20 % цитоплазмы и имеет объём около 0,2 мкм³. Репликация геномной ДНК начинается от участка инициации (oriC), из которого две репликативные вилки движутся в противоположных направлениях и встречаются в сайте терминации (Ter), в котором далее происходит размыкание сцепленных дочерних хромосом. Количество хромосом в бактериальной клетке зависит не только от видовой принадлежности, но и от фазы развития популяции. В качестве бактерий, постоянно имеющих несколько хромосом, можно привести Deinococcus radiodurans (от 4 до 10 хромосом), Borrelia hermsii (от 8 до 16 хромосом), Desulfovibrio gigas (от 9 до 17 хромосом), Azotobacter vinelandii (до 80 хромосом). Молодые клетки обычно содержат больше хромосом, чем старые. Иногда хромосомы представляют не просто копии геномной ДНК: в некоторых случаях геном распределяется между несколькими неодинаковыми хромосомами и внехромосомными элементами (плазмидами). Так, у Agrobacterium tumefaciens, Brucella melitensis и Rhodobacter sphaeroides имеются две разные хромосомы, а у Bacillus cereus, Leptospira interrogans и Rhizobium meliloti кроме хромосом есть одна или две мегаплазмиды, состоящие из 100—500 тысяч пар оснований (п. о.)[4].
Ряд бактерий имеют и кольцевые, и линейные хромосомы, а некоторые — только линейные. Например, линейная хромосома имеется у возбудителя болезни Лайма — Borrelia burgdorferi. Известны и линейные плазмиды. Роль теломер при этом могут выполнять или одноцепочечные концевые шпильки, или особые белки, ковалентно связанные с концами линейной хромосомы или плазмиды; в некоторых случаях на время репликации линейная хромосома становится кольцевой[5].
ДНК-связывающие белки
За компактизацию бактериальной хромосомы отвечают несколько разнородных белков, из которых наиболее важны HU, H-NS и SMC. Белки HU и H-NS называют гистоноподобными, их взаимодействие с ДНК зависит от её последовательности, наличия в молекуле шпилек и изгибов. Выявлены также минорные гистоноподобные белки FIS и IHF, которые, наряду с HU и H-NS, не только обеспечивают пространственную организацию бактериальной геномной ДНК, но также влияют на её репликацию, рекомбинацию и экспрессию генов. HU, совместно с ДНК-гиразой, обеспечивает отрицательную сверхспирализацию ДНК. H-NS компактизует ДНК, влияет на её сверхспирализацию, вызывает в ней изгибы, однако по большей части он задействован в экспрессии генов: под его контролем находятся до 40 генов. Белки SMC имеются у представителей всех трёх доменов жизни. Они гомологичны миозину, у эукариот они играют роль моторов при конденсации хроматина[6].
Таким образом, можно выделить следующие уровни компактизации бактериальной ДНК. Первый уровень обеспечивается гистоноподобными белками HU и H-NS. Второй уровень — отрицательная сверхспирализация — обеспечивается SMC-белками и топоизомеразами. Наконец, высший уровень компактизации представлен независимо сверхспирализованными петлями (доменами), на которые разделена бактериальная хромосома. В состав каждого домена входит около 10 тысяч п. о[7].
Археи
У архей компактные нуклеоиды располагаются в центре клетки, как и у бактерий, и ориентированы вдоль её продольной оси. Число хромосом зависит от видовой принадлежности и фазы развития популяции, кроме того, в большинстве случаев в молодых клетках содержится больше хромосом. Так, у Methanocaldococcus jannaschii в стационарной фазе роста имеется от одной до пяти хромосом, а в фазе экспоненциального роста — 3—15 хромосом. Как и бактерии, археи могут иметь плазмиды[8].
У многих архей, например, у эвриархеот, нижний уровень компактизации ДНК обеспечивается белками, гомологичными гистонам эукариот. Гистоны архей по размеру уступают гистонам эукариот и имеют укороченный N-концевой участок, поэтому взаимодействуют с ДНК несколько иначе. Нуклеосомы архей не имеют октамерной структуры в силу отсутствия гистонов H2A и H2B, однако они похожи на тетрасому (H3/H4)2. Кренархеоты, ведущие термофильный и гипертермофильный образ жизни, не имеют гистонов, но у них имеются негистоновые ДНК-связывающие белки. Например, один из них, Sul7D, взаимодействует с малой бороздкой ДНК и защищает её от тепловой денатурации[9].
В отличие от бактерий, геномная ДНК архей характеризуется положительной сверхспирализацией, которая, как полагают, стабилизирует ДНК у экстремофильных видов. Положительную сверхспирализацию обеспечивает так называемая «обратная» гираза[10].
Примечания
- Нетрусов, Котова, 2012, с. 37—38.
- Wang W., Li G. W., Chen C., Xie X. S., Zhuang X. Chromosome organization by a nucleoid-associated protein in live bacteria. (англ.) // Science (New York, N.Y.). — 2011. — 9 September (vol. 333, no. 6048). — P. 1445—1449. — doi:10.1126/science.1204697. — PMID 21903814.
- Кассимерис, Лингаппа, Плоппер, 2016, с. 945.
- Пиневич, 2006, с. 194—195.
- Пиневич, 2006, с. 195.
- Пиневич, 2006, с. 196—197.
- Пиневич, 2006, с. 197—198.
- Пиневич, 2006, с. 198.
- Пиневич, 2006, с. 198—199.
- Пиневич, 2006, с. 199.
Литература
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т.. — СПб.: Издательство С.-Петербургского университета, 2006. — Т. I. — 352 с. — ISBN 5-288-04057-5.
- Нетрусов А. И., Котова И. Б. Микробиология. — 4-е изд., перераб. и доп.. — М.: Издательский центр «Академия», 2012. — 384 с. — ISBN 978-5-7695-7979-0.
- Кассимерис Л., Лингаппа В. Р., Плоппер Д. . Клетки по Льюину. — М.: Лаборатория знаний, 2016. — 1056 с. — ISBN 978-5-906828-23-1.