Плотные контакты
Пло́тные конта́кты (англ. tight junctions) — запирающие межклеточные контакты, присущие клеткам позвоночных животных, в составе которых мембраны соседних клеток максимально сближены и «сшиты» специализированными белками клаудинами и окклюдинами. Распространены в эпителиальных тканях, где составляют наиболее апикальную часть (лат. zonula occludens) комплекса контактов между клетками, в который входят адгезионные контакты и десмосомы. Плотные контакты построены из нескольких лент, опоясывающих клетку, которые, пересекаясь между собой, образуют сетевидную связь. С цитоплазматической стороны ассоциированы с актиновыми филаментами[1][2].
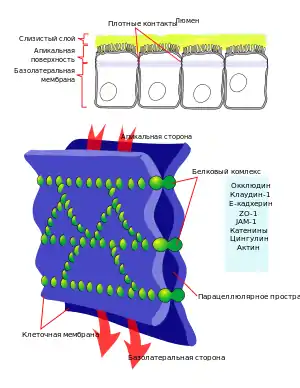
Эпителиальные ткани выполняют барьерную и транспортную функции, для этого они должны быть способны пропускать одни вещества и задерживать другие. Такую выборочную проницаемость успешно обеспечивают клеточные мембраны, однако между клетками остаются промежутки, через которые может проходить так называемый парацеллюлярный (параклеточный) транспорт. Роль плотных контактов заключается в том, чтобы ограничивать и регулировать параклеточную диффузию: они предотвращают протекание тканевой жидкости через эпителий, но при необходимости могут быть проницаемыми для ионов, небольших гидрофильных молекул и даже макромолекул. Также плотные контакты выполняют так называемую функцию «ограждения», они предотвращают диффузию компонентов мембраны в её внешнем слое, благодаря чему поддерживается разница в составе апикальной и базолатеральной мембран. Плотные контакты задействованы в сигнальных путях, регулирующих пролиферацию, поляризацию и дифференциацию эпителиальных клеток[3].
Аналогом плотных контактов у беспозвоночных являются септированные контакты[1].
Строение и молекулярный состав

Плотные контакты состоят из тонких лент, пересекающихся между собой, которые полностью опоясывают клетку и контактируют с аналогичными лентами на соседних клетках. На электронных микрофотографиях заметно, что в участках плотных контактов мембраны соприкасаются одна с другой или даже сливаются. Комбинация метода замораживания-скалывания с электронной микроскопией с высоким разрешением позволила установить, что плёнки плотных контактов построены из белковых частиц диаметром 3—4 нм, которые выступают с обеих поверхностей мембраны. Также в пользу того, что в образовании плотных контактов ключевую роль играют белки, свидетельствует разделение клеток под действием протеолитического фермента трипсина[2].
Всего в состав плотных контактов входит около 40 различных белков, как мембранных, так и цитоплазматических. Последние необходимы для прикрепления актиновых филаментов, регуляции и передачи сигнала[3].
Мембранные белки
Мембранные белки плотных контактов можно разделить на две группы: те, которые пересекают мембрану 4 раза, и те, которые пересекают её только раз. Первая группа очень обширная, в неё входят белки клаудины, окклюдины и трицеллюлин. Они имеют общие черты строения, в частности, в них имеются четыре α-спиральных трасмембранных домена, N- и С-концы обращены к цитозолю, а домены, выступающие в межклеточное пространство, участвуют в гомо- или гетерофильных взаимодействиях с подобными белками на соседней клетке[3].
Основными белками плотных контактов являются клаудины. Их роль была продемонстрирована на примере мышей с нокаутом гена клаудина-1, — в эпидермисе таких животных не формируются плотные контакты и они погибают в течение дня после рождения из-за обезвоживания вследствие интенсивного испарения[1]. Клаудины также участвуют в формировании селективных каналов для транспорта ионов. В геноме человека есть гены по крайней мере 24 различных клаудинов, экспрессия которых происходит тканеспецифично[3].
Второе место по распространенности в плотных контактах занимают белки окклюдины (от лат. occludo — закрывать), они регулируют транспорт маленьких гидрофильных молекул и прохождение нейтрофилов через эпителий[3]. Наибольшие концентрации третьего белка — трицеллюлина — наблюдаются в местах контакта трех клеток[1].
К белкам плотных контактов, пересекающим мембрану один раз, относятся JAM-A, -B, -C и -D (англ. junctional adhesion molecules) и родственные им CAR (англ. coxsackievirus and adenovirus receptor), CLMP (англ. CAR-like membrane protein) и ESAM (англ. endothelial-cell selective adhesion molecule), имеющие по два иммуноглобулиновых домена, а также белки CRB3 (англ. Crumbs homologue 3) и BVES[3].
Цитоплазматические белки
Цитоплазматическая часть плотных контактов необходима для их присоединения к актиновым филаментам, регуляции сцепления клеток и параклеточного транспорта, а также для передачи сигналов от поверхности внутрь клетки. В её состав входят адаптерные, каркасные и цитоскелетные белки, а также элементы сигнальных путей (киназы, фосфатазы). Наиболее изучен белок цитоплазматической пластинки ZO-1, он имеет несколько доменов, участвующих в белок-белковых взаимодействиях, каждый из которых обеспечивает контакт с другими компонентами. Так, три PDZ-домена взаимодействуют с клаудинами и другими адаптерными белками — ZO-2 и ZO-3, GUK-домен (англ. guanylate kinase homology) — с окклюдинами, а SH3-домен — с сигнальными белками[3].
С цитоплазматической стороной плотных контактов также ассоциированы комплексы белков PARD3/PAR6 и Pals1/PATJ, необходимые для установления полярности клеток и морфогенеза эпителия[3].
Функции
Первые исследования функций плотных контактов привели к представлению, что это статические непроницаемые структуры, необходимые для того, чтобы ограничить диффузию веществ между клетками. Впоследствии было выяснено, что они избирательно проницаемы, к тому же, их пропускная способность отличается в различных тканях и может регулироваться[4]. Также установлена ещё одна функция плотных контактов: роль в поддержании полярности клеток путём ограничения диффузии липидов и белков во внешнем слое плазматической мембраны. В первом десятилетии XXI века также накоплены данные, свидетельствующие об участии этих структур в сигнальных путях, в частности, регулирующих пролиферацию и полярность клеток[3].
Регулирование парацеллюлярного транспорта
Непроницаемость плотных контактов для большинства водорастворимых соединений может быть продемонстрирована в опыте по введению гидроксида лантана (электронно-плотный коллоидный раствор) в кровеносные сосуды поджелудочной железы. Через несколько минут после инъекции ацинарные клетки фиксируются, и из них готовятся препараты для микроскопии. В таком случае можно наблюдать, что гидроксид лантана диффундирует из крови в пространство между латеральными поверхностями клеток, но не может проникнуть через плотные контакты в их верхней части[2]. Другие опыты показали, что плотные контакты также непроницаемы для солей. Например, при выращивании культуры клеток почек собаки MDCK (англ. Madin-Darby canine kidney) в среде с очень низкой концентрацией кальция клетки формируют монослой, однако не связаны друг с другом плотными контактами. Через такой монослой могут свободно двигаться соли и жидкости. Если культуре добавить кальция, то за час формируются плотные контакты, и слой становится непроницаемым для жидкостей[2].
Однако не во всех тканях плотные контакты полностью непроницаемы, существуют так называемые неплотные эпителии (англ. leaky epithelia). Например, эпителий тонкого кишечника пропускает в 1000 раз больше ионов Na+, чем эпителий канальцев почек. Ионы проникают через параклеточные поры диаметром 4 Å, селективные по заряду и размеру частиц, которые формируются белками клаудинами[4]. Поскольку эпителии различных органов эксрессируют разные наборы клаудинов, то отличается и их проницаемость для ионов. Например, специфический клаудин, присутствуюий только в почках, позволяет проходить ионам магния в процессе реабсорбции[1].
Межклеточное пространство эпителия может быть проницаемым и для больших частиц, например, при повторении упомянутого опыта с гидроксидом лантана на ткани эпителия тонкого кишечника кролика можно наблюдать прохождение коллоидных частиц между клетками. Крупные молекулы транспортируются через специальные пути утечки (англ. leak pathway) диаметром более 60 Å[4]. Это важно, например, для процессов всасывания аминокислот и моносахаридов, концентрация которых в тонком кишечнике возрастает после еды достаточно для их пассивного транспорта[1].
Поддержание различия между апикальной и базолатеральной мембранами
Если в среду, контактирующую с апикальной частью монослоя MDCK-клеток, добавить липосомы, содержащие флуоресцентно меченные гликопротеины, некоторые из них спонтанно сливаются с клеточными мембранами. После этого флуоресценцию можно обнаружить в апикальной, но не в базолатеральной части клеток при условии целостности плотных контактов. Если же их разрушить, удалив из среды кальций, флуоресцентные белки диффундируют и равномерно распределяются по всей поверхности клетки[2].
Цитозольный слой мембраны имеет одинаковый липидный состав, как в апикальном, так и в базолатеральном участках, эти липиды могут свободно диффундировать. С другой стороны, липиды внеклеточного слоя двух частей клетки существенно различаются, и обмену между ними препятствуют плотные контакты. Например, все гликолипиды, как и белки, заякоренные гликозилфосфатидилинозитолом, в мембранах MDCK клеток расположены исключительно в внеклеточном слое апикальной части, а фосфатидилхолин — почти исключительно в базолатеральной части[2].
Болезни, связанные с плотными контактами
С нарушением формирования плотных контактов связаны некоторые наследственные заболевания человека, например, мутации в генах клаудина-16 и клаудина-19, которые приводят к гипомагниемии вследствие чрезмерной потери магния с мочой. Мутации в генах клаудина-13 и трицеллюлина вызывают наследственную глухоту. Дисрегуляция некоторых белков плотных контактов связана с онкологическими заболеваниями, например, экспрессия ZO-1 и ZO-2 снижается во многих типах рака. Компоненты плотных контактов также могут быть мишенями для онкогенных вирусов[3].
Некоторые вирусы используют мембранные белки плотных контактов для проникновения в клетку, в частности, клаудин-1 является корецептором для вируса гепатита C. Другие вирусы присоединяются к белкам плотных контактов, чтобы разрушить барьер, отделяющий их от настоящих рецепторов на базолатеральной слое эпителиальных клеток или неэпителиальных клетках[3].
Плотные контакты могут быть мишенью и для бактериальных патогенов, например, Clostridium perfringens — возбудитель газовой гангрены — выделяет энтеротоксин, действующий на внеклеточные домены клаудинов и окклюдинов, и вызывает протечки эпителия. Helicobacter pylori — возбудитель гастрита — вводит в клетки белок CagA, взаимодействующий с комплексом ZO-1-JAM-A; считается, что это помогает бактерии преодолеть защитный барьер желудочного эпителия[3].
Примечания
- Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. Molecular Biology of the Cell. — 5th. — Garland Science, 2007. — ISBN 978-0-8153-4105-5.
- Harvey Lodish et al. 15.7 Transport across Epithelia // Molecular Cell Biology. — 4th. — W H Freeman, 2000. — ISBN 0-7167-3136-3.
- Balda M. S., Matter K. Tight junctions at a glance. (англ.) // Journal Of Cell Science. — 2008. — 15 November (vol. 121, no. Pt 22). — P. 3677—3682. — doi:10.1242/jcs.023887. — PMID 18987354.
- Anderson J. M., Van Itallie C. M. Physiology and function of the tight junction. (англ.) // Cold Spring Harbor Perspectives In Biology. — 2009. — August (vol. 1, no. 2). — P. 002584—002584. — doi:10.1101/cshperspect.a002584. — PMID 20066090.
Литература
- Строение и молекулярный состав:
- Furuse M. Molecular basis of the core structure of tight junctions. (англ.) // Cold Spring Harbor Perspectives In Biology. — 2010. — January (vol. 2, no. 1). — P. 002907—002907. — doi:10.1101/cshperspect.a002907. — PMID 20182608.
- Krause G., Winkler L., Mueller S. L., Haseloff R. F., Piontek J., Blasig I. E. Structure and function of claudins. (англ.) // Biochimica Et Biophysica Acta. — 2008. — March (vol. 1778, no. 3). — P. 631—645. — doi:10.1016/j.bbamem.2007.10.018. — PMID 18036336.
- Физиология:
- Shen L., Weber C. R., Raleigh D. R., Yu D., Turner J. R. Tight junction pore and leak pathways: a dynamic duo. (англ.) // Annual Review Of Physiology. — 2011. — Vol. 73. — P. 283—309. — doi:10.1146/annurev-physiol-012110-142150. — PMID 20936941.
- Anderson J. M., Van Itallie C. M. Physiology and function of the tight junction. (англ.) // Cold Spring Harbor Perspectives In Biology. — 2009. — August (vol. 1, no. 2). — P. 002584—002584. — doi:10.1101/cshperspect.a002584. — PMID 20066090.
- Van Itallie C. M., Anderson J. M. Claudins and epithelial paracellular transport. (англ.) // Annual Review Of Physiology. — 2006. — Vol. 68. — P. 403—429. — doi:10.1146/annurev.physiol.68.040104.131404. — PMID 16460278.
- Регуляция:
- González-Mariscal L., Tapia R., Chamorro D. Crosstalk of tight junction components with signaling pathways. (англ.) // Biochimica Et Biophysica Acta. — 2008. — March (vol. 1778, no. 3). — P. 729—756. — doi:10.1016/j.bbamem.2007.08.018. — PMID 17950242.
- Tsukita S., Yamazaki Y., Katsuno T., Tamura A., Tsukita S. Tight junction-based epithelial microenvironment and cell proliferation. (англ.) // Oncogene. — 2008. — 24 November (vol. 27, no. 55). — P. 6930—6938. — doi:10.1038/onc.2008.344. — PMID 19029935.
- Патофизиология:
- Förster C. Tight junctions and the modulation of barrier function in disease. (англ.) // Histochemistry And Cell Biology. — 2008. — July (vol. 130, no. 1). — P. 55—70. — doi:10.1007/s00418-008-0424-9. — PMID 18415116.
- Turner J. R. Molecular basis of epithelial barrier regulation: from basic mechanisms to clinical application. (англ.) // The American Journal Of Pathology. — 2006. — December (vol. 169, no. 6). — P. 1901—1909. — doi:10.2353/ajpath.2006.060681. — PMID 17148655.
| В библиографических каталогах |
|---|