Адгезивные контакты
Адгези́вные конта́кты[1] (англ. adherens junctions, AJ) — якорные межклеточные контакты, ассоциированные с микрофиламентами, обеспечивающие целостность и механическую прочность ткани. Они противостоят растяжению, придают клеткам возможность координированно использовать актиновый цитоскелет. Адгезивные контакты относятся к гомофильным, то есть соединяют клетки одинакового типа. В их формировании принимают участие белки кадгерины и катенины[2][3].
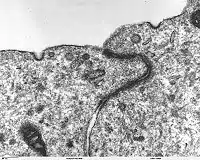
Морфологически адгезивные контакты являются относительно простыми, в отличие от десмосом, плотных и щелевых контактов, они не имеют высокоспециализированных ультраструктур, кроме скопления актиновых филаментов. От других типов соединений клеток они отличаются относительной гибкостью и изменчивостью[4].
Распространение
Чаще всего адгезионные контакты встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки поясок, который называют зоной прилипания (англ. zonula adherens). Такие зоны в эпителии позвоночных животных преимущественно размещаются базальнее участка плотных контактов (англ. zonula occludens) и апикальнее десмосом (англ. macula adherens)[2][3].
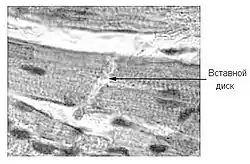
Однако распространение адгезивных контактов в организме не ограничено только эпителием: во многих неэпителиальных тканях, например, в нервной и соединительной, они присутствуют в форме точечных или полосковидных клеточных контактов. Также они хорошо выражены в сердечной мышце, где обеспечивают косвенное сообщение сократительного аппарата кардиомиоцитов. Вместе с десмосомами адгезивные контакты образуют так называемые вставные диски между клетками миокарда[2][4].
Строение
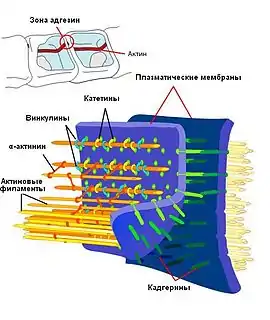
В зоне адгезивных контактов мембраны соседних клеток удалены друг от друга на расстояние 10—20 нм[4]. В состав адгезионных контактов входят три основных элемента:
- Собственно сообщение клеток происходит благодаря трансмембранным молекулам клеточной адгезии (англ. cell adhesion molecules, CAM), из которых самыми распространенными в адгезионных контактах являются белки кадгерины. Их N-концевая внеклеточная часть в присутствии ионов кальция взаимодействует с подобной молекулой на соседней клетке, обеспечивая слипание клеток, а внутриклеточный C-концевой домен связывается с якорными белками. Также в адгезивных контактах были обнаружены другие трансмембранные белки: нектины и везатины[5].
- Адаптером между CAM и элементами цитоскелета выступают внутриклеточные якорные белки, большинство из них крепят цитоплазматическую сторону кадгеринов к актиновым филаментам. Кадгерины непосредственно взаимодействуют с β-катенином и плакоглобином (γ-катенином), который, в свою очередь, присоединяется к молекулам α-катенина, а те — к винкулину, α-актинину или ZO-1, которые уже связываются с актином[3][5]. В комплексе якорных белков с кадгеринами присутствует также белок p120-катенин, который, вероятно, участвует в регуляции силы адгезионных контактов между клетками[2]. Нектины крепятся к актиновым филаментам через белок афадин[4][5].
- Третьим компонентом являются пучки актиновых филаментов в соседних клетках, которые косвенно соединены между собой. Также была продемонстрирована связь адгезивных контактов с микротрубочками, хотя значительного их скопления в этих участках не наблюдается. Микротрубочки могут присоединяться к адгезивнымконтактам как плюс-, так и минус-концами. Вместе с моторными белками они, вероятно, участвуют в транспортировке белков, необходимых для формирования контактов, в частности, кадгеринов[4].
Функции
Одной из основных функций адгезионных контактов является физическое соединение клеток в единую ткань, их ослабление часто приводит к диссоциации клеток. Такого эффекта можно достичь, обработав ткань или монослойную культуру хелатирующим агентами, такими как ЭДТА, связывающими ионы кальция, вследствие чего взаимодействие между кадгеринами нарушается. Однако хелатирующих агентов обычно недостаточно для полного разделения клеток, поскольку между ними существуют другие контакты, не зависящие от кальция[4].
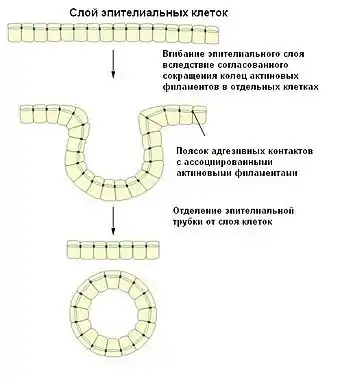
Адгезивные контакты обеспечивают образование широкой межклеточной сети из пучков сократительных актиновых филаментов, расположенных параллельно мембранам клеток и соединенных между собой с помощью белков катенинов и кадгеринов. Такая организация позволяет не только противостоять механическому напряжению, но и согласовывать поведение клеток во время процессов морфогенеза. Например, координированное сокращение колец актиновых филаментов соседних клеток является необходимым для формирования трубок из слоя эпителия, в частности, во время закладки нервной трубки[2]. Одним из примеров является Shroom3-зависимое сокращение зоны прилипания, при этом актин-связывающий белок Shroom3 привлекает к участку адгезионные контакты Rho-киназы и активирует миозин-II, в результате чего и происходит сокращение[4].
Адгезивные контакты также задействованы в межклеточной передаче сигналов. Об этом свидетельствует локализация в зрелых контактах рецепторной тирозинфосфатазы μ и белка RACK1, который взаимодействует с ней. При снижении экспрессии α-катенина наблюдается значительное увеличение темпов пролиферации эпителиальных клеток. Было показано, что за этот эффект отвечает сигнальный путь инсулин/MAPK[5].
Примечания
- Кассимерис Л., Лингаппа В. Р., Плоппер Д. . Клетки по Льюину. — М.: Лаборатория знаний, 2016. — С. 851. — 1056 с. — ISBN 978-5-906828-23-1.
- Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. Molecular Biology of the Cell. — 5th. — Garland Science, 2007. — ISBN 978-0-8153-4105-5.
- Harvey Lodish et al. 22.1 Cell-Cell Adhesion and Communication // Molecular Cell Biology. — 4th. — W H Freeman, 2000. — ISBN 0-7167-3136-3.
- Meng W., Takeichi M. Adherens junction: molecular architecture and regulation. (англ.) // Cold Spring Harbor Perspectives In Biology. — 2009. — December (vol. 1, no. 6). — P. 002899—002899. — doi:10.1101/cshperspect.a002899. — PMID 20457565.
- Nagafuchi A. Molecular architecture of adherens junctions. (англ.) // Current Opinion In Cell Biology. — 2001. — October (vol. 13, no. 5). — P. 600—603. — PMID 11544029.
