Пили
Пили́, или фи́мбрии, или ворси́нки[1] — нитевидные белковые структуры, расположенные на поверхности клеток многих бактерий. Размер пилей варьирует от долей мкм до более чем 20 мкм в длину и 2—11 нм в диаметре. Пили участвуют в передаче генетического материала между бактериальными клетками (конъюгация), прикреплении бактерий к субстрату и другим клеткам, отвечают за адаптацию организмов, служат местами прикрепления многих бактериофагов.
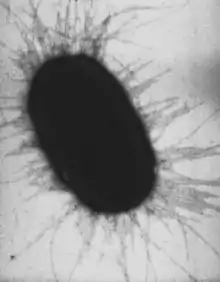
Структура
Пили состоят из одного или нескольких типов спирально уложенных белковых молекул, которые называют пилинами или фимбринами. Часто на концах пилей присутствуют особые прикрепительные субъединицы (адгезины), но иногда весь пиль обладает адгезивными свойствами. Структурно пили могут быть от тонких нитевидных образований до толстых палочкообразных структур с осевыми отверстиями. Очень тонкие пили (диаметром менее 2 нм) называют «кудряшками», они сливаются в пушистую липкую массу на поверхности клеток, которая обеспечивает их агрегацию. Пили-кудряшки являются амилоидными фибриллами, родственными белкам, формирующим амилоидные бляшки в мозге пациентов с болезнью Альцгеймера[2]. Часто пили разбросаны по всей поверхности клетки, но иногда локализуются только на одном её конце[3].
Функции
Основная функция большинства пилей заключается в обеспечении правильного позиционирования особых молекул, обеспечивающих адгезию клеток. Взаимодействие бактерий с другими про- и эукариотическими клетками, обеспечиваемое пилями, является важным этапом заселения эпителия и проникновения патогенных бактерий в клетки организма хозяина, образования биоплёнок и переноса генетического материала при конъюгации. Некоторые пили задействованы в движении бактериальных клеток. Пили иногда служат рецепторами бактериофагов. Например, при инфекции мочевыводящих путей, вызванной патогенными штаммами Escherichia coli, клетки прикрепляются к эпителию мочевого пузыря с помощью пилей, на конце которых находятся молекулы белка-адгезина FimH, который взаимодействует с остатками маннозы на поверхности эпителиальных клеток. Благодаря прикреплению к эпителию клетки бактерии не выводятся из организма с мочой. Пили являются важными факторами вирулентности многих кишечных бактерий, таких как Salmonella enterica, энтеропатогенные штаммы E. coli, Vibrio cholerae. Прикрепление пилей к эукариотическим клеткам может запускать в них определённые сигнальные каскады. Например, прикрепление пилей Neisseria к эпителиальным клеткам вызывает в последних выход ионов кальция в цитоплазму, при этом кальций является важным вторичным посредником в передаче сигнала в эукариотических клетках[4][5].
Образование
В образовании пилей, помимо самих белков-пилинов, участвуют дополнительные белки, способствующие правильной сборке. У грамотрицательных бактерий они должны пройти через клеточную мембрану, периплазматическое пространство и наружную мембрану. Ключевую роль в сборке пилей играет периплазматический шаперон PapD и транспортный белок внешней мембраны, известный как Usher-белок. PapD связывается с пилинами, вышедшими в периплазму, и доставляет их к Usher-белку, после чего высвобождается назад в периплазму. Доставленные к Usher-белку пилины приобретают активированную конформацию и далее собираются в пили[6].
Пили-кудряшки образуются по специфическому, тщательно регулируемому механизму. У E. coli в их сборке задействованы белковые продукты оперонов csgBA и csgDEFG. Фибрилла кудряшки образована белком CsgA, который, наряду с минорным белком CsgB, участвует в фазе нуклеации фибриллы. Когда нуклеация пройдена и начинается рост ворсинки, CsgB включается в неё. Фибриллы CsgA очень устойчивы и разрушаются лишь под действием 75 %-ной муравьиной кислоты. Оперон csgDEFG кодирует транскрипционный фактор CsgD и три предполагаемых фактора сборки пилей[2].
Классификация
Традиционно пили подразделяют на 4 типа.
- Пили I типа связываются с остатками маннозы на других клетках;
- Пили II типа независимы от маннозы и не вызывают гемагглютинацию;
- Пили III типа вызывают агглюцинацию эритроцитов человека, обработанных таниновой кислотой;
- Пили IV типа также известны как половые пили, они участвуют в процессе конъюгации[7].
Некоторые учёные подразделяют пили на пили главного типа и альтернативного типа. Пили главного типа включают пили типов I—III по традиционной классификации. Они располагаются по всей поверхности клетки и прикрепляются непосредственно к её наружной поверхности. Пили главного типа выполняют только адгезионные функции. Пили альтернативного типа (тип IV по традиционной классификации) образуются в количестве 1—2 на клетку и располагаются на одном или двух её полюсах. Они, подобно жгутикам, имеют базальное тело и заякорены в клеточной мембране, а потому насквозь проходят периплазму и наружную мембрану. Пили альтернативного типа участвуют не только в адгезии. Они могут быстро разбираться от основания, при этом дистальные их концы остаются прикреплёнными к субстрату или другой клетке. За счёт этого одна клетка может подтягиваться ближе к другой, что особенно важно при конъюгации[8].
Примечания
- Нетрусов, Котова, 2012, с. 54.
- Кассимерис, Лингаппа, Плоппер, 2016, с. 940.
- Нетрусов, Котова, 2012, с. 54—56.
- Нетрусов, Котова, 2012, с. 56—58.
- Кассимерис, Лингаппа, Плоппер, 2016, с. 939.
- Кассимерис, Лингаппа, Плоппер, 2016, с. 939—940.
- Пиневич, 2006, с. 338.
- Пиневич, 2006, с. 338—340.
Литература
- Кассимерис Л., Лингаппа В. Р., Плоппер Д. . Клетки по Льюину. — М.: Лаборатория знаний, 2016. — 1056 с. — ISBN 978-5-906828-23-1.
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб.: Издательство С.-Петербургского университета, 2006. — Т. I. — 352 с. — ISBN 5-288-04057-5.
- Нетрусов А. И., Котова И. Б. Микробиология. — 4-е изд., перераб. и доп. — М.: Издательский центр «Академия», 2012. — 384 с. — ISBN 978-5-7695-7979-0.
- Современная микробиология / Под ред. Й. Ленгелера, Г. Древса, Г. Шлегеля. — М.: Мир, 2005. — Т. 1. — 654 с.
| Клеточная оболочка |
|
|---|---|
| Наружная оболочка | |
| Форма |
|
