Внешняя бактериальная мембрана
Вне́шняя бактериа́льная мембра́на, или нару́жная бактериа́льная мембра́на (англ. bacterial outer membrane) — биологическая мембрана, располагающаяся поверх слоя пептидогликана у грамотрицательных бактерий. По составу она отличается от внутренней, клеточной мембраны. На её поверхности находятся липополисахариды, являющиеся антигенами грамотрицательных патогенных бактерий.
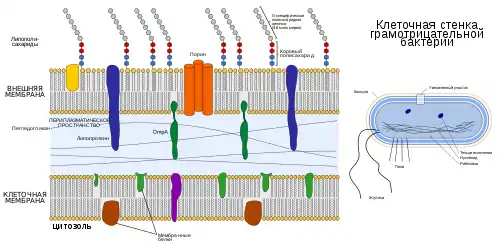
Состав

Внешняя мембрана, как и клеточная, имеет толщину 7—8 нм[1]. Подобно клеточной мембране, внешняя мембрана представляет собой липидный бислой, который состоит из фосфолипидов, преимущественно фосфатидилэтаноламина, фосфатидилглицерола и дифосфатидилглицерола. Однако во внешней мембране фосфолипиды распределены между слоями несимметрично: внешний лист мембраны образован преимущественно липополисахаридами. Исключениями являются цианобактерии и виды рода Neisseria, у которых во внешнем слое мембраны одновременно присутствуют фосфолипиды и липоолигосахариды. В структуре липополисахарида можно выделить четыре структурных блока:
- липид A;
- внутренний олигосахаридный кор, включающий две молекулы октозы и трёх молекул гептозы;
- олигосахаридного наружного кора, состоящего из пяти молекул гептозы;
- полисахаридной наружной цепи, состоящей из повторяющихся олигосахаридных звеньев (антигена O), каждое из которых состоит из 2—8 молекул гексоз[2].
Внешняя мембрана богата белками, их обозначают Omp (от англ. outer membrane protein). Белки могут покрывать до половины поверхности внешней мембраны. Белки Omp подразделяют на главные и минорные. Главные Omp-белки синтезируются постоянно; к их числу относятся липопротеин Брауна, липопротеины спирохет, OmpA и неспецифический порин OmpF[3].
В структурно-функциональном отношении все белки внешней мембраны можно подразделить на три группы:
- амфипатические липопротеины, на N-конце которых находится липидный домен. Они связывают внешнюю мембрану с слоем пептидогликана;
- интегральные структурные белки (например, OmpA);
- интегральные белки, формирующие гидрофильные каналы[3].
Иногда на препаратах обнаруживаются зоны контакта клеточной и внешней мембран. Такие участки называют контактами Байера. Вероятно, непосредственного контакта между мембранами не происходят, и они связываются друг с другом специальными белками в образующейся на месте контакте бреши в пептидогликановом слое[4].
Функции
Во внешней мембране имеются постоянно открытые ионные каналы, из-за чего на ней не может поддерживаться постоянный электрохимический градиент ионов натрия или протонов, поэтому внешняя мембрана не принимает участия в метаболизме клетки. Она также не имеет отношения к биосинтезу белков, липидов и полисахаридов, хотя может быть задействована в их секреции. Однако во внешней мембране находятся некоторые ферменты — пермеазы, гидролазы, иногда — окислительные ферменты, такие как марганец-оксидаза[1].
Функции внешней мембраны в бактериальной клетке очень разнообразны. Она совместно с клеточной мембраной образует периплазматическое пространство (периплазму), наряду с клеточной стенкой придаёт жёсткость клетке, служит фильтром, который не даёт попасть в периплазму крупным гидрофильным молекулам и задерживает гидрофобные молекулы. Она не даёт покинуть клетку многим ферментам периплазмы, участвует в поступлении в клетку питательных веществ, а также выделении наружу антибиотиков, токсинов, метаболитов и разнообразных белков. Внешняя мембрана опосредует неспецифичную адгезию бактериальных клеток, взаимодействует с бактериофагами, поверхностными рецепторами как прокариотических, так и эукариотических клеток, а также антителами. Благодаря внешней мембране жёлчные кислоты и другие амфифильные детергенты, а также антибиотики действуют на грамотрицательные бактерии слабее, чем на грамположительные[5]. Липоолигосахариды — важнейшие антигены патогенных грамотрицательных бактерий, их также называют эндотоксинами[3].
Биогенез
Механизмы, которые обеспечивают доставку компонентов внешней мембраны на поверхность клетки, до конца не ясны. Компоненты липополисахарида — липид A и O-антигеновые повторяющиеся единицы — синтезируются на цитоплазматической стороне клеточной мембраны и доставляются наружу независимо двумя специализированными транспортными системами, а именно, переносчиком O-антигена Wzx (RfbX) и ABC-транспортера MsbA, который перемещает липид A с внутреннего липидного слоя клеточной мембраны в наружный[6][7][8][9][10]. Полимеризация единиц O-антигена происходит в периплазматическом пространстве специализированной полимеразой Wzy, и полимерный фрагмент далее присоединяется к коровому липиду A лигазой WaaL, образуя липополисахарид[11][12]. Аппарат переноса молекул липополисахарида наружу клетки состоит из белков LptA, LptB, LptC, LptD, LptE. Для пяти из них удалось установить, в каких частях клетки они находятся, что может помочь разобраться в том, как функционирует аппарат сборки и выделения молекул липополисахарида[12]. Известно, что LptC переносит липополисахарид с клеточной мембраны во внешнюю[12]. LptE формирует комплекс с LptD, который обеспечивает встраивание молекул липополисахарида во внешнюю мембрану[12][13][14].
Везикулы наружной мембраны
От наружной мембраны могут отпочковываться везикулы (везикулы бактериальной внешней мембраны) диаметром от 20 до 500 нм. Образование везикул может быть связано с ростом бактериальной клетки, они могут служить средством доставки ферментов и прочих белков, например, патогенные бактерии могут транспортировать в составе везикул факторы вирулентности. Например, у Pseudomonas aeruginosa в составе везикул наружной мембраны в числе прочих белков выделяется β-лактамаза, разрушающая пенициллин[15].
См. также
Примечания
- Пиневич, 2006, с. 240.
- Пиневич, 2006, с. 241.
- Пиневич, 2006, с. 250.
- Пиневич, 2006, с. 257.
- Пиневич, 2006, с. 240—241.
- Feldman M. F., Marolda C. L., Monteiro M. A., Perry M. B., Parodi A. J., Valvano M. A. The activity of a putative polyisoprenol-linked sugar translocase (Wzx) involved in Escherichia coli O antigen assembly is independent of the chemical structure of the O repeat. (англ.) // The Journal Of Biological Chemistry. — 1999. — 3 December (vol. 274, no. 49). — P. 35129—35138. — PMID 10574995.
- Liu D., Cole R. A., Reeves P. R. An O-antigen processing function for Wzx (RfbX): a promising candidate for O-unit flippase. (англ.) // Journal Of Bacteriology. — 1996. — April (vol. 178, no. 7). — P. 2102—2107. — PMID 8606190.
- Doerrler W. T., Reedy M. C., Raetz C. R. An Escherichia coli mutant defective in lipid export. (англ.) // The Journal Of Biological Chemistry. — 2001. — 13 April (vol. 276, no. 15). — P. 11461—11464. — doi:10.1074/jbc.C100091200. — PMID 11278265.
- Polissi A., Georgopoulos C. Mutational analysis and properties of the msbA gene of Escherichia coli, coding for an essential ABC family transporter. (англ.) // Molecular Microbiology. — 1996. — June (vol. 20, no. 6). — P. 1221—1233. — PMID 8809774.
- Zhou Z., White K. A., Polissi A., Georgopoulos C., Raetz C. R. Function of Escherichia coli MsbA, an essential ABC family transporter, in lipid A and phospholipid biosynthesis. (англ.) // The Journal Of Biological Chemistry. — 1998. — 15 May (vol. 273, no. 20). — P. 12466—12475. — PMID 9575204.
- Raetz C. R., Whitfield C. Lipopolysaccharide endotoxins. (англ.) // Annual Review Of Biochemistry. — 2002. — Vol. 71. — P. 635—700. — doi:10.1146/annurev.biochem.71.110601.135414. — PMID 12045108.
- Sperandeo P., Lau F. K., Carpentieri A., De Castro C., Molinaro A., Dehò G., Silhavy T. J., Polissi A. Functional analysis of the protein machinery required for transport of lipopolysaccharide to the outer membrane of Escherichia coli. (англ.) // Journal Of Bacteriology. — 2008. — July (vol. 190, no. 13). — P. 4460—4469. — doi:10.1128/JB.00270-08. — PMID 18424520.
- Wu T., McCandlish A. C., Gronenberg L. S., Chng S. S., Silhavy T. J., Kahne D. Identification of a protein complex that assembles lipopolysaccharide in the outer membrane of Escherichia coli. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2006. — 1 August (vol. 103, no. 31). — P. 11754—11759. — doi:10.1073/pnas.0604744103. — PMID 16861298.
- Bos M. P., Tefsen B., Geurtsen J., Tommassen J. Identification of an outer membrane protein required for the transport of lipopolysaccharide to the bacterial cell surface. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2004. — 22 June (vol. 101, no. 25). — P. 9417—9422. — doi:10.1073/pnas.0402340101. — PMID 15192148.
- Пиневич, 2006, с. 256.
Литература
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб.: Издательство С.-Петербургского университета, 2006. — Т. I. — 352 с. — ISBN 5-288-04057-5.