Корецептор
Кореце́птор (англ. Co-receptor) — находящийся на поверхности клетки дополнительный рецептор, который связывается с сигнальной молекулой (лигандом) в дополнение к первичному рецептору. Корецепторы Т-лимфоцитов усиливают взаимодействие T-клеточного рецептора с комплексом пептид (фрагмент антигена) + молекула главного комплекса гистосовместимости (MHC). Корецепторы нередко используются вирусами для проникновения в клетку.
Общая характеристика
Корецептор — это поверхностный рецептор, который после связывания с лигандом взаимодействует с соответствующим первичным рецептором и запускает сигнальный путь в клетке. В иммунологии и биохимии корецепторы также называют дополнительными рецепторами. У корецепторов, как правило, имеется большой внеклеточный домен, на который приходится от 76 % до 100 % всей молекулы рецептора. Именно внеклеточный домен отвечает за связывание с лигандом и формирование комплекса с первичным рецептором. В состав мотивов, отвечающих за эти функции, могут входить гликозаминогликаны, повторы EGF, остатки цистеина или домены ZP-1. Благодаря разнообразным мотивам во внеклеточным доменам корецепторы не только могут связываться с 2—9 различными лигандами, но и взаимодействовать с другими корецепторами. У большинства корецепторов нет внутриклеточного домена, и они связаны с клеточной мембраной посредством GPI-якоря, хотя у некоторых корецепторов всё же есть короткий цитоплазматический домен, лишённый киназной активности. Так как внутриклеточного домена у корецепторов чаще всего не бывает, то сами они не могут запускать сигнальные пути в клетке — это делают первичные рецепторы[1][2][3].
Лиганды, с которыми связываются корецепторы, могут быть самыми разнообразными: интерлейкины, нейротрофные факторы, фактор роста фибробластов (FGF), трансформирующий фактор роста (TGF), фактор роста сосудистого эндотелия (VEGF) и эпидермальный фактор роста (EGF). Корецепторы клеток тканей эмбриона играют важнейшую роль в образовании градиентов морфогенов, которые необходимы для правильной дифференцировки тканей. Корецепторы клеток эндотелия усиливают способность клеток к пролиферации и миграции. Кроме того, корецепторы играют важную роль в функционировании иммунной системы и часто используютя вирусами для проникновения в клетку[3][2].
Примеры
Семейство CD
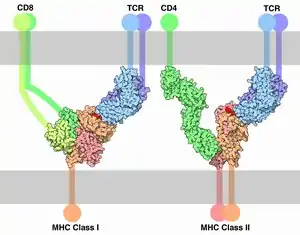
В семейство CD входят разнообразные рецепторы иммунных клеток[4]. Представители семейства, как правило, функционируют как корецепторы. Классическим примером может служить корецептор CD4, обеспечивающий взаимодействие T-хелпера с комплексом MHC II класса-пептид посредством T-клеточного рецептора (TCR). Корецепторы необходимы для активации иммунных клеток и программируемой клеточной гибели активированных иммунных клеток[5]. Показано, что блокада связывания корецептора CD4 с MHC-II приводит к тому, что активная T-клетка теряет возможность к программируемой гибели[6]. Корецептор CD4 состоит из четырёх иммуноглобулиноподобных доменов и заякорен в клеточной мембране посредством одного трансмембранного домена. Чаще всего рецепторы CD являются мономерами или димерами и выполняют широкий спектр функций. Упомянутый CD4 не только необходим для функционирования T-клеток, но и служит главным рецептором, необходимым для проникновения ВИЧ в клетку (CD4 взаимодействует с поверхностным гликопротеином GP120 ВИЧ). CD28 функционирует как костимулирующий корецептор, влияющий на взаимодействие с MHC-II с TCR и CD4. Хотя CD28 усиливает выделение интерлейкина 2 T-клетками при их активация, блокада этого корецептора никак не отражается на способности T-клеток подвергаться программируемой клеточной гибели[6].
Семейство CCR
Представители семейства CCR являются рецепторами, связанными с G-белками (GPCR), и функционируют как рецепторы хемокинов. Эти рецепторы располагаются преимущественно на иммунных клетках, в особенности, T-клетках[7]. Рецепторы CCR также экспрессируют некоторые клетки нервной системы, в частности, клетки микроглии[7]. Один из наиболее изученных представителей семейства — CCR5 (и его близкий родственник CXCR4), который функционирует как корецептор для ВИЧ[7][8]. Поверхностный гликопротеин GP120 ВИЧ взаимодействует со своим главным рецептором CD4, а CCR5 формирует комплекс с CD4 и ВИЧ, позволяя вирусу проникнуть внутрь клетки. Некоторые штаммы ВИЧ в качестве корецепторов могут также использовать близкие друг к другу CCR2b, CCR3 и CCR8, однако все штаммы ВИЧ могут использовать CCR5 и близкий к нему белок CXCR4 для проникновения в клетку. CCR5, кроме того, имеет сродство к воспалительному белку макрофагов (англ. macrophage inflammatory protein, MIP) и играет роль в развитии воспалительной реакции. Роль CCR5 в возникновении воспаления хуже понятна, чем его роль в проникновении ВИЧ в T-клетку[7][8].
Клиническое значение
Так как корецепторы играют важную роль в передаче сигналов между клетками, они задействованы в развитии многих заболеваний. Мыши, нокаутные по корецепторам, чаще всего нежизнеспособны, и нокаут корецепторов, как правило, является эмбриональной или перинатальной леталью[2].
Многие заболевания, связанные с корецепторами, вызываются мутациями в кодирующей области генов, кодирующих корецепторы. Например, мутации LRP5, служащего корецепторами для гликопротеинов пути Wnt, которые регулируют костную массу, приводят к пониженной костной массе и развитию остеопороза[9]. Некоторые случаи гигантизма могут быть связаны с корецептором, известным как глипикан 3[2].
Белок CEACAM1 (от англ. Carcinoembryonic antigen cell adhesion molecule-1) функционирует как корецептор, необходимый для адгезии эпителиальных, эндотелиальных и гемопоэтических клеток. Он связывается с VEGF и играет важную роль в формировании кровеносных сосудов[10]. Мыши, лишённые гена Caecam1, не имеют интенсивного роста сосудов, необходимого для питания опухолей, и образуют меньше оксида азота NO, поэтому Caecam1 может быть мишенью противораковой терапии. Важную роль в ангиогенезе в опухолях играют корецепторы семейства нейропилинов, связывающиеся с VEGF наряду с рецепторами VEGFR1/VEGFR2 и плексином[2].
Корецептор CD109 является отрицательным регулятором рецептора фактора роста опухолей TGF-β. При связывании с TGF-β рецептор интернализуется в составе везикул под действием CD109, который снижает передачу сигнала через него. Таким образом, корецептор может функционировать как регуляторная молекула, снижающая способность клеток к росту и миграции, которые необходимы для развития опухоли[11][2].
Корецепторы синдекан 1 и 4 задействованы в развитии разнообразных видов рака (рак шейки матки, молочной железы, лёгкого и толстой кишки), причём ненормальный уровень экспрессии этих белков соответствует худшему прогнозу[2].
Помимо описанных выше корецепторов CD4, CCR5 и CXCR4, необходимых для проникновения ВИЧ в T-лимфоцит, другие корецепторы также могут быть задействованы в проникновении вирусов в клетку-мишень. Так, для вируса гепатита C необходим корецептор CD81, кроме того, проникновение этого вируса в клетку может регулировать белок плотных контактов клаудин 1[12].
Блокада корецептора CD4 с помощью антител может использоваться для того, чтобы снизить уровень активации T-клеток при аутоиммунных заболеваниях[13].
Примечания
- Gomperts B. D., Kramer I. M., Tatham P. E. R. Signal transduction. — Academic Press. — 2002. — ISBN 0-12-289631-9.
- Kirkbride K. C., Ray B. N., Blobe G. C. Cell-surface co-receptors: emerging roles in signaling and human disease. (англ.) // Trends In Biochemical Sciences. — 2005. — November (vol. 30, no. 11). — P. 611—621. — doi:10.1016/j.tibs.2005.09.003. — PMID 16185874.
- Guo D., Jia Q., Song H. Y., Warren R. S., Donner D. B. Vascular endothelial cell growth factor promotes tyrosine phosphorylation of mediators of signal transduction that contain SH2 domains. Association with endothelial cell proliferation. (англ.) // The Journal Of Biological Chemistry. — 1995. — 24 March (vol. 270, no. 12). — P. 6729—6733. — doi:10.1074/jbc.270.12.6729. — PMID 7896817.
- Bobbitt K. R., Justement L. B. Regulation of MHC class II signal transduction by the B cell coreceptors CD19 and CD22. (англ.) // Journal Of Immunology (Baltimore, Md. : 1950). — 2000. — 15 November (vol. 165, no. 10). — P. 5588—5596. — doi:10.4049/jimmunol.165.10.5588. — PMID 11067914.
- Wang J. H., Meijers R., Xiong Y., Liu J. H., Sakihama T., Zhang R., Joachimiak A., Reinherz E. L. Crystal structure of the human CD4 N-terminal two-domain fragment complexed to a class II MHC molecule. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2001. — 11 September (vol. 98, no. 19). — P. 10799—10804. — doi:10.1073/pnas.191124098. — PMID 11535811.
- Boehme S. A., Zheng L., Lenardo M. J. Analysis of the CD4 coreceptor and activation-induced costimulatory molecules in antigen-mediated mature T lymphocyte death. (англ.) // Journal Of Immunology (Baltimore, Md. : 1950). — 1995. — 15 August (vol. 155, no. 4). — P. 1703—1712. — PMID 7636229.
- Berson J. F., Doms R. W. Structure-function studies of the HIV-1 coreceptors. (англ.) // Seminars In Immunology. — 1998. — June (vol. 10, no. 3). — P. 237—248. — doi:10.1006/smim.1998.0130. — PMID 9653050.
- Bleul C. C., Wu L., Hoxie J. A., Springer T. A., Mackay C. R. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1997. — 4 March (vol. 94, no. 5). — P. 1925—1930. — doi:10.1073/pnas.94.5.1925. — PMID 9050881.
- Sawakami K., Robling A. G., Ai M., Pitner N. D., Liu D., Warden S. J., Li J., Maye P., Rowe D. W., Duncan R. L., Warman M. L., Turner C. H. The Wnt co-receptor LRP5 is essential for skeletal mechanotransduction but not for the anabolic bone response to parathyroid hormone treatment. (англ.) // The Journal Of Biological Chemistry. — 2006. — 18 August (vol. 281, no. 33). — P. 23698—23711. — doi:10.1074/jbc.M601000200. — PMID 16790443.
- Nouvion A. L., Oubaha M., Leblanc S., Davis E. C., Jastrow H., Kammerer R., Breton V., Turbide C., Ergun S., Gratton J. P., Beauchemin N. CEACAM1: a key regulator of vascular permeability. (англ.) // Journal Of Cell Science. — 2010. — 15 December (vol. 123, no. Pt 24). — P. 4221—4230. — doi:10.1242/jcs.073635. — PMID 21081647.
- Bizet A. A., Liu K., Tran-Khanh N., Saksena A., Vorstenbosch J., Finnson K. W., Buschmann M. D., Philip A. The TGF-β co-receptor, CD109, promotes internalization and degradation of TGF-β receptors. (англ.) // Biochimica Et Biophysica Acta. — 2011. — May (vol. 1813, no. 5). — P. 742—753. — doi:10.1016/j.bbamcr.2011.01.028. — PMID 21295082.
- Evans M. J., von Hahn T., Tscherne D. M., Syder A. J., Panis M., Wölk B., Hatziioannou T., McKeating J. A., Bieniasz P. D., Rice C. M. Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry. (англ.) // Nature. — 2007. — 12 April (vol. 446, no. 7137). — P. 801—805. — doi:10.1038/nature05654. — PMID 17325668.
- Waldmann H., Adams E., Cobbold S. Reprogramming the immune system: co-receptor blockade as a paradigm for harnessing tolerance mechanisms. (англ.) // Immunological Reviews. — 2008. — June (vol. 223). — P. 361—370. — doi:10.1111/j.1600-065X.2008.00632.x. — PMID 18613847.