Чешуекрылые
Чешуекры́лые, или ба́бочки (лат. Lepidóptera, от др.-греч. λεπίς, род. п. λεπίδος — чешуя и πτερόν — крыло), — отряд насекомых с полным превращением, наиболее характерная особенность представителей которого — наличие густого покрова хитиновых чешуек (уплощённых волосков) на передних и задних крыльях (при этом чешуйки расположены как на жилках, так и на крыловой пластинке между ними). Для большинства видов характерен специализированный сосущий ротовой аппарат с хоботком, образованным удлинёнными лопастями нижней челюсти. Форма и размах крыльев весьма разнообразны: от 2 мм[1] до 28 см[2][3].
| Чешуекрылые | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Отряд: Чешуекрылые |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Lepidoptera Linnaeus, 1758 | |||||||||||||||||||||||||||||||||||||||||
| Подотряды | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 190 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Развитие — с полным превращением: имеются стадии яйца, личинки (называемой гусеницей), куколки и имаго. Личинка червеобразная, с недоразвитыми брюшными ногами, мощно склеротизованными покровами головы, грызущим ротовым аппаратом и парными шёлкоотделительными железами, выделения из которых, при соприкосновении с воздухом, образуют шёлковую нить[4].
Чешуекрылые, ископаемые остатки которых известны начиная с юрского периода, в настоящее время являются одним из наиболее богатых видами отрядов насекомых — в отряде насчитывается более 158 000 видов. Представители отряда распространены на всех континентах, за исключением Антарктиды.
Раздел энтомологии, изучающий чешуекрылых, называется лепидоптерологией.
Этимология названий

Латинское название отряда, Lepidoptera, происходит от др.-греч. λεπίς, род. п. λεπίδος («чешуя») и πτερόν («крыло»). Такое название отряда связано с тем, что крылья его представителей покрыты чешуйками, представляющими собой видоизменённые щетинки.
Общеупотребительное русское название представителей этого отряда — «бабочка» — восходит к праслав. *babъka (< *baba; «старуха, бабка») и представлениям об этих насекомых, как о душах умерших[5][6]. И по сей день во многих сёлах и деревнях их называют «бабуля», «бабушка», «бабучка», «бабурка», «бабка».
Слово «мотылёк», которым называют некоторых представителей отряда, восходит к праслав. *motyl'ь (ср. чеш. motýl, польск. motyl — «бабочка»), которое является производным от *mesti, *motati sę («кружиться, двигаться рывками») из-за особенностей полёта насекомого[7][8].
Для обозначения мелких неярко окрашенных чешуекрылых используют слово «моль», происходящее от праслав. *molь, которое, по одной версии, является производным от *melti («молоть», и тогда этимологическое значение слова — «дробящее (повреждающее) насекомое»), а по другой — связано с др.-греч. μῆλον («мелкий скот»), ирл. míl («животное» — с этимологическим значением «мелкое животное»)[9].
Общая численность


Отряд чешуекрылых по видовому разнообразию несомненно выделяется среди таксонов подобного ранга. Чешуекрылые являются одной из крупнейших групп насекомых, включающей в себя, по состоянию на август 2013 года, 158 570 видов, в том числе 147 ископаемых таксонов[10]. Предполагается, что до 100 тысяч видов остаются всё ещё не известными науке и, таким образом, общее число существующих на планете видов чешуекрылых может быть оценено приблизительно в 200[11][12] — 225 тысяч видов[13]. На территории России встречается 2251 род и 9617 видов из 97 семейств[14]. Из них 7 семейств встречаются только на Дальнем Востоке (где всего обнаружено 1609 родов и 4871 вид бабочек)[15].
Чешуекрылые характеризуются большим разнообразием, при этом большинство их видов плохо изучены. Некоторые виды известны по находкам только из одной местности либо даже по единственному экземпляру. Истинная оценка общего количества существующих видов никогда не будет известна, потому что многие виды вымерли ещё до их обнаружения. Систематика бабочек, представленная в различных работах, отражает различные взгляды их авторов и является, вне всякого сомнения, дискуссионной. Имеются споры, посвященные систематическому положению или необходимости сохранения статуса некоторых подвидов или видов. Исследования ДНК свидетельствуют, что некоторые из известных в настоящее время видов должны быть разделены. Хорошо известным примером является случай, когда на первый взгляд идентичные Colias alfacariensis и Colias hyale, ранее считавшиеся одним видом, были разделены на два, после того как были обнаружены существенные различия в строении их гусениц и куколок.
Размеры
Размеры представителей отряда колеблются в широких пределах. Южноамериканская тизания агриппина (Thysania agrippina), вероятно, является крупнейшей по размаху крыльев бабочкой в мире. Согласно литературным данным, размах её крыльев может достигать 28 см[2]. Самой большой по размаху крыльев дневной бабочкой является орнитоптера королевы Александры (Ornithoptera alexandrae) — размах крыльев самки достигает 27,3 см[2]. К крупнейшим дневным бабочкам также относится Papilio antimachus из Центральной Африки, размах крыльев самцов которых может достигать 25 см[11]. Самки сатурнии Coscinocera hercules, обитающей в Австралии и Новой Гвинее, обладают самой большой площадью крыльев[16] — до 263 см²[1]. Одними из самых больших бабочек в мире также являются павлиноглазки Attacus atlas (размах крыльев — до 24 см) и Attacus caesar (размах крыльев — до 25 см[17])[18].
Одними из самых мелких известных бабочек является так называемые моли-малютки семейства Nepticulidae: Stigmella (Trifurcula) ridiculosa[1][19], обитающая на Канарских островах и обладающая размахом крыльев 3,8—4,5 мм[20], а также распространённые в северной Европе Stigmella filipendulae с размахом крыльев от 3,25 до 5 мм и Enteucha acetosae с размахом крыльев около 4 мм. К самым мелким видам также относятся голубянки Micropsyche ariana из Афганистана с размахом крыльев около 7 мм; Brephidium exilis из США с размахом крыльев 6—10 мм.
Самая крупная дневная бабочка России — парусник Маака (Papilio maackii), с размахом крыльев до 13,5 см[11]. Крупнейшей ночной бабочкой Европы и России является павлиноглазка грушевая (Saturnia pyri), с размахом крыльев до 15 см[21][22].
Морфология
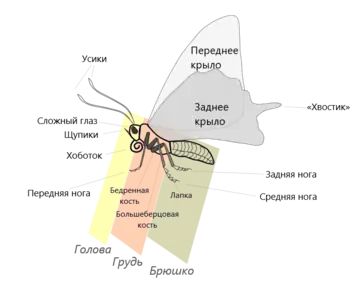
Строение
Как у всех насекомых, тело бабочек делится на три главных отдела: голову, грудь и брюшко. Снаружи оно защищено твёрдым хитиновым покровом, образующим наружный скелет.
Голова
Голова малоподвижная, свободная, округлой формы, с уплощённой затылочной поверхностью. Цервикальные склериты хорошо развиты, треугольные или Г-образные. Тенториум П- или (у ряда примитивных семейств, например, Agathiphagidae) Y-образной формы, у примитивных семейств (например, Micropterigidae, Eriocraniidae) также с хорошо выраженными дорсальными отростками.
Ротовые аппараты чешуекрылых разнообразны. У ряда примитивных семейств (например, Micropterigidae, Heterobathmiidae) — грызущий, с функционирующими мандибулами и развитыми жевательными лопастями максилл. У видов подотряда Glossata ротовой аппарат сосущего типа — хоботок — образован сильно видоизменёнными нижними челюстями, образующими трубочку, в покое спирально свёрнутую. Хоботок состоит из двух полутрубчатых частей, которые соприкасаются краями и скрепляются заходящими друг на друга щетинками. Хоботок служит для питания жидкой пищей. Внутри него расположены мелкие чувствительные волоски, выполняющие рецепторную функцию.
Имаго ряда видов имеют редуцированные ротовые органы (например, платяная моль, павлиноглазки и многие другие), не питаются вовсе и живут исключительно за счёт запасов питательных веществ, накопленных в стадии гусеницы[23]. Верхние челюсти у представителей отряда отсутствуют или представлены небольшими бугорками. Нижняя губа редуцирована, образует треугольный или сердцевидный валик у основания хоботка с нижней стороны. Её щупики хорошо развиты и состоят из трёх члеников.
У булавоусых бабочек и бражников на последнем членике щупиков располагается глубоко залегающая ямка, в нижней части которой находятся чувствительные рецепторы в виде конусов. Её отверстие располагается на свободном конце щупика и окружено целым рядом чешуек.
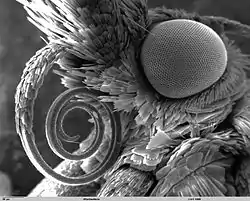
По бокам головы расположены полушаровидные, хорошо развитые, сложные, фасеточного типа глаза, состоящие из большого числа (до 27 000) омматидиев, дающих общее мозаичное изображение. Глаза обычно выпуклые, круглые или овальные, иногда почковидные. Они занимают значительную часть боковой области головы и окружены волосками, расходящимися венчиком. Порой тонкими короткими волосками покрыта вся глазная кутикула. Чешуекрылые способны различать цвета, но в какой мере — пока изучено недостаточно. Некоторые бабочки, например, крапивница (Aglais urticae) и капустница (Pieris brassicae), различают красный цвет, в то время как сатиры (Satyrinae) его не видят. В наибольшей степени привлекают бабочек два цвета — сине-фиолетовый и жёлто-красный[24]. Бабочки воспринимают также ультрафиолетовую часть спектра, чувствительны к поляризованному свету и способны ориентироваться по нему в пространстве. Движущиеся объекты различают гораздо лучше неподвижных[11][24].
Кроме фасеточных глаз, у чешуекрылых позади усиков на темени нередко расположены 2 теменных простых глазка[25]. Лобный глазок отсутствует.
Усики располагаются на границе между теменем и лбом и являются органами чувств, воспринимающими запахи и колебания воздуха. Усики также помогают удерживать равновесие при полёте. По строению различают щетинковидные, нитевидные, булавовидные, крючковатые, гребенчатые, перистые типы усиков[23]. У самцов, отыскивающих самок по феромонам (павлиноглазки, волнянки), усики сильно разветвлены и имеют гораздо большую поверхность, чем у самок[11].
Усики обладают неодинаковой длиной у представителей разных групп. У представителей семейства длинноусые моли, особенно у самцов, значительно превышают длину тела, а у тонкопрядов они являются сильно укороченными, немногим длиннее головы; у бескрылых самок мешочниц они почти полностью редуцированы. Количество члеников в усиках обычно достигает нескольких десятков. Первый членик, или скапус, крупнее остальных.
Грудь
Грудь, как и у всех насекомых, состоит из трёх сегментов: переднегруди, среднегруди и заднегруди — несущих три пары ног и две пары крыльев. Переднегрудь значительно меньше крылоносных сегментов (средне- и заднегруди), её тергит обычно несёт пару обособленных пластинок (патагии), наиболее развитых, главным образом, у представителей семейства совки.
Тегулы (подвижно сочленённые пластинки в основании крыльев) среднегруди мощно развиты, покрыты чешуйками, почти полностью прикрывают основания передних крыльев и хорошо заметны, в отличие от большинства отрядов насекомых[23].
Строение ног большинства бабочек типично для насекомых, они ходильные или бегательные. Тазики крупные, соединение их с грудью малоподвижное, свободная часть ноги включает вертлуг, бедро, голень и лапку (у большинства видов все лапки пятичлениковые с парой коготков на конце). К специфическим особенностям отряда следует отнести характерную формулу шпор (1—2—4) — подвижно сочленённых с голенями шипов. Посередине передних голеней, как правило, располагается одна шпора — так называемый эпифиз, используемый для чистки усиков. Средние голени несут на своей вершине одну пару шпор, а задние — две пары, которые располагаются посередине и на вершине голени. У некоторых групп бабочек часть шпор редуцирована. У представителей немногих групп ноги подвергаются более или менее значительной редукции: у самцов тонкопряда Hepiolopsis hecta задние лапки редуцированы, а задние голени лопатообразно расширены и несут пахучие чешуйки. Почти полностью редуцированы ноги самок некоторых мешочниц (Psychidae). У некоторых дневных бабочек в той или иной степени редуцированы передние ноги, у голубянок — менее значительно, у нимфалид — до полной утраты способности к хождению, в результате чего эти бабочки передвигаются на четырёх ногах (средних и задних). У большинства видов на лапках располагаются вкусовые рецепторы — видоизменённые сенсиллы[24].
Крылья
Крыльев две пары, перепончатые, с небольшим количеством поперечных жилок. Жилкование крыльев играет важную роль в систематике бабочек на различных уровнях — от вида до подотряда. Ранее многие энтомологи разделяли всех чешуекрылых на Homoneura (с приблизительно одинаковыми по форме и жилкованию передними и задними крыльями) и Heteroneura (с уменьшенным количеством радиальных жилок на заднем крыле и часто — значительно отличающимися по размерам и форме передними и задними крыльями). Эти названия и сейчас сохраняются для обозначения нетаксономических групп в пределах отряда.
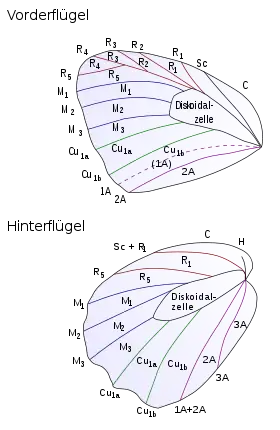 |
 |
У самцов крылья всегда вполне развиты; у самок представителей ряда семейств — листовёртки (Tortricidae), Oecophoridae, волнянки (Lymantriidae), медведицы (Arctiidae), пяденицы (Geometridae) — встречаются отдельные случаи частичной или полной редукции, а в семействе мешочницы (Psychidae) бескрылы самки большинства видов[23]. У Pterophoridae крылья расщеплены на линейные, перистые лопасти.
По участию крыльев и их мускулатуры в процессе полёта чешуекрылых относят к бимоторным и переднемоторным[26]. Бимоторность означает в одинаковой степени развитие обеих пар крыльев и приводящие их в движение мышц. Переднемоторность представляет собой объединение и содружественность в работе передних и задних крыльев. В ходе такого объединения возникает функциональная двукрылость — обе пары крыльев функционируют как одна. При этом во время полёта основная нагрузка приходится на переднюю пару крыльев, что порой приводит к ослаблению задней пары[26].
Сцепочный аппарат крыльев у низших групп образован выступом сзади у основания переднего крыла, который выступает поверх переднего края заднего крыла[23]. У высших групп бабочек этот же эффект достигается с помощью щетинки или пучка щетинок, располагающихся у переднего края заднего крыла[23]. Большинство видов булавоусых бабочек лишены такого «крепления», и скреплению крыльев способствует выступающий вперёд край задних крыльев, на который заходит край верхних крыльев[23][26].
У представителей инфраотряда разнокрылых бабочек большее значение в процессе полёта приобретает передняя пара крыльев, а задняя пара часто уменьшается в размерах и теряет часть жилок. Соответственно происходит усиление среднегрудного сегмента и его мускулатуры, тогда как заднегрудной сегмент ослабляется.

Крылья, как правило, полностью покрыты чешуйками, которые представляют собой видоизменённые щетинки[11][23][27]. Чешуйки имеют разнообразную форму, но чаще всего бывают плоскими. В их основании находится короткий стебелёк, которым чешуйка прикрепляется в углублении крыла, называемом мешочком чешуйки[28]. На конце стебелька имеется расширение, входящее во внутреннюю часть мешочка. Часто в стебельковой части чешуйки находятся выемки (синусы), имеющиеся исключительно у булавоусых бабочек, в то время как разноусых чешуйки цельнокраевыми[28]. Однако чешуйки на нижней стороне крыльев булавоусых бабочек также являются цельнокраевыми[28].
По краю крыла бабочек обычно располагаются узкие волосовидные чешуйки, нередко с разветвлёнными вершинами, образующие так называемую бахромку, в середине крыла — более широкие[11]. Тонкое строение чешуек играет важную роль в макросистематике чешуекрылых[28].
Чешуйки располагаются не только на крыльях, но и по телу бабочки[28]. Так, у булавоусых бабочек чешуйки на груди мелкие, неправильной формы, с заострённым краем. Грудные чешуйки прочих групп крупнее, одинаковой формы и размера. Чешуйки, покрывающие ноги, продольные и мелкие. Часто чешуйки имеют продольную ребристость, обусловленную выступающими хитиновыми трёхгранными канальцами[28].




Брюшко
Брюшко чешуекрылых обычно состоит из 9—10 сегментов, I из которых частично редуцирован, его стернум слит со стернумом II сегмента[19]. Остальные сегменты отличаются простотой и постоянством строения, образуя склеротизованные кольца, по бокам прерванные мембранозными участками — плейритами и снабжённые дыхальцами вплоть до VII сегмента включительно. Брюшко удлинённое, цилиндрической формы, причём у самцов оно тоньше и иногда несколько уплощённое с боков. Тергиты и стерниты примерно одинаковы по величине, дыхальца расположены на мембранозных плеврах. В состав брюшка, как и у остальных насекомых, входит десять сегментов, причём I и II сегменты обычно более или менее редуцированы, а X сегмент нередко бесследно сливается с IX. VIII сегмент у самок бывает удлинён, образуя часть яйцеклада, а у самцов — видоизменён в связи с функционированием копулятивного аппарата. Обычно IX—X сегменты самцов несут копулятивные придатки, а VIII—X сегменты самок преобразованы в той или иной степени развитый телескопический яйцеклад. Гениталии самца в спокойном состоянии втянуты внутрь VIII сегмента брюшка (исключение: Micropterigidae)[19] и выдвигаются наружу только перед копуляцией. Их скелетной основой является IX сегмент, являющийся у архаичных форм цельным склеротизованным кольцом — аннулус (annulus). В более развитых таксонах IX сегмент обычно разделён на дорсальную часть — тегумен (tegumen), и вентральную — винкулум (vinculum). Особенности их строения иногда используется для диагностики семейств, подсемейств и триб[19].
Ряд семейств (например, совки, пяденицы, многие огнёвки) обладают своеобразным органом слуха — так называемым тимпанальным аппаратом[19], который представляют собой затянутые мембраной углубления заднегруди, или первых двух сегментов брюшка, снабжённых механорецепторами. Распространяющиеся в воздухе звуковые колебания заставляют мембрану вибрировать, что вызывает возбуждение соответствующих нервных центров[29][30]. Эти органы способны воспринимать ультразвуковые колебания от 10 до 100 кГц[30].
Внутренние органы
Внутреннее строение чешуекрылых в целом типично для насекомых, за исключением нескольких характерных черт. В пищеварительном тракте имаго пищевод расширен, образуя зоб, мышечный желудок редуцирован. Единственный фермент, выделяемый пищеварительной системой имаго некоторых чешуекрылых — инвертаза[28].
Мужские репродуктивные органы состоят из двух дольчатых семенников, которые заключены в один общий, ярко окрашенный мешочек, из двух семяпроводов, местами образующих расширения (семенные пузырьки) и соединённых в один общий семяизвергательный канал, который открывается в основании выворачивающегося эндофаллуса или везики, заключённой внутрь склеротизованного копулятивного органа.
Женские органы размножения состоят из двух яичников с 4—5 (у примитивных видов — 20-ю[31]) яйцевыми трубочками у каждого, парных яйцеводов, объединяющихся в общий яйцевод, открывающийся наружу яйцевыводным отверстием. У Ditrysia, составляющих большинство бабочек, имеется также отдельное копулятивное отверстие, ведущее в копулятивную сумку, проток которой соединяется с общим яйцеводом в месте впадения семяприёмника (сперматеки) и придаточных желез. У Exoporia (Hepialidae) копулятивное и яйцевыводные отверстия также обособлены, но соединены желобком на брюшной поверхности тела. Наконец, у монотризных чешуекрылых копулятивное отверстие соединено с яйцевыводным.
Строение копулятивного аппарата у каждого вида имеет свои характерные особенности, и в некоторых случаях близкие формы, которые тяжело отличаются по внешним признакам, могут быть распознаны именно по копулятивным органам самцов.
Пигменты и окраска
Внешний вид и окраска крыльев бабочек имеют важную роль в повышении их выживаемости. Это и защитный камуфляж, внутривидовое половое распознание при спаривании, и апосематическая демонстрация. Окраска крыльев чешуекрылых может быть пигментной (зависит от содержащихся пигментов), оптической (зависит от преломления света) и комбинационной (сочетающая два предыдущих типа окраски). Яркая окраска крыльев некоторых бабочек (например, таких крупных и эффектно окрашенных бабочек, как Ornithoptera) контролируется двумя основными механизмами: диффузным отражением падающего света от мультислоёв и спектральной фильтрацией внутреннего пигмента[32].
 |
.jpg.webp) |
 |
| Лимонница (Gonepteryx rhamni). Пример пигментной окраски крыльев. | Сатир Cithaerias esmeralda. Прозрачные крылья почти полностью лишены чешуек. | Morpho rhetenor. Пример оптической окраски крыльев. |
Каждая пигментная чешуйка содержит в себе только один пигмент. Наиболее часто это меланин, придающий чёрный и коричневый цвета, и липохромы каротиноиды, характеризующиеся светочувствительностью и обуславливающие жёлто-зелёную, жёлтую, бурую, оранжевую и красную окраску[11][28]. Флавоноиды обеспечивают окраску жёлтого, белого, красного и коричневого цветов[27]. У белянок жёлтая окраска обусловлена пигментом ксантоперином и пуриновыми основаниями, образующимися из накопленной мочевой кислоты[11].
Выделяют два типа пигментов — диффузные и зернистые. Первые окрашивают хитин, но у чешуек пигменты содержатся в рельефе, а хитиновая пластинка часто лишена пигмента либо слабо окрашена. Зернистые пигменты имеются только у представителей семейства белянки (Pieridae) и погружены в хитиновую оболочку чешуек[27][28].

Расположение различно окрашенных чешуек на крыле закономерно и образует сложные рисунки, характерные для каждого вида, а часто и систематической группы.
При оптической окраске в чешуйках крыльев возникает тонкослойная интерференция. Пигмент в нижней части этих чешуек не пропускает свет и придаёт бо́льшую яркость интерференционной окраске. Лучи света, проходя через чешуйки, отражаются как от их внешних, так и от внутренних поверхностей, и в результате два отражения накладываются и усиливают друг друга[11]. Среди обладающих металлической окраской чешуек бабочек можно различить два типа: с морщинистыми пластинками чешуек и косыми параллельными бороздками (морфиды); и с гораздо более толстой верхней пластинкой, которая является относительно ровной (урании).
Чешуекрылые с оптической окраской крыльев — морфиды, урании и отчасти парусники, некоторые виды нимфалид, голубянок, белянок и некоторые другие.
Своеобразны сатиры рода Cithaerias из Южной Америки, крылья которых, утратив чешуйки, стали прозрачными, и только на задних крыльях сохраняются типичные для сатиров «глазки». Частично прозрачные крылья также имеют некоторые виды геликонид, стеклянниц, бражников[27].
Почти у всех бабочек рисунок на левой паре крыльев идентичен рисунку правой, являясь его зеркальным отражением, но существует исключение — мадагаскарская урания (Chrysiridia madagascariensis) с асимметричностью рисунка на крыльях.
Полиморфизм и иные формы изменчивости окраски
В некоторые жаркие и засушливые годы обнаруживается связь между погодными условиями и изменениями внешнего вида бабочек: появление бабочек с признаками южных климатических форм. Смена времён года вызывает у многих видов явления сезонного диморфизма и триморфизма. Среди дневных бабочек можно также найти также несколько примеров полиморфизма — одновременного и совместного существования различно окрашенных форм в одном виде, свободно скрещивающихся между собою и передающих свои признаки потомству (например, желтушки, большая перламутровка и другие).
Полиморфизм
Одним из типов изменчивости являются случаи полиморфизма, когда среди особей одного и того же вида в одной и той же местности и в одно и то же время встречается несколько различных форм, резко отличающихся между собою внешностью, не связанных между собою переходами, хотя данные формы могут свободно скрещиваться между собою[33]. Полиморфизм — появление различных форм, отличающихся по окраске и ряду признаков, внутри одного вида. Например, у особей медведицы-кайя (Arctia caja), развивающихся в одинаковых условиях, порой наблюдается вариабельность окраски, причём иногда эти различия настолько велики, что бабочек можно принять за представителей различных видов[28].
Среди европейских видов, примером полиморфизма служит большая лесная перламутровка (Argynnis paphia), у которой самцы одинаковый окраски, но самки бывают двух форм. Наряду с обычными рыжими самками, встречается тёмная форма valesina, у которой общий фон крыльев является зеленовато-серым[33]. Если отличия между различными формами самцов или самок ограничиваются цветом окраски, не задевая других признаков, то такие частные случаи полиморфизма сводятся к дихроизму (двухцветности) или полихроизму (многоцветности) особей каждого пола; так, например, у упомянутой выше большой лесной перламутровки имеет место дихроизм самок[33].
Сезонный полиморфизм
Различия внешних условий, действующих на куколку так или иначе отражаются в признаках самой бабочки. Под влиянием температурного воздействия на куколку у многих видов появляются изменения в окраске и рисунке крыльев, а иногда даже и в форме крыльев. Такая изменчивость выражается в виде так называемого сезонного диморфизма, когда различные поколения одного и того же вида, развивающиеся в различные месяцы года при разных погодных условиях, отличаются внешне какими-то признаками[33]. Это явление также называется сезонным полифенизмом.
Примером сезонной изменчивости является пестрокрыльница изменчивая (Araschnia levana). У весеннего поколения крылья рыжевато-красной окраски, с рисунком из чёрных пятен, с белыми пятнышками у вершины переднего крыла; у летнего поколения окраска крыльев чёрная с белыми пятнами. Изменение окраски в разные сезоны связано не только с температурным режимом, но и с состоянием куколки. Количество пигмента в крыльях бабочки зависит от содержания гормона 20-гидроксиэкдизона в куколке — весной его вырабатывается меньше, а летом больше[28][34]. Многие тропические бабочки также имеют сезонные формы, которые называются формами сухого и влажного сезонов.
Сезонный полиморфизм также может наблюдаться в менее выраженной форме у ряда других видов: Leptosla sinapis, Chrysophanus phlaeas, Chrysophanus dorilis, Everes argiades, Cyaniris argiolus и многих других[33].
Явления сезонного полиморфизма не всегда являются связными с действием низких температур на куколку весеннего поколения. У многих тропических видов чешуекрылых имеются формы дождливого сезона и формы сухого сезона.
| Сезонный полиморфизм | Географический полиморфизм | Половой диморфизм |
|---|---|---|
 |
_male_(8361988622).jpg.webp) |
.jpg.webp) |
 |
 |
 |
| Пестрокрыльница изменчивая (Araschnia levana). Вверху весеннее поколение f. levana Araschnia levana f. levana. Внизу летнее поколение 'f. prorsa |
Самец Ornithoptera priamus urvillianus (верхнее фото). Самец Ornithoptera priamus priamus (нижнее фото) |
Ornithoptera goliath. Самец (наверху) и самка |
Температурные формы и аберрации


У некоторых видов умеренных широт также имеются сезонные различия во внешности бабочек различных поколений, хотя ни одно из них не подвергалось воздействию низких температур во время стадии куколки. Таковыми являются желтушка луговая, голубянка икар, буроглазка большая, перламутровка селена и некоторые другие[33]. В подобных случаях сезонные различия между бабочками различных поколений могут зависеть от разницы в условиях жизни гусениц, частью и от неодинаковой температуры окружающей среды, действию которой подвергаются куколки в различные месяцы летнего сезона.
Температурные воздействия на куколку вызывают изменения внешности у ряда видов, например крапивницы, траурницы, многоцветницы, многоцветницы чёрно-рыжей и других, называемые «температурными формами»[33].
При воздействии повышенной температуры, вышедшая из куколки бабочка будет иметь окраску, характерную для южных форм, а при пониженной температуре возникают формы с северным типом окраски[35]. Так, у траурницы, при воздействии низких температур на её куколку, темнеет общий фон, и уменьшаются голубые пятна на крыльях. Красный цвет на крыльях может заменяться чёрным (медведица Гера), жёлтый — на красный (медведица-госпожа)[28].
Аберрациями называют непостоянные, случайно возникающие изменения внешнего вида бабочки, появляющиеся под воздействием различных, преимущественно температурных, факторов на куколку. Чаще всего аберрации проявляются появлением нетипичной окраски крыльев. К аберрациям также принято относить появление особей с разным размером одноимённых крыльев, когда одно крыло является больше другого.
Географический полиморфизм

Разные популяции в различных частях своего ареала отличаются друг от друга определёнными признаками. Примером может служить окраска различных подвидов орнитоптеры приам (Ornithoptera priamus), обитающих на Молуккских и Соломоновых островах, на Новой Гвинее. Так самцы подвида poseidon имеют ярко-зелёную окраску крыльев, подвида demophaneus жёлто-зелёную окраску, подвида urvillianus — ярко-синюю окраску, а подвида miokensis обладают крыльями от синего до цвета морской волны[27].
Ещё более известным примером географического полиморфизма являются представители рода аполлонов (Parnassius), в том числе обыкновенный аполлон — расположение и размер пятен на крыльях которого сильно варьирует, образуя более 600 известных форм[36]. Аполлоны образуют локальные популяции, не имеющие между собой контактов, а из-за сильной стенотопности видов и слабой миграционной способности скрещивание между популяциями одного вида практически не происходит. Почти в каждом изолированном ареале рисунок на крыльях одного и того же вида аполлонов немного отличается[36].
Альбинизм, меланизм, нигризм
Окраска иногда может изменяться и вследствие отсутствия пигментов — альбинизм, или же наоборот изменяться от более яркого пигмента, либо избытка тёмного — меланизм[27][37]. Появление альбинизма и меланизма гораздо чаще встречаются у представителей разноусых чешуекрылых.
Крылья и другие части тела могут быть равномерно побледневшими. Иногда бледнеют не все чешуйки, а среди них остаются небольшие группы с нормальной окраской, равномерно разбросанные по поверхности крыла. Такие экземпляры иногда встречаются среди разных видов Satyridae, Epinephete jurtitta и Epinephete tycoon[33]. У других особей побелевшие участки могут быть распределены по крылу несимметрично. Встречается также и такая форма альбинизма, когда побелевшими бывают лишь те участки крыла, которые у нормальных особей окрашены в один определённый цвет (например, рыжий фон крыльев у некоторых перламутровок), тогда как все другие части рисунка сохраняют свою обычную окраску[33].
Альбинизм у чешуекрылых неизменно бывает связан с недоразвитием самих чешуек, которые приобретают уродливую форму[33].
Избыточное окрашивание крыльев и тела чёрными пигментами наблюдается гораздо реже альбинизма. При меланизме обычный чёрный рисунок бабочки остаётся нормальным, но вместе с тем общий фон крыльев бывает более или менее потемневшим, иногда настолько, что вся бабочка кажется равномерно чёрной (различные виды белянок, Synchloe uaptidice)[33].
От настоящего меланизма следует отличать явления нигризма, когда чёрный рисунок на крыльях бывает изменён — чёрные полосы и пятна увеличиваются относительно своих размеров, расплываются или вытягиваются и сливаются между собой. Сюда принадлежат такие примеры, как самки капустниц, у которых два чёрных пятна соединяются перемычкой (ab. nigronotata), экземпляры голубянок, с круглыми пятнами на нижней стороне крыльев в форме восклицательного знака или слившиеся в продольные полоски и т. п.[33] Более резкие и эффектные примеры нигризма можно встретить среди перламутровок (Argynnis) и шашечниц. В крайних случаях, обычный рисунок бабочки бывает изменён настолько, что от рыжего фона крыльев остаётся только несколько пятен вдоль края крыла.
Изменения в сторону нигризма возникают обычно под влиянием внешних условий — преимущественно температурных[33], поэтому данные изменения необходимо относить к числу аберраций.
Половой диморфизм

Половой диморфизм — различия во внешнем виде, окраске и/или форме тела самца и самки — широко распространён и порой резко выражен среди различных чешуекрылых. Кроме отличий в строении усиков, самки многих чешуекрылых, как правило, крупнее самцов, с крупным брюшком; обладают менее яркой окраской, иногда полностью отличающейся от таковой у самцов. В большинстве случаев, такие отличия вызваны репродуктивной яйцекладущей функцией и меньшей подвижностью самок. Наиболее сильное проявление полового диморфизма — частичная или полная утрата крыльев самками (мешочницы, некоторые пяденицы и волнянки), а также ног и усиков. Такая бескрылость самок связана с климатическими условиями и типична для высокогорных и арктических видов, а также летающих ранней весной или поздней осенью. Например, самка волнянки Lachana selenophora из Памира и Тянь-Шаня с полностью редуцированными крыльями почти лишена ног и обладает большим бочковидным брюшком для продукции яиц[38].
У бабочек из рода орнитоптеры (Ornithoptera) самцы меньше самок и имеют яркую окраску крыльев, самки же гораздо крупнее и окрашены в коричнево-чёрные тона. Самцы многих морфид (Morpho) обладают яркой синей окраской, в то время как самки — жёлто-коричневые.
Одним из проявлений полового диморфизма являются различия в окраске крыльев, видимые только в ультрафиолетовом излучении[28].

Индустриальный меланизм
Индустриальный меланизм — повышение частоты тёмноокрашенных форм, связанное с интенсивным развитием промышленности[39][40]. Около районов с крупными промышленными центрами в Европе (особенно Англии) и в Северной Америке описаны десятки видов, в популяциях которых обнаружены меланистические формы.
Наиболее известный пример данного явления — берёзовая пяденица (Biston betularia). До середины XIX века все собранные энтомологами экземпляры берёзовой пяденицы имели бело-сероватую окраску крыльев с тёмными пятнышками (morpha typica), что обеспечивало покровительственную окраску на стволах деревьев. Сейчас многие популяции полиморфны, в них присутствуют чёрные меланистические формы — Biston betularia morpha carbonaria[39].
Увеличение частоты меланистических форм — следствие направленного отбора, главным движущим фактором которого является избирательное поедание особей бабочек птицами[41]. В лесах вокруг промышленных конгломератов и городов стволы деревьев часто лишены лишайников и могут быть почерневшими от копоти. В таких районах покровительственной окраской является чёрная, а в незагрязнённых районах — светлая пятнистая окраска[39].
Также известна третья форма, имеющая промежуточную по темноте окраску между меланистической и светлой формами берёзовой пяденицы — morpha insularia. Данный тип окраски, как и меланистическая, доминирует над светлой, однако, видимо, детерминируется не одним, а несколькими локусами.
На основании лабораторных опытов предполагалось, что аллели, контролирующие меланистическую окраску, обладают плейотропным действием, определяя не только окраску бабочки, но и её поведение (выбор фона)[40]. Однако это не было подтверждено наблюдениями в природе.
Физиология
Полёт


Основной формой передвижения чешуекрылых является полёт, как активный (машущий), так и пассивный (парящий, планирование, пикирование)[26].
Действия крыльев бабочки при активном машущем полёте представляют собой последовательность определённых движений, образующих замкнутый цикл. Каждый цикл состоит из взмаха крыльями, находившимися в крайне верхнем положении, вниз, не ниже корпуса, и возвратного взмаха крыльями вверх. При взмахе крыльями вниз бабочка приобретает некоторый импульс движения вверх, отталкиваясь от воздуха.
Способностью к парящему полёту обладают представители семейств: данаиды, Nymphalidae, Papilionidae, Pieridae, Morphinae, Uraniidae. Мелкие виды из семейства нимфалиды для парения используют термические потоки, образующиеся у самой земли, более крупные виды парят гораздо выше, часто на уровне крон деревьев[26]. Ещё более приспособленными к парению являются представители семейства парусников, особенно парнассиусы, которые могут длительное время парить над горными склонами, корректируя траекторию полёта взмахами крыльев и меняя высоту[11][26].
Повышение устойчивости при парящем полёте достигается благодаря длинным хвостовидным выростам задних крыльев.
Особенно большой длины подобные выросты достигают у бабочек семейства сатурнии — например, Actias artemis, Argema mittrei, Coscinocera hercules, Argema mimosae[26].
Все виды, способные к парению, также используют и планирование, к которому часто прибегают нимфалиды, парусники, данаиды и другие чешуекрылые[26]. Характерным признаком приспособления к последнему служит вытянутость вершин передних крыльев. Планирующий полет характерен также для чешуекрылых с большой площадью крыльев. Разогнавшись, они прекращают движения крыльями, и некоторое время планируют благодаря инерции[26].
Пикирование как разновидность пассивного полёта отмечается у обитающих в траве медведиц Callimorpha и совок из группы Leucanilis. Крылья бабочки при таком пикировании немного отведены назад, что помогает достичь высокой скорости[26].
Полёт представителей группы Heterocera заметно отличается от полёта булавоусых бабочек, в первую очередь это связано с углом дуги, описываемой крылом — у булавоусых бабочек он достигает 140—150°, а у разноусых бабочек — 80—90°[11].
Для полёта большинства видов оптимальной температурой тела являются показатели в пределах 30—35 °C[16]. Ночные виды перед полётом усиленно вибрируют крыльями, повышая таким образом температуру тела[16].
Наиболее совершенен полёт у представителей семейства бражники — обладая быстрым маневренным полётом, они способны зависать в воздухе, летать вертикально вверх и вниз, а также назад, подобно колибри. Большинство бабочек могут развивать скорость 8—17 км/ч[11].
Питание

Большинство видов, будучи антофилами, питается цветочным нектаром. Многие чешуекрылые также питаются соком деревьев, гниющими и перезревшими фруктами. Бражник мёртвая голова охотно поедает мёд из гнёзд и ульев пчёл. Первичные зубатые моли питаются пыльцой.
Виды с редуцированными ротовыми органами не питаются и живут исключительно за счёт запасов питательных веществ, накопленных в стадии гусеницы.
Ряду видов из семейств нимфалиды, парусники и других, для жизнедеятельности необходимы микроэлементы, в первую очередь натрий[11]. Они охотно летят на влажную минеральную глину, на экскременты и мочу крупных животных, влажный древесный уголь, человеческий пот — откуда получают влагу и необходимые микроэлементы[42]. Часто самцы этих видов собираются группами на влажном песке и глинистых почвах, по берегам ручьёв, возле луж[11][16][27].
Присутствуют среди бабочек, как исключение, и хищники[43], каковыми являются совки рода Calyptra: например, Calyptra eustrigata, обитающая на территории от Индии до Малайзии, Calyptra thalictri и Calyptra lata. Самцы питаются слёзной жидкостью (лакрифагия) и кровью крупных животных, прокалывая их покровы острым хоботком[43]. Самки же питаются соком плодов и растений[27]. У совки Hemiceratoides hieroglyphica, пьющей птичьи слёзы, развит специализированный хоботок с гарпуновидным кончиком[44].
Терморегуляция

Чешуекрылые, как и все насекомые, традиционно рассматриваются как пойкилотермные организмы (температура тела которых зависит от температуры окружающей среды), в отличие от теплокровных организмов (которые поддерживают стабильную внутреннюю температура тела, независимо от внешних воздействий). Однако термин «терморегуляциия» в настоящее время используется для описания способности насекомых поддерживать стабильную температуру (выше или ниже температуры окружающей среды) по меньшей мере в какой-либо части их тела, физиологическими или поведенческими средствами[45].
Многие чешуекрылые являются эктотермами (животные, для которых источником тепла является окружающая среда), другие же могут быть эндотермами (самостоятельно производить тепло внутри своего тела). Эндотермических насекомых корректней описать как гетеротермов, потому что они не являются целиком эндотермичными. Например, активные ночью виды бабочек производят необходимое для своего полёта тепло путём сокращения мышц груди, но их брюшко при этом практически не нагревается[46].
Определение пола
Вероятно, исходным для бабочек механизмом определения пола был механизм Z0 самка/ZZ самец[47]. Затем, путём хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У видов с системой Z0/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом, но у тутового шелкопряда (система WZ/ZZ) обнаружен отвечающий за развитие женского пола ген Fem в W-хромосоме.
Гинандроморфизм

Во время развития бабочки возможны генетические изменения, приводящие к появлению гинандроморфных бабочек — полусамцов-полусамок, у которых одна сторона тела полностью мужская, а другая — женская (так называемые билатеральные гинандроморфы).
Иногда аномальными становятся лишь небольшие участки тела бабочки, тогда говорят о мозаичном гинандроморфизме. Так, в различных участках крыла у самца могут появиться участки окраски самки и наоборот. Как правило, такие особи нежизнеспособны, но при сохранении функциональности внутренних органов имаго являются жизнеспособными.
У чешуекрылых основой для появления гинандроморфа обычно является образование двуядерных ооцитов, у которых одно из ядер образуется, например, из ядра полярного тельца. Если одно из этих ядер содержит Z-хромосому, а другое — W-хромосому, то при оплодотворении двумя спермиями при делении этих ядер могут возникнуть клетки-потомки мужской и женской половин тела. Второй возможной причиной является утрата ядром зародыша одной из Z-хромосом на первых стадиях дробления (обычно при первом делении зиготы)[27][37]. При развитии зародыша из потомков данного ядра произойдёт развитие клеток женской половины тела, этот вариант описан у пядениц рода Abraxas[48].
Частота появления гинандроморфов составляет примерно один случай на 90 000[27]. За свою редкость они высоко ценятся коллекционерами.
Интерсексуальность
Обычно развитие клеток насекомых генетически автономно, и половые гормоны не влияют на вторичные половые признаки. Поэтому для бабочек характерна генетически обусловленная интерсексуальность, возникающая в результате отклонения от нормы соотношения числа половых хромосом и аутосом (триплоидная интерсексуальность) или нарушения баланса генов (диплоидная интерсексуальность)[47]. Диплоидная интерсексуальность была описана Р. Гольдшмидтом у непарного шелкопряда (Lymantria dispar) при скрещивании разных географических рас, хотя в дальнейшем его данные ставились под сомнение[49].
Обычно бактерия Wolbachia вызывает у бабочек цитоплазматическую несовместимость или смену пола (превращение генетических самцов в функциональных самок), в определённых условиях у белянки Eurema hecabe и огнёвки Ostrinia scapulalis её воздействие может приводить к появлению истинных интерсексов[50][51].
У Morpho aega известны случаи появления форм самок с синей окраской крыльев, характерной для самцов данного вида. Синяя окраска у таких самок обусловлена тем, что форма чешуек на их крыльях такая же, как у самцов; возможно, эти самки являются интерсексами[27].
Межвидовое скрещивание

Межвидовое скрещивание — скрещивание особей различных видов. У чешуекрылых известны случаи межвидового скрещивания у близкородственных видов парусников, павлиноглазок, нимфалид. Гибриды чешуекрылых могут быть внутривидовыми (при скрещивании различных подвидов), внутриродовыми (при скрещивании видов принадлежащих одному роду). Как правило, межвидовые гибриды не способны к размножению[28]. Ряд гибридов бабочек специально искусственно разводятся путём культивирования человеком с коммерческой целью в виду их большой популярности и высокой стоимости среди коллекционеров насекомых, например гибриды Graellsia isabellae x Actias dubernardi, Graellsia isabellae x Actias luna, а также всевозможные межвидовые гибриды орнитоптер. Возникновение многих таких гибридов в природных условиях невозможно в виду географической или сезонной изолированности видов.
Генетика окраски
В последние годы большое внимание генетиков привлекает изучение генов, отвечающих за крыловой рисунок чешуекрылых[52].
Размножение

В биологии размножения бабочек важную роль играют расцветка и форма крыльев (привлечение партнёров), а также сложные формы ухаживания (брачные полёты и танцы). Некоторые виды характеризуются половым диморфизмом, который играет важную роль в нормальном течении полового цикла, так как внешние различия облегчают опознание и нахождение брачного партнёра своего вида[53].
Отыскиванию партнёрами друг друга способствуют и феромоны, вырабатываемые самцами булавоусых бабочек и самками ночных видов. Многие самцы булавоусых бабочек имеют пахучие чешуйки — андроконии, которые расположены на крыльях, реже — на лапках[11]. У белянок андроконии расположены почти по всей поверхности крыльев[33]. У других видов они сосредоточены на определённых участках тела, и часто образовывают на крыльях особые бархатистые пятна или полоски[33].
Органы, выделяющие феромоны, у самок находятся на заднем конце брюшка между 8 и 9 сегментами, и у ряда видов могут выдвигаться наружу. Самцы сатурний могут отыскать самку по феромонам на расстоянии нескольких километров[11][54].
У чешуекрылых могут отмечаться сложные формы ухаживания — брачные полёты и танцы. Некоторые, например, хвостатки устраивают брачные полёты — самцы летят под самкой, а затем разворачиваются в другую сторону, касаясь крыльями усиков самки; у лимонницы — самка летит впереди, а самец следует за ней, сохраняя постоянную дистанцию[16]. У других видов самка может сидеть на растении, а самец совершает «танцующий» полёт прямо над ней, выделяя феромоны, стимулирующие её к спариванию[16].
Спаривание происходит на земле или растении и длится от 20 минут до нескольких часов; всё это время спаривающиеся особи сохраняют неподвижность. Во время спаривания самка получает от самца не только сперматозоиды, но и ряд некоторых белков и микроэлементов, необходимых для формирования и откладывания яиц[16].
Самки парусников после спаривания несут на себе жёсткий хитиновый придаток на нижней стороне брюшка — сфрагис (лат. sphragis — печать, пломба), образуемый самцом в ходе спаривания. Чаще всего сфрагисы встречаются у парнассиусов, а также родственных видов. Предназначение сфрагиса — исключение повторного оплодотворения самки другими самцами.
Партеногенез встречается у бабочек крайне редко, наблюдается у мешочниц (Psychidae), молей рода Solenobia, в виде исключения — у некоторых шелкопрядов (тутовый шелкопряд)[28].
Геликонида харитония (Heliconius charithonia) отличается особенностями процесса размножения. За несколько часов до вылупления из куколки, самки выделяют феромоны, привлекающие самцов, которые соперничают за право спариваться с самкой. Обычно в таком соперничестве побеждает наиболее крупный самец, который с началом выхода самки из куколки сразу же начинает спариваться с ней. Данное явление в англоязычной литературе получило название «изнасилование куколки» (англ. pupal rape)[55].
Большинство бабочек в умеренном поясе Северной Америки и Европы производят одно или два-три поколения в год. В тропиках может быть до 8—10 и более поколений за тот же период времени. Например, жизненный цикл Hypolimnas misippus занимает всего 23 дня.
Жизненный цикл

Бабочки относятся к насекомым с полным превращением, или голометаморфозом. Их жизненный цикл включает четыре фазы:
Яйцо
.jpg.webp)
Яйца бабочек покрыты плотной твёрдой оболочкой и могут быть разнообразной формы. В зависимости от рода и вида яйца могут быть круглыми, цилиндрическими, шаровидными, яйцеобразными, угловатыми. Их внешняя поверхность может быть рельефной, образовывая углубления, точки, полоски, бугорки, расположенных, как правило, в симметричном порядке. Окраска чаще всего белая и зеленоватых оттенков, реже — бурая, жёлтая, красная, голубая, тёмно-зелёная, иногда с цветным рисунком[28].
Самки откладывают яйца на листья, стебли или ветки кормовых растений. Их число в кладке зависит от вида и может быть больше 1000[25][28], однако до взрослой стадии выживают немногие. В зависимости от вида, яйца могут откладываться поодиночке или группами по 10 и более[25]. Самки могут покрывать яйца волосками из густого пучка на конце брюшка. Отложенные яйца могут также покрываться выделениями придаточных половых желёз, которые, застывая, образуют защитный щиток (яблонная моль).
Самки эпикопеи (Epicopeia mencia) отличаются удивительной привязанностью к однажды выбранным для откладывания яиц местам, которые остаются неизменными на протяжении многих лет и, как правило, представлены одними и теми же деревьями ильма. Такая избирательность приводит к тому, что основная масса мест откладывания яиц концентрируется на немногих, пользующихся особой привлекательностью для бабочек, кормовых деревьях[56].
Средняя продолжительность стадии яйца 8—15 дней[28]. У многих видов умеренного пояса отложенные осенью яйца зимой находятся в состоянии диапаузы[28].
Гусеница
Гусеница является личинкой бабочки. Она обычно червеобразная, с грызущим ротовым аппаратом. Физиологической особенностью гусениц является наличие пары изменённых слюнных желез, открывающихся общим каналом на нижней губе и вырабатывающих особый секрет, который быстро затвердевает на воздухе, образуя шёлковую нить.
Большинство гусениц являются фитофагами — питаются листьями, цветками и плодами различных растений[4][57]. Всех растительноядных гусениц можно разделить на три категории: полифаги, олигофаги и монофаги Гусеницы некоторых видов — кератофаги — питаются воском, шерстью, роговыми веществами (гусеницы молей рода Ceratophaga живут в рогах африканских антилоп, питаясь кератином). Немногие виды (стеклянницы и древоточцы) являются ксилофагами и питаются древесиной, прогрызают ходы в стволах и ветвях древесных растений или корнях травянистых растений[58][59]. Гусеницы некоторых видов — хищники, питаются тлями, червецами, личинками и куколками муравьёв[4]. К примеру, гусеницы пядениц рода Eupithecia, которые обитают на Гавайских островах, являются насекомоядными и обладают целым арсеналом приспособлений для поимки добычи.
.jpg.webp)

Гусеницам некоторых видов свойственна олигофагия — питание очень ограниченным количеством видов растений. Например, гусеницы поликсены питается лишь четырьмя видами растений рода кирказон.
Гусеницами выделяются три главные группы пищеварительных ферментов — протеазы, карбогидразы и липазы[4].
Тело гусеницы состоит из 3 грудных и 10 брюшных сегментов. Кожа гладкая, с редкими бородавками, щетинками или волосками, шипами. Грудные сегменты несут 3 пары членистых ног, брюшные обычно имеют 5 пар более толстых ног, с крючками на подошвах, помогающие гусенице удерживаться на субстрате. У пядениц и некоторых совок имеет место уменьшение числа брюшных ног до 2—3 пар, или же они редуцируются[4].
Большинство гусениц ведут наземный образ жизни, но гусеницы ряда видов семейства ширококрылые огнёвки (Pyraustidae) обитают под водой, а у подводной огнёвки Acentria ephemerella под водой живут и взрослые бескрылые самки.
По образу жизни гусеницы условно разделяются на две большие группы[4]:
- гусеницы, ведущие свободный образ жизни, открыто питающиеся на кормовых растениях;
- гусеницы, ведущие скрытый образ жизни.
Гусеницы, ведущие скрытый образ жизни, обитают в переносных чехликах, которые они сооружают из шелковистой нити (мешочницы, Psychidae), либо из кусочков листьев (ряд огнёвок)[23][60]. Гусеницы таскают такой чехлик на себе, прячась в него при опасности. Другие гусеницы строят убежища из листьев, свёртывая и скрепляя их шелковистой нитью, часто образуя сигарообразную трубку. Скрытый образ жизни ведут и гусеницы, обитающие внутри различных частей растений. Сюда можно отнести бабочек из групп: моли-минёры (Tischeriidae), плодожорки (Laspeyresiini), а также виды-галлообразователи.
Окраска тела гусениц взаимосвязана с их образом жизни. Гусеницы, ведущие открытый образ жизни, имеют покровительственную окраску, сочетающуюся с определённой формой тела, порой напоминающей части растений[4]. Наряду с покровительственной окраской, распространена яркая демонстрационная окраска, свидетельствующая об их несъедобности. Некоторые гусеницы в момент опасности принимают угрожающую позу, например, большой гарпии, винного среднего бражника[61].
Гусеницы бабочек, живущих в северных широтах, порой не успевают завершить свой жизненный цикл за одно лето, из-за чего вынуждены впадать в зимнюю диапаузу до следующего лета. Обитающая за полярным кругом в Гренландии и Канаде волнянка Gynaephora groenlandica известна тем, что жизненный цикл её гусеницы может длиться до 14 лет[62]. Обычно продолжительность жизненного цикла составляет до 7 лет[62]. Вторая уникальная возможность гусениц данного вида заключается в том, что во время зимовок в стадии диапаузы они могут переживать температуру −60…−70 °C[62]. При снижении температуры окружающей среды в конце арктического лета гусеницы начинают синтезировать защищающие от холода химические соединения, такие как глицерин и бетаин[62]. Гусеницы проводят около 90 % своей жизни в замёрзшем виде в состоянии диапаузы и лишь около 5 % питаясь скудной растительностью. Жизненный цикл гусениц родственного ей вида волнянка Кузнецова, эндемика острова Врангель, длится до 6 лет[63].
Куколка
Куколка бабочек малоподвижная; у примитивных чешуекрылых, образующих таксоны Protolepidoptera и Palaeolepidoptera, куколки относятся к типу зубастых (pupa dectica); у подавляющего большинства чешуекрылых, образующих таксон Neolepidoptera, куколки относятся к типу склеенных (pupa conglutinata)[64]. У таких семейств, как сатурнии и коконопряды, куколка находится внутри специального сплетённого гусеницей кокона.
Форма куколки от вытянутой цилиндрической, яйцевидной до почти округлой. Окраска преимущественно покровительственная, однотонная от светлой до тёмной или с различным рисунком из точек, пятен и полос. Сразу после окукливания цвет часто бледный, иногда почти белый или зеленоватый, потом он меняется на свойственный данному виду, и куколка превращается, например, из зелёной в коричневую[65].
У неё хорошо различимы очертания зачатков крыльев, хоботка и ног, брюшко с дыхальцами по бокам, кремастер на конце брюшка — специальное утолщение, часто раздвоенное на вершине, с крючочками для крепления паутинными нитями к субстрату[65].
Иногда располагается открыто на растении, но чаще находится внутри оплетённых свёрнутых и склеенных листьев, либо в поверхностном слое почвы, на стеблях и стволах растений.
Куколка чаще прикреплена к субстрату (различают висячих куколок и подпоясанных)[65], реже лежит свободно на подстилке или земле среди растительных остатков в редкой паутине. Висячая куколка прикрепляется головным концом вниз при помощи кремастера, подпоясанная — головой вверх, кроме кремастера её поддерживает в таком положении специальный паутинистый поясок[65].
Куколки не питаются. У некоторых видов куколка является стадией, впадающей в зимнюю диапаузу.
Имаго
Из куколки выходит взрослое насекомое — имаго. Примерно за сутки до выхода бабочки, оболочка куколки становится маслянисто-прозрачной. Затем происходит разрыв куколки вдоль головы и переднего края крыльев, а имаго, цепляясь ногами за край разорванной оболочки, выползает наружу. Во многих группах чешуекрылых самцы выходят из куколок раньше самок, поэтому в начале периода лёта бабочек встречаются только самцы, а конце — только самки[16].
В первые минуты после выхода из куколки бабочка ещё не способна летать. Она взбирается на вертикальные возвышения, где остаётся до расправления крыльев, происходящего под действием гидростатического давления гемолимфы[66]. Расправленные крылья отвердевают и приобретают окончательную окраску.
У многих групп и видов имеется разница в сроках появления самцов и самок, которая составляет до 10 дней. Подобная задержка начала лёта самок обусловлена более высокой скоростью преимагинального развития самцов[67].
Имаго является половозрелой формой, способной к размножению. Основная функция данной стадии жизненного цикла — размножение и расселение. Продолжительность жизни имаго колеблется от нескольких часов до нескольких месяцев (у видов впадающих в диапаузу) и в среднем составляет 2—3 недели[28].
Распространение и места обитания

Чешуекрылые широко распространены по всему земному шару[53]. Их распространение совпадает с распространением наземной флоры, преимущественно цветковых растений. Только крайние полярные области, отдельные пустыни и океанические острова, а также высокогорья с вечными снегами не заселены бабочками. Наиболее многочисленны и разнообразны бабочки в тропиках. В умеренных широтах их видовой состав не так велик. Для общей характеристики расположения ареалов животных, в том числе и чешуекрылых обычно сушу Земли принято разделять на семь биогеографических царств или регионов. На территории шести из них распространены чешуекрылые[53][68].
- Палеарктический регион. На территории Палеарктики обитает более 22 тысяч видов чешуекрылых, из которых 1600 — булавоусые. На территории одной только Европы известно примерно 4500 видов, из которых более 400 видов — булавоусые[11].
- Неарктика. Фауна региона близка по видовому составу к Палеарктическому региону[53]. В Неарктической области встречаются около 14 тысяч видов бабочек, из которых 700 видов — дневные[11]. Виды чешуекрылых, которые обитают и в палеарктической и неарктической областях имеют голарктическое распространение.
- Неотропический регион. В регионе обитает 45 тысяч видов бабочек[11]. Большое количество видов из труднодоступных регионов Южной Америки, вероятно, ещё не описано.
- Афротропический регион. На территории региона обитает примерно 2,5—3 тысячи видов дневных чешуекрылых[11][53], среди которых три четверти обитающих на острове Мадагаскар являются эндемиками[11][53].
- Восточный (Индо-Малайский) регион. В регионе обитает около 4 тысяч видов дневных чешуекрылых[11]. Только на Филиппинах известно около 800 видов дневных бабочек[11][53].
- Австралийский регион. Многие виды являются общими для Индо-Малайского и Австралийского регионов, но имеются и эндемичные[53]. В Новой Гвинее описано около 800 видов дневных чешуекрылых, в Австралии — около 400 видов[11].
Среди районов, особо выделяющихся видовым разнообразием чешуекрылых, следует отметить Перу и индийский штат Сикким. В Сиккиме, площадью 7300 км2, обитает примерно 690 видов дневных чешуекрылых, а в Непале, площадь которого почти в 20 раз больше, — лишь 623 вида[11]. Местом, где встречается наибольшее количество видов чешуекрылых, считаются окрестности посёлка Тинго-Мария в Перу. Вообще, Перу является страной с наибольшим разнообразием чешуекрылых — здесь их встречается 3450 видов, тогда как в Бразилии, которая по площади больше Перу в 7 раз, обитает только 3130 видов[11].
Важнейшим фактором, предопределяющим распространение бабочек, являются их пищевые связи. У монофагов и узких олигофагов их ареалы часто сопряжены с распространением кормовых растений гусениц. Некоторые виды бабочек являются широко распространёнными. Например, репейница обитает в тропических и умеренных областях всех континентов, за исключением Южной Америки. Некоторые роды также обладают обширным ареалами — иногда почти всесветными. Наряду с этим, многие таксоны обладают весьма узкими ареалами либо являются эндемиками, к числу которых относятся многочисленные горные и островные виды[68].
В горах чешуекрылые встречаются до границы вечных снегов и льдов, однако их распространение в высокогорных районах очень плохо изучено. Недалеко от полярного круга на северо-востоке Якутии на высоте около 1500 м над уровнем моря встречается аполлон арктический (Parnassius arcticus)[11]. В Непале парусник «Слава Бутана» обитает в горах на высоте до 2800 м над ур. моря[11]. В Афганистане и Таджикистане желтушка Марко Поло (Colias marcopolo) обитает в горах на высотах от 3000 до 5000 м над ур. моря[69]. В Гималаях Parnassius charltonius обитает на высоте до 5800 м над уровнем моря[70]. Несколько видов заполярных волнянок — Gynaephora rossii и Gynaephora groenlandica — обитают даже на самой северной в мире суше — севере Канадского архипелага и севере Гренландии[71], а волнянка Кузнецова встречается только на острове Врангеля.
Время лёта и его продолжительность весьма разнообразны у различных групп чешуекрылых и ограничены тёплым временем года в умеренных широтах. Некоторые ранние весенние и поздние осенние виды совок и пядениц в средней полосе Евразии летают даже при минусовой температуре. Например, Leucobrephos middendorffi из Восточной Сибири летает исключительно при слабоминусовой температуре над снегом.
Экология
Территориальное поведение
Для некоторых бабочек, преимущественно нимфалид, морфо, белянок, голубянок, характерно территориальное поведение, как правило, отмечающееся у самцов. При этом они могут занимать в ожидании самок наивысшую точку ландшафта, либо же облетать свой участок обитания. Интересно, что со своих участков эти самцы прогоняют не только самцов своего вида, но и представителей других видов бабочек со схожей окраской, а также ос, пчёл и т. п.
В ряде случаев самцы воспринимают в качестве соперников неодушевлённые предметы соизмеримые с бабочками их вида размерами и цветом[72] — например, данное поведение отмечается у самцов морфо, хараксес, парусника Улисса.
Миграции

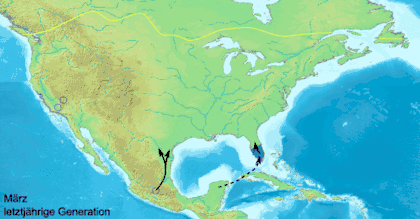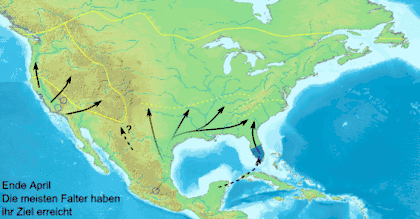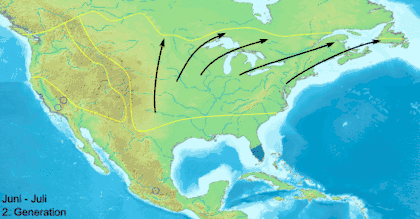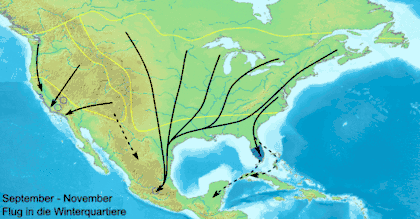
Миграционное поведение встречается у бабочек нечасто. Оно известно только примерно у 250 видов[73][74], причём всего два десятка из них совершают регулярные и продолжительные по расстоянию перелёты[73][74].
Мигрирующие бабочки совершают перелёты, как в одиночку, так и объединяясь в стаи. Виды, совершающие регулярные миграции, обычно чётко следуют определённому маршруту, который часто совпадает с направлением миграционных путей птиц.
Данаида монарх (Danaus plexippus) — самая известная мигрирующая бабочка, ежегодно в ходе миграции преодолевающая большие континентальные расстояния[25]. В Северной Америке данаиды-монарх мигрируют в южном направлении, начиная с августа до первых заморозков. Северные особи мигрируют в весенний период. К концу октября численность особей к востоку от Скалистых гор мигрирует в большинстве в убежище в Биосферный заповедник бабочки монарх, находящийся в мексиканском штате Мичоакан. На отдельных деревьях скапливается до 100 000 особей одновременно, а общее число мигрирующих особей может достигать 50 миллионов[25][27]. В ходе весенней миграции монарх летит от дальнего севера через восток Скалистых гор в штаты Оклахома и Техас. Второе, третье и четвёртое поколения весной возвращаются в северные районы США и Канады. Как различным поколениям удаётся вернуться в миграции на юг, туда, где зимовали их предки, до сих пор остаётся загадкой для учёных.
Среди африканских бабочек самые длинные миграции совершает белянка Catopsilia florella. Каждый год с декабря по февраль бабочки из засушливых областей Сахеля улетают на юг в Заир, с началом сезона дождей и появлением множества цветов, обеспечивающих бабочек пищей. С наступлением сухого сезона бабочки мигрируют назад в Сахель[75].
К миграциям способны и европейские виды, например, репейницы зимуют в Северной Африке, где они размножаются, и новое поколение репейниц мигрирует на север, где выводится летнее поколение бабочек. В конце лета бабочки этого поколения мигрируют обратно в Африку. Весной цикл повторяется снова. Протяжённость их перелёта достигает 5000 км. Репейницы, обитающие в Северной Америке и Австралии, также совершают осенне-весенние миграции, но маршруты этих миграций иные.
Одну группу составляют виды, совершающие перелёты нерегулярно, а в зависимости от условий. Это траурница, адмирал, крапивница, капустница, махаон. Все данные виды обитают и размножаются в Средней и Северной Европе, но в данные регионы летом регулярно мигрируют особи из южных районов[75]. Другую группу составляют виды (например, мёртвая голова (Acherontia atropos), олеандровый бражник), совершающие ежегодные миграции из южный регионов — Турция, Северная Африка — в Центральную и Восточную Европу, где они оставляют потомство, в большинстве случаев погибающее зимой. Новое поколение снова мигрирует весной в эти регионы с юга. Передвижения данных видов в умеренные широты скорее можно считать рассеивающим, чем миграционным.
Симбионты
Большое число видов чешуекрылых состоит в мутуализме с представителями различных биологических групп, от прокариот, таких как бактерии рода Wolbachia, и растений (юкковая моль и растения юкка, ангрекум полуторафутовый и бражник Xanthopan morganii praedicta) до собратьев по классу насекомых.
У ряда видов гусеницы живут в муравейниках, находясь с муравьями в симбиотических отношениях, например, с родом Myrmica[76]. Примерно половина всех видов голубянок (Lycaenidae) так или иначе связана в цикле своего развития с муравьями[76].

Взаимосвязи между муравьями и голубянками по своему типу варьируются от факультативных до облигатных, и от мутуализма до паразитизма[76]. Гусеницы и куколки голубянок обладают комплексом эволюционно выработанных химических и акустических сигналов для контролирования поведения муравьёв. Они также выделяют из дорзальной нектароносной железы сладкую жидкость, привлекающую муравьёв. Муравьи, ощупывая усиками этот орган, вызывают рефлекторное выделение гусеницей жидкости, которая, вероятно, содержит феромоны лихневмоны, обусловливающие поведение муравьёв. Некоторые гусеницы голубянок и риодинид обладают также звукоиздающими органами, влияющими на поведение муравьёв — одни виды имеют только шаровую щетинку на теле гусеницы, другие — вместе с звуковой ресничкой, при отсутствии нектароносной железы[76].
Все виды мирмекофильных голубянок приурочены к луговым муравейникам. Одним из таких видов является голубянка алькон (Maculinea alcon), самки которой откладывают яйца на цветы растений семейства горечавковые (Gentiana). Большинство видов голубянок развивается в гнёздах лишь одного вида муравьёв, но гусеницы голубянки алькон обитают в гнёздах муравьёв различных видов, на разных участках своего ареала[76][77]. Некоторые бабочки, такие как Mechanitis polymnia isthmia Bates, M. lysimnia doryssus Bates (Nymphalidae, Ithomiinae) и Graphium philolaus (Boisduval) (Papilionidae) привлекаются колоннами кочевых муравьёв Eciton burchelli (по наблюдениям в Гондурасе). По разным гипотезам, это может быть объяснено или запахами, сходными с половыми феромонами бабочек, или с запахами, связанными с разложившейся пищей (привлекающей муравьёв), либо с помётом птиц, следующих за колонами Eciton (по наблюдениям за теми же видами, плюс M. lilis imitata в Финка-ла-Сельва, Коста-Рика). Известно, что бабочки из подсемейства Ithomiinae используют мочевую кислоту или частично переваренные белки в птичьем помете для развития и откладки яиц[78].
Несколько видов чешуекрылых из группы Chrysauginae (семейство Огнёвки настоящие) как то Cryptoses choloepi или Bradipodicola hahneli являются комменсалами млекопитающих из семейства трёхпалые ленивцы (форезия). Когда ленивец спускается с дерева для дефекации, бабочки данных видов откладывают в его экскременты яйца, из которых затем выходят гусеницы, питающиеся ими (копрофагия)[79].
Природные враги
.JPG.webp)
Природными врагами чешуекрылых и преимагинальных стадий их развития в первую очередь являются насекомоядные позвоночные животные: птицы, ящерицы, жабы, грызуны, насекомоядные млекопитающие[80]. Кроме насекомоядных позвоночных, на бабочек охотятся насекомые и пауки[80]. К таким хищникам, среди насекомых, относятся ктыри и стрекозы, которые охотятся на бабочек во время полёта, богомолы и пауки-бокоходы, которые охотятся из засады на цветках, привлекающих бабочек[80]. Пойманных бабочек часто можно наблюдать в паутине пауков. Охотятся на чешуекрылых также жуки-жужелицы и некоторые осы.
Чешуекрылые страдают от микозов и вирусов (например, вирус ядерного полиэдроза)[80]. Для каждого вида чешуекрылых свойственен свой собственный специфичный видовой вирус[81]. Например, вирус Baculovirus reprimens — специфичен только непарному шелкопряду, Baculovirus srilpnotiay — ивовой волнянке, Baculovirus neustriae — кольчатому коконопряду и т. д. Насчитывается более 100 видов чешуекрылых — хозяев этой группы возбудителей полиэдрозов общего типа (вирусы рода Baculovirus)[81]. При попадании в кишечник гусениц полиэдры под воздействием кишечного сока растворяются, высвобождая вирионы, которые распадаются «провирусы». Они проникают сквозь стенки кишки в миксоцель, разносятся гемолимфой по всему телу, проникая в восприимчивые ткани различных органов. Поражаются клетки жирового тела, эпителий трахей, гиподерма, нейроны, гонады, мышечная ткань. Патогенез вируса заканчивается гибелью гусениц. Их внутренние органы и ткани лизируются, превращаясь в беловато-желтоватую жидкость, содержащую вирусные агенты[81].
Паразиты и паразитоиды
Представители мирмекофильных разнокрылых бабочек рода Cyclotorna (австралийские эндемики из надсемейства Zygaenoidea), являются единственными среди бабочек эктопаразитами[82]. Гусеницы (первоначально похожие на уплощённых мокриц, а на последних стадиях более ярко окрашенные) являются эктопаразитами равнокрылых насекомых. Самки бабочек откладывают до 1400 яиц на растения, поражённые цикадками (Cicadellidae) или листоблошками (Psyllidae). Молодые гусеницы первого возраста находят свою жертву и высасывают из неё гемолимфу. Гусеницы Cyclotorna в итоге покидают своих хозяев и становятся хищниками личинок мясных муравьёв (Iridomyrmex purpureus), по-видимому, используя химические вещества (феромоны), индуцируют муравьёв для переноски гусениц в свои муравейники (перед окукливанием покидая их гнёзда)[82].
Гусеницы Zenodochium coccivorella и Euclemensia bassettella являются внутренними паразитами самок семейства кермесов (Coccidae)[83][84].
Среди настоящих паразитов, специализирующихся на чешуекрылых, следует отметить, прежде всего, одноклеточных, например, микроспоридии родов Thelohania и Plistophora, и отдельные виды грибов, например, Cordyceps sinensis и другие представители рода Cordyceps.

Существует также большое число организмов, развивающихся за счёт чешуекрылых, но не являющихся истинными паразитами, так как в результате их жизнедеятельности организм-хозяин в любом случае либо гибнет, либо теряет способность к продолжению рода. Принятое в науке название представителей группы, обладающей описанной формой биологических взаимоотношений, — паразитоиды. На различных стадиях жизненного цикла бабочки подвержены нападению определённых типов паразитоидов: яйцевых, яйцеличиночных, личиночных, личиночно-куколочных и куколочных.
Многие представители браконид (Braconidae), одного из семейств наездников, паразитируют на гусеницах[85]. Как и большинство других наездников, бракониды в основном откладывают яйца в свободно передвигающихся гусениц, некоторые — предварительно парализуют жертву, частично либо полностью. Последнее характерно для видов, специализирующихся на скрытоживущих хозяевах, вынужденных отыскивать и доставать при помощи своего длинного яйцеклада гусениц, например, под корой деревьев. Длительный или необратимый паралич наступает, как правило, от вводимого наездником в тело хозяина избыточного количества токсина.
Из браконид наибольшее значение в России имеют Apanteles glomeratus — главный истребитель белянок, Habrobracon juglandis — паразит мучной огнёвки, Apanteles solitarius и Meteorus versicolor — паразиты ивовой волнянки. Бракониды рода Microgaster заражают гусениц капустницы (Pieris brassicae), которые после перехода в стадию куколки поражаются представителями другого семейства — птеромалид (Pteromalidae) — Pteromalus puparum[85][86].
Самки наездников-ихневмонид (Ichneumonidae) из отряда перепончатокрылых, принадлежащие исключительно к личиночным и яйцеличиночным паразитам[87], откладывают свои яйца под кутикулу гусениц или непосредственно на неё. В последнем случае вылупившиеся личинки сами внедряются внутрь хозяина. Обычно молодые личинки питаются гемолимфой, а в конечных стадиях своего развития переходят на питание тканями и внутренними органами гусеницы. Окукливание происходит в теле гусеницы либо вне его[29][88].
На гусеницах также паразитируют двукрылые, прежде всего тахины, или ежемухи (Tachinidae)[89]. Их самки откладывают яйца или личинок, в случае живорождения, непосредственно на гусеницу. Другие виды ежемухи могут откладывать яйца на листву кормового растения гусениц. У гусениц, съевших их вместе с листвой, выход личинок происходит в пищеварительном тракте хозяина, откуда в дальнейшем они внедрятся в полость тела. Окукливание происходит внутри хризалиды, либо, в случае гибели гусеницы — в почве[29].
Защита от хищников
Окраска и внешний вид многих бабочек и их преимагинальных стадий развития тем или иным способом связаны с защитой от врагов. Методы защиты от хищников очень разнообразны. Ряд видов обладают скверным запахом и неприятным вкусом, либо ядовиты, всё это делает их несъедобными. Попробовав такую бабочку один раз, хищники будут избегать подобный вид в дальнейшем. Так, например, геликониды в стадии гусеницы поглощают в составе листьев кормовых растений гликозиды с цианистой группой, из которых, в процессе метаболизма, образуются цианиды, передающиеся и имаго бабочки[37]. Другие виды также несут в себе токсичные соединения: алкалоиды, гликозиды, производные фенолов[11]. Бабочкам рода Ornithoptera, гусеницы которых питаются на лианах рода кирказон, передаётся также аристолохиевая кислота[11].
Ядовитые и несъедобные виды часто обладают предостерегающей яркой окраской. Бабочки, лишённые таких средств защиты, часто мимикрируют под несъедобные виды, «подражая» не только окраске, но и форме крыльев. Данный вид мимикрии наиболее развит именно у чешуекрылых и называется «бейтсовской»[37]. Так, южноамериканские парусники Papilio bachus, Papilio zagreus, белянка Dismorphia astynome окраской и формой крыльев подражают несъедобным видам геликонид. Парусник Papilio agestor подражает несъедобной данаиде сита (Paratica sita)[37].
.jpg.webp) |
.jpg.webp) |
.JPG.webp) |
|||
| Отпугивающая окраска в виде «глаз» на крыльях. Сатурнии Automeris io. | Предостерегающая яркая окраска ядовитых и несъедобных видов. Eueides isabella. | Покровительственная окраска. Ленточница малиновая на коре дерева. | |||
 |
 |
 |
|||
| Мимикрия насекомым других отрядов. Шмелевидка жимолостная подражает шмелям. | Мимикрия отмершим сухим частям растений и деревьев. Коконопряд дуболистный подражает сухому листу. | Мимикрия несъедобным видам. Papilio laglaizei (на фото) мимикрирует Alcides agathyrsus. |
При «мюллеровской» мимикрии подражающие друг другу виды в одинаковой мере несъедобны. Сходная окраска и внешний вид повышают эффективность отпугивания[37]. Одним из таких примеров является дневной парусник Papilio laglaizei (Новая Гвинея), подражающий окраской и формой крыльев ночной урании Alcides agathyrsus. Оба вида несъедобны[16][27][37].
Африканский парусник Papilio dardanus характеризуется полиморфизмом. Известно около 40 различных форм его самок — в соответствии с числом видов, которым они подражают. У самок данного вида отсутствует единый вариант окраски, что ещё больше повышает шансы на выживание[37].
Некоторые бабочки подражают осам и шмелям, например, стеклянницы (Sesiidae), бражник шмелевидка жимолостная (Hemaris fuciformis), шмелевидка скабиозовая (Hemaris tityus). Данное сходство достигается благодаря окраске, контурам тела и структуре крыльев — они почти лишены чешуек и прозрачные, задние крылья короче передних, а чешуйки на них сосредоточены на жилках[37]. Бабочка Brenthia hexaselena из семейства риодинид (Riodinidae), обитающая в Коста-Рике, будучи потревоженной, своеобразно складывает крылья и своими движениями напоминает пауков из семейства Salticidae.
_at_Samsing%252C_Duars%252C_West_Bengal_W_IMG_6238.jpg.webp)
.jpg.webp)
Многие виды имеют покровительственную окраску и маскируются под сухие листья, веточки, кусочки коры[27]. Бабочки из рода Kallima из Юго-Восточной Азии, имеющие ярко окрашенную верхнюю сторону крыльев, садясь на ветку и складывая крылья, принимают вид сухого листа: короткими выростами задних крыльев бабочка упирается в ветку, и они представляют сходство с черешком; рисунок же и цвет нижней стороны сложенных крыльев в такой степени напоминают цвет и жилкование засохшего листа, что даже на близком расстоянии бабочку чрезвычайно трудно отличить от листьев[16][27][37]. Другие виды — например, лунка серебристая (Phalera bucephala) — напоминают отломанный сучок; коконопряд дуболистный (Gastropacha quercifolia) походит на сухой берёзовый лист.
В отличие от бабочек, активных в светлое время суток, виды, активные в сумерках или ночью, обладают иной защитной окраской. Верхняя сторона их передних крыльев окрашена в цвета субстрата, на котором они сидят в покое. При этом их передние крылья складываются вдоль спины наподобие плоского треугольника, прикрывая нижние крылья и брюшко[27].
Одним из видов отпугивающей окраски являются «глазки» на крыльях. Они располагаются на передних и/или задних крыльях и имитируют глаза позвоночных животных. В спокойном состоянии бабочки с такой окраской обычно сидят, сложив крылья, а будучи потревоженными, раздвигают передние крылья и демонстрируют отпугивающие ярко окрашенные нижние крылья. У некоторых видов, например бабочек рода Калиго, на нижней стороне крыльев отчётливо выделяются большие и очень яркие тёмные округлые пятна с бело-жёлтым контуром, похожие на глаза совы[11][16][25][27][53].
Ночные бабочки для защиты от летучих мышей обладают густо опушёнными волосками телом. Волоски помогают поглощать и рассеивать ультразвуковые сигналы летучих мышей и тем самым маскировать местонахождение бабочки[30][54]. Многие бабочки замирают, когда обнаруживают сигнал сонара летучей мыши. Медведицы способны генерировать серии щелчков, которые по мнению ряда исследователей также препятствуют их обнаружению[30].
Бражник мёртвая голова, будучи потревоженным, издаёт пронзительный писк путём колебании выроста верхней губы эпифаринкса, засасывая воздух в глотку, а затем продавливая его обратно[11].

Роль в природе
Роль бабочек в естественных экосистемах заключается в том, что, будучи консументами первого порядка, они являются неотъемлемыми участниками пищевых цепей. В процессе коэволюции организмов сформировалась трофическая сеть связей между автотрофами и гетеротрофами, в которую чешуекрылые включаются на стадиях личинки, куколки и имаго. Личинки и куколки являются звеньями в питании птиц и паразитических насекомых-энтомофагов. Имаго включаются в трофическую сеть значительно более широкого спектра консументов второго порядка — это разнообразные птицы, мелкие млекопитающие, пресмыкающиеся и т. п.
Бабочки также выступают опылителями многих видов цветковых растений, ведь значительная часть этих насекомых характеризуется развитием антофилии. В ходе эволюции бабочки приспособились к определённым цветковым растениям, а ряд видов растений приспособились к опылению исключительно бабочками. Например, только юкковой молью (Pronuba juccasella) опыляются растения вида Юкка нитчатая (Jucca filametosa). Мадагаскарская орхидея Ангрекум полуторафутовый (Angraecum sesquipedale) с очень глубокой цветочной чашечкой опыляется единственным эндемичным видом бражников Xanthopan morgani, имеющим хоботок длиной 225 мм[11].
Филогения, систематика и классификация
Систематическое положение
Филогенетически чешуекрылые близки к ручейникам (Trichoptera), вместе с которыми они образуют надотряд Amphiesmenoptera. Представители обоих отрядов имеют целый ряд общих признаков: самки, а не самцы являются гетерогаметными, что не характерно для других отрядов насекомых; плотные волоски на крыльях; специфическое жилкование передних крыльев; наличие у личинок желез, вырабатывающих шёлковую нить. Надотряд Amphiesmenoptera, вероятно, начал эволюционировать в юрском периоде и отделился от вымерших Necrotaulidae[90]. Чешуекрылые, в свою очередь, отличаются от ручейников некоторыми особенностями жилкования крыльев; формой волосков, модифицированных в чешуйки[90].
Филогения и эволюция
Древнейшие ископаемые остатки чешуекрылые, известные под названием Archaeolepis mane, найдены в отложениях раннего юрского периода вблизи Дорсета, Великобритания, их возраст составляет около 190 млн лет[90][91]. Наибольшее развитие группы совпадает с расцветом цветковых растений в меловой период[90], когда происходит развитие и становление чешуекрылых, с активными растительноядными личинками, питавшимися зелёными тканями растений, и крылатой половозрелой стадией жизненного цикла, первоначально обладавшей малой активностью и имевшей слабо развитый ротовой аппарат.
Предполагаемая филогения основных групп бабочек[92] |
Чешуекрылые, с чертами начальной дезимагинизации, в меловом периоде образовали ряд форм, у которых процессы дезимагинизации затормозились. У данных чешуекрылых произошло удлинение ротовых органов, с образованием длинного хоботка, способного всасывать жидкую пищу, что позволило активно питаться нектаром. Продолжительность жизни крылатой стадии увеличивалась, происходило усовершенствование крыльев и полёта, увеличение абсолютных размером. Чешуекрылые также приобрели разнообразную яркую окраску тела и, в первую очередь, крыльев, на которых стали возникать сложные рисунки.
Возникновение такой яркой окраски, в некоторой степени, отражает их тесные взаимосвязи с цветковыми растениями, цветки которых бабочки посещали для питания нектаром, тем самым, участвуя в процессах опыления. Темпы эволюции чешуекрылых были высоки. Параллельное развитие чешуекрылых и цветковых растений является ярким примером сингенетического развития двух групп организмов, которое происходило относительно быстрыми темпами в каждой из названных групп организмов[93].
Остатки ископаемых бабочек из палеогена представлены находками преимущественно внутри янтаря. Большинство найденных таким образом видов относится к современным семействам.
Самым древним видом ныне живущих булавоусых чешуекрылых, по-видимому, является Baronia brevicornis — реликтовый вид, эндемик Мексики[94]. Он характеризуется общими чертами с ископаемым таксоном Praepapilio и считается самым примитивным сохранившихся видом группы Papilionidae[94].
Сбоку на схеме показана предполагаемая филогения основных групп бабочек.
Классификация
Классификация отряда разработана не окончательно и ранг многих таксонов (даже выше уровня семейства) является порой дискуссионным. В мировой фауне, по разным оценкам, насчитывается от 124 до 200 семейств чешуекрылых, в фауне России — 91[15][95].
Современная классификация чешуекрылых выделяет четыре подотряда:
- Первичные зубатые моли (Zeugloptera) — в настоящее время представлен одним надсемейством Micropterigoidea, в котором имеется единственное одноимённое семейство (Micropterigidae);
- Бесхоботковые (Aglossata) — в настоящее время представлен одним надсемейством Agathiphagoidea и семейством Agathiphagidae;
- Heterobathmiina — в настоящее время представлен одним надсемейством Heterobathmioidea, в которое включено единственное семейство Heterobathmiidae;
- Хоботковые (Glossata) — самый многочисленный подотряд.
Можно встретить и другие (не вполне отвечающие современной научной классификации) варианты деления отряда чешуекрылых на группы. В частности, выделяют четыре большие группы[11]:
- низшие разноусые бабочки,
- огнёвкообразные бабочки,
- дневные, или булавоусые бабочки,
- высшие разноусые бабочки.
Последнюю группу часто называют ночными, что не совсем корректно, так как многие из них ведут дневной образ жизни. Булавоусые бабочки отличаются от разноусых по ряду характерных признаков: форма усиков, механизм крепления крыльев к груди, окраска, форма тела и время наибольшей активности. Для дневных характерны булавовидные усики. Высшие разноусые бабочки, большинство из которых активно в тёмное время суток, имеют нитевидные или перистые усики. Механизм крепления передних и задних крыльев к груди различен — булавоусые бабочки способны складывать крылья «за спиной», в то время как разноусые бабочки оставляют крылья раскрытыми плашмя или складывают их «домиком»[28]. Однако существуют и исключения. Известны булавоусые бабочки, находящиеся в покое с распростёртыми крыльями, обладающие толстым брюшком и активные в тёмное время суток и, наоборот, разноусые с яркой окраской, тонкими булавовидными усиками, стройным брюшком, и активные в светлое время суток.
Замечания по охране
В результате хозяйственной деятельности человека — вырубки лесов, распашки целинных степей, осушения болот — численность многих видов бабочек значительно сократилась, а некоторые из них находятся на грани вымирания. На размерах популяции бабочек сказываются малейшие изменения в видовом разнообразии растительности мест их обитания. В первую очередь в группе риска находятся бабочки, обитающие на ограниченной территории, не склонные к миграциям, гусеницы которых питаются растениями только одного вида. В качестве примера можно привести европейскую брамею (Acanthobrahmaea europea), обитающую только в прибрежном лесу вулканического озера в Лукании (Италия)[96].

Для привлечения внимания к необходимости охраны бабочек в Красные книги разных стран заносятся виды, находящиеся под угрозой исчезновения. Красная книга служит научно-директивной основой организации охраны редких и находящихся под угрозой исчезновения видов. В Красный список угрожаемых видов (The IUCN Red List of Threatened Species), публикуемый Международным союзом охраны природы и природных ресурсов (МСОП) внесено 217 редких и исчезающих вида чешуекрылых, из них 27 видов — вымершие, 11 видов — находящиеся на грани исчезновения, 51 видов — вымирающие, 128 видов — уязвимые. Всего на сайте МСОП перечислено 786 видов чешуекрылых с учётом видов, информации о которых недостаточно (Data Deficient, 87 видов), близки к уязвимому положению (Near Threatened, 62 видов) или чьё положение под наименьшей угрозой (Least Concern, 420 видов)[97][98].
Важным инструментом сохранения редких видов животных и растений в Европе является «Бернская конвенция об охране дикой фауны и флоры и природных сред обитания», принятая в 1979 году. В Приложение II Бернской конвенции занесён 41 вид бабочек[80]. Первая Красная книга России, вышедшая в 1984 году, включала 12 видов бабочек. Через два года вышла Красная книга СССР со списком из 105 видов чешуекрылых[99]. В 2001 году в составе Красной книги России был опубликован новый перечень находящихся под угрозой исчезновения животных России, куда попали 33 вида чешуекрылых[100].
С каждым годом всё больше стран объявляют о запрете неконтролируемого отлова бабочек на своей территории. Строже всего охраняются многие виды парусников: орнитоптеры, аполлоны, тейнопальпусы и другие. В Австралии за поимку парусника Улисс (Papilio ulysses) без соответствующего разрешения грозит лишение свободы на срок от 6 месяцев до 2 лет. Ещё бо́льшие сроки тюремного заключения грозят ловцам орнитоптер в Папуа — Новой Гвинее[96]. На Мадагаскаре и Цейлоне введены специальные денежные вознаграждения для населения при сообщении в местные органы по охране общественного порядка о фактах незаконного лова бабочек приезжими ловцами[101].
В последние десятилетия были разработаны и приняты законы, направленные на сохранение ряда насекомых, в том числе и чешуекрылых. Международный союз охраны природы (The International Union for Conservation of Nature — IUCN) выпустил несколько Красных книг со списками чешуекрылых, отлов которых запрещён или существенно ограничен. Эти списки признаны более чем в ста странах мира, в том числе и в России.

Перечень чешуекрылых экспорт, реэкспорт и импорт которых регулируется в соответствии с Конвенцией о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (СИТЕС)[102]:
- Atrophaneura jophon
- Atrophaneura pandiyana
- Bhutanitis spp.
- Ornithoptera alexandrae
- Ornithoptera spp.
- Papilio chikae
- Papilio homerus
- Papilio hospiton
- Parnassius apollo
- Teinopalpus spp.
- Trogonoptera spp.
- Troides spp.
- Agrias amydon boliviensis
- Morpho godartii lachaumei
- Prepona praeneste buckleyana
Наиболее надёжную охрану бабочек и мест их обитания могут обеспечить заповедники, заказники, национальные парки[25]. В ряде стран в местах размножения мигрирующих бабочек создаются охраняемые временные заповедники, например, на острове Родос (городок Салакос, Долина бабочек), где откладывают яйца самки медведицы Геры (Callimorpha quadripunctaria), и на путях миграции монарха в США[96].
Чешуекрылые и человек
Использование в пищу

Энтомофагия (поедание насекомых) является широко распространённым явлением по всему миру. Гусеницы и куколки представителей примерно восьмидесяти родов из двадцати семейств чешуекрылых употребляются в пищу[103], как богатые белком пищевые продукты. Их едят в сыром или жареном виде, и они употреблялись человеком в пищу с доисторических времён. Например, индейцы Северной Америки в Каскадных горах и горах Сьерра-Невады собирали гусениц сатурнии пандоры (Coloradia pandora), которых подсушивали на слое горячих углей или отваривали[103]. Австралийские аборигены с давних времён употребляют в пищу гусениц древоточца Xyleutes leucomochla, которых добывают, выкапывая корни акациевых кустов и разрубая их, чтобы добраться до находящихся в них насекомых[103][104].
В наше время в Азии куколок тутового шелкопряда, размотав шёлк с кокона, не выбрасывают, а употребляют в жареном или засахаренном виде. В Китае гусениц тутового шелкопряда засаливают, добавляют в омлет из куриных яиц, либо просто жарят с луком и густыми соусами. В 1987 году в Таиланде Министерство здравоохранения включило куколок тутового шелкопряда в перечень продуктов, которые могут дополнять разработанный рацион для детей дошкольного возраста[103] — куколки рекомендовалось прожаривать и слегка измельчать для последующего использования в составе карри и первых блюд. Во Вьетнаме с ними варят местную разновидность капустного супа. Южная Корея поставляет консервированных куколок тутового шелкопряда в азиатские магазины по всему миру[103]. В Индии ежегодно потребляется более 20 000 тонн этих насекомых[37]. В Африке деликатесом считаются жареные или копчёные гусеницы сатурнии Gonimbrasia belina, которые стоят в 4 раза дороже обычного мяса[37]. В отдельных районах Северной Африки эти гусеницы настолько популярны, что в сезон существенно падают продажи говядины и мяса других животных. В Ботсване их продают большими пакетами в сушеном виде, как любые другие сухие продукты. В Мозамбике и Зимбабве сельские женщины собирают гусениц Gonimbrasia belina ранней весной и тушат их с овощами либо сушат[103].
Прочее использование
На Соломоновых островах местные жители делали краску из бабочек с жёлтыми крыльями, которой окрашивали изделия, плетённые из травы[105].
В последнее время всё большую популярность приобретают живые тропические бабочки в качестве нестандартного и экзотического подарка на многие праздники и торжества, как то юбилей или свадьба. Специально для этой цели по всему миру появляются частные компании, занимающиеся искусственным разведением бабочек.
Также с увеличением числа коллекционеров в ряде стран стали создаваться фермы по разведению бабочек. Наиболее известные из них — остров Пинанг в Малайзии, Булоло на Новой Гвинее, Сатипо в Южной Америке. Выставки живых тропических бабочек имеются во многих крупных городах мира, и их число постоянно увеличивается[11].
Бабочки активно используются в энтомодизайне — оформлении интерьеров жилых и рабочих помещений профессионально собранными коллекциями насекомых[106]. Особым видом последнего является использование засушенных бабочек, находящихся внутри специального стекла, при создании витражей окон и межкомнатных дверей.
Чешуекрылые и здоровье человека
Гусеничный дерматит — энтомоз, обусловленный раздражением кожи волосками гусениц некоторых видов. Заболевание возникает при проникновении в кожу щетинок и волосков гусеницы, содержащих токсические вещества, аналогичные по своему химическому составу кантаридину. Развивается также при воздействии веществ, выделяемых железистыми клетками покровов гусеницы. Сопровождается сильными субъективными ощущениями и развитием воспалительно-экссудативной реакции кожных покровов: на поражённых участках кожи возникают ярко-красные зудящие высыпания, на периферии которых отмечаются зудящие папуло-везикулезные элементы. Наибольшим раздражающим свойством обладают гусеницы Taumatopea processionopea.
В нетрадиционной медицине для лечения целого ряда заболеваний используется спиртовой настой из окукливающихся гусениц пчелиной огнёвки[107].
Экономическое значение
Для человека в первую очередь полезны виды, гусеницы которых производят шёлк. В природе шёлк образуют гусеницы многих видов бабочек, сооружая из него коконы. Текстильная промышленность отдаёт предпочтение тутовому, или шелковичному, шелкопряду (Bombyx mori), одомашненному человеком. В шелководстве также используются китайская дубовая павлиноглазка (Antheraea pernyi), которую уже более 250 лет разводят в Китае. Из её коконов получают шёлк, который идёт на изготовление чесучи. Другие виды шелкопрядов плохо развиваются в неволе, поэтому ограничиваются лишь сбором их коконов в природе[37]. Тутовый шелкопряд играет важную экономическую роль в производстве шёлка. Для получения шёлковой нити куколок предварительно умертвляют с помощью горячего пара и воды на десятый день после окукливания. В шёлковом коконе обычно содержится до 3500 м волокна, но размотать его удаётся едва на треть[37]. Чтобы получить 1 килограмм шёлка-сырца, нужны коконы примерно тысячи гусениц, поедающих за полтора месяца 60 килограмм листьев тутовника. Сегодня во всём мире ежегодно производится 45 000 тонн шёлка[37]. Главными поставщиками являются Китай, Индия, Узбекистан[108]. В Индии разводится сатурния Attacus atlas, гусеницы которой выделяют шёлк отличающийся от такового у тутового шелкопряда — он коричневого цвета, более крепкий и шерстистый — и носит название фагаровый шёлк (fagara silk)[109].

Гусеницы некоторых видов могут использоваться в борьбе с сорными растениями. Самым ярким примером служит специально завезённая в 1925 году в Австралию из Уругвая и из северных районов Аргентины кактусовая огнёвка (Cactoblastis cactorum)[110]. Гусеницы этой бабочки помогли избавиться от завезённого кактуса опунции, которым заросли миллионы гектаров пастбищ. В 1938 году в долине реки Дарлинг австралийские фермеры поставили специальный памятник гусеницам, спасшим Австралию[111].
Вредители

Очень велико хозяйственное значение чешуекрылых как вредителей сельского и лесного хозяйства. На территории стран СНГ зарегистрировано свыше 1000 видов чешуекрылых, гусеницы которых повреждают полевые, садовые или лесные культуры[89].
Полеводству вредят озимая совка (Agrotis segetum), восклицательная совка (A. exclamationis), яровая совка (Amphipoea fucosa), совка гамма (Plusia gamma). Вред огородам наносят капустница (Pieris brassicae), огородная совка (Mamestra oleracea), гороховая совка (М. pisi)[89]. Вредит лесам и садам — дубовый походный шелкопряд (Cnethocampa processionea), непарный шелкопряд (Lymantria dispar), сосновая совка (Panolis flammea), сосновый шелкопряд (Dendrolimus pini), зимняя пяденица (Operophthera brumata), яблонная плодожорка (Cydia pomonella), восточная плодожорка (Grapholita molesta), горностаевая черёмуховая моль (Yponomeuta evonymella), сосновый бражник (Sphinx pinastri) и многие другие. Амбарам и кладовым вредит амбарная моль (Nemapogon granella), мучная огнёвка (Pyralis farinalis) и бесхоботная огнёвка (Aglossa pinguinalis). Мехам, одежде из натуральных тканей и мебели вредят шубная моль (Tinea pellionella), платяная моль (Tineola bisselliella) и мебельная моль (Tineola furciferella). Пчеловодству вредит малая пчелиная огнёвка и большая восковая моль[24]. Гусеницы походных шелкопрядов (Thaumetopoeidae) отличаются способностью к длительным миграциям в поисках пищи, и при массовом размножении могут выедать целые гектары лесов и лесных массивов.
Известны факты завоза вредных чешуекрылых из других стран. В Европе широко распространилась американская белая бабочка (Hyphantria cunea), родина которой — Северная Америка. На Европейском континенте она впервые была обнаружена в 1940 году в Венгрии, а через несколько лет она распространилась на территорию других стран: от Франции до Каспийского моря. В Японии она известна с 1945 года, откуда проникла также в Корею, Китай, Монголию и Приморский край[89].
С целью борьбы с гусеницами-вредителями хозяйственных растений и лесного хозяйства используется как классическая фумигация инсектицидами, так и биологические, а также биохимически-физиологические методы. Одновременно, разработка таких методов предоставляет материал для научных исследований более фундаментального характера.
Чешуекрылые в культуре
Верования, мифы, символизм
С древних времён разные народы связывали бабочек с любовью, душой, возрождением.
- В эпоху палеолита одним из насекомых, представлявших богиню «Великую мать», была бабочка[112].
- У ацтеков бабочка была одним из атрибутов бога растительности, весны и любви Шочипильи. По поверьям южноамериканских индейцев морфо — это души умерших людей, стремящиеся ввысь, а местное название этих бабочек можно перевести как «частица неба, упавшая на землю»[112].
- Древние римляне считали, что бабочки — это цветы, которые сорвал ветер. Они также называли бабочек «feralis» — «свирепая», и верили, что они могут предвещать войны[112].
- Древние греки считали бабочку символом бессмертия души. Психея (с греч. — «душа») изображалась в образе девушки с крыльями бабочки[112].
- В Китае бабочка является воплощением бессмертия, изобилия, влюблённости, лета. Бабочка, изображённая со сливой, символизировала долгожительство и красоту человека, с хризантемой — красоту в старости, а с пером — долголетие. Образ бабочки часто встречается в даосских притчах. И сегодня, жених перед свадьбой дарит невесте живую или нефритовую бабочку как символ любви[112].

- С глубочайшим почтением относятся к бабочкам буддисты: ведь именно к бабочке обратился Будда со своей проповедью[112].
- В Японии нимфалида Sasakia charonda является символом страны. По верованиям японцев, бабочка символизирует молодую женщину. Порхающие друг вокруг друга бабочки символизируют семейное счастье, а белая бабочка — душу умершего. Именно поэтому все торжественные шествия и праздники в этой стране начинаются с ритуального «танца бабочек», выражающего радость жизни[112].
- У христиан стадии жизненного цикла бабочки олицетворяли жизнь, смерть и воскресение, поэтому бабочку иногда изображали в руке младенца Христа как символ возрождения и воскресения души[112].
- Древние славяне связывали с бабочками представление о душе, в том числе и о душах умерших, предвестниц смерти[112]. В ряде случаев у славян бытовало поверье о душе ведьмы в виде бабочки; а южные славяне часто называли ночных бабочек «ведьмами». Болгары веровали, что ведьмы напускали на скот больших пёстрых бабочек, которые отбирали у них молоко[11][113]. Наибольшей дурной славой у разных народов, пожалуй, пользовался бражник мёртвая голова. С мрачным рисунком в виде черепа на его спинке связано немало легенд — так, по поверью, эта бабочка была предвестником несчастий, смерти, войн и эпидемий. Так, эпидемию 1733 года народ приписывал появлению этой бабочки[112]. В Иль-де-Франс до сих пор верят, что чешуйка с крыльев этой бабочки, попав в глаз, причиняет слепоту и возможную скорую гибель[11].
У славян представление об этих насекомых как о душе отразилось не только на его диалектных названиях — «душа» и «душечка», но и общеупотребительном названии представителей данного отряда — «бабочка», восходящем к праславянскому «baba», означающее в древности предка[11][114].
Изобразительное искусство, литература, поэзия

Образ бабочек встречается в произведениях множества художников, писателей и поэтов со всего мира. Художественные изображения бабочек с давних времён существуют во многих мировых культурах. Первые из них известны по древнеегипетским фрескам, возрастом 3500 лет[112]. Также имеются многочисленные картины, например, «Охотник за бабочками» Карла Шпицвега, «Бабочки» В. фон Каульбаха (ок. 1860), «Бабочки» В. Д. Поленова, являющиеся наглядными изображениями бабочек.
В мае 1889 года Ван Гог написал картину, изображающую бабочку, названную им «Бражник мёртвая голова». Ван Гог ошибочно посчитал, что перед ним эта редкая в Европе бабочка. На самом же деле на картине изображена Павлиноглазка малая (Saturnia pavonia). Музей ван Гога в Амстердаме переименовал картину в «Императорская ночная бабочка» («Павлиноглазка»)[115][116].
Мимо образа бабочек не прошли и многие писатели, в том числе и великий американский писатель Эдгар По. Прекрасная и жутковатая бабочка бражник мёртвая голова описана им в рассказе «Сфинкс». Бабочка ползала по паутине на окне, а герою рассказа казалось, что существо двигается по далёким склонам холмов.

Бабочка с прозрачными крыльями Hetaera esmeralda стала центральным символом романа Томаса Манна «Доктор Фаустус». Герой, композитор Адриан Леверкюн, отождествляет эту бабочку с любимой женщиной, роман с которой оказался пагубным для его жизни, но послужил вдохновлением для творчества.
И всё же, в литературе бабочки чаще выступали в качестве символа свободы, любви и надежды. Так воспринимали этих насекомых русские поэты Василий Жуковский, Иван Бунин, Афанасий Фет и другие.
Ты прав. Одним воздушным очертаньем
Я так мила.
Весь бархат мой с его живым миганьем —
Лишь два крыла
Не спрашивай: откуда появилась?
Куда спешу?
Здесь на цветок я лёгкий опустилась
И вот — дышу.
Надолго ли, без цели, без усилья,
Дышать хочу?
Вот-вот сейчас, сверкнув, раскину крылья
И улечу.
Кино и музыка
Бабочки в качестве центральных образов фигурируют в целом ряде кинофильмов. Например, «Голубая бабочка» (The Blue Butterfly) 2004 года, режиссёра Леа Пул, в котором умирающий от рака ребёнок просит энтомолога поехать с ним в джунгли Южной Америки, где водятся бабочки морфо, которые, согласно легендам, обладают волшебными свойствами исцелять больных. Фильм «Бабочка» (фр. Le papillon) 2002 года режиссёра Филиппа Мюиля рассказывает о том, как ворчливый пожилой коллекционер бабочек вместе с маленькой дочерью своей соседки отправляется на поиски уникального экземпляра для своей коллекции — бабочки Graellsia isabellae, которая по красоте и редкости не имеет себе равных.

Определённую роль играет бабочка бражник мёртвая голова в фильме режиссёра Джонатана Демми «Молчание ягнят», снятом по мотивам одноимённого романа Т. Харриса. Маньяк-убийца кладёт куколку этой бабочки в рот своим жертвам.
Во многих кинофильмах рамки с энтомологическими коллекциями бабочек часто украшают стены в домах персонажей. Таким образом подчёркиваются положительные стороны героя, его стремление к прекрасному, либо злодейство — убийство беззащитных насекомых. Например, в кинофильме «Коллекционер» (англ. The Collector) 1965 года режиссёра Уильяма Уайлера, снятому по одноимённому роману Джона Фаулза, банковский клерк коллекционирует бабочек и знает про них всё, но у него не получается строить отношения с девушками, и однажды он похищает одну из них. Фильм «Ангелы и насекомые» (Angels and Insects) 1995 года режиссёра Филипа Хааса повествует об энтомологе, узнающем, что его жена состоит в кровосмесительной связи с двоюродным братом.
Бабочки также становятся частыми персонажами анимационных фильмов: во французском анимационном телесериале Minuscule дневная бабочка предстаёт изящной и аристократичной, а ночная моль — любвеобильной.
Образ бабочек возникает и в популярной музыке: существует американская рок-группа «Железная бабочка» (Iron Butterfly), а также песни Scorpions — «Yellow Butterfly», Crazy Town — «Butterfly», Майкл Джексон — «Butterflies», HIM — «Wings of a butterfly», Diary of Dreams — «Butterfly Dance», и многие другие.
В геральдике

В геральдике бабочки обычно являлись символом непостоянства, а их крылья в ряде случаев могли украшать щит[117].
В Российской империи Ферганская область, а также более двух десятков городов и районов имели или имеют изображение имаго бабочек или гусениц на своих гербах[117][118]. Причём к «говорящим гербам» можно отнести таковые городов Ферганской области, через которые проходил Великий шёлковый путь.
В современной культуре
Бабочки являются популярными объектами в современной культуре. Их изображения, в той или иной интерпретации, часто появляются как и на обложках научных книг, посвящённых природе, так и художественных произведений; на рекламных плакатах и постерах кинофильмов, на обложках музыкальных дисков и в музыкальных видеоклипах, на керамических и ювелирных изделиях.
Образ бабочек получил своеобразное преломление и в татуировках, которые в первую очередь являются женскими и очень разнообразны по формам и размерам. Они могут обладать различным символизмом, в зависимости от культурных и самобытных особенностей человека. В западной культуре такие татуировки обычно являются символом утончённой красоты, а у азиатских народов — символом удачи или свободы[119].
Коллекционирование бабочек

Бабочки являются наиболее популярной группой насекомых для частных коллекционеров и научных работников. Это объясняется огромным количеством видов, большим разнообразием форм, их широким распространением, разнообразием размеров и цветов в окраске. Коллекционирование бабочек зародилось давно. Ещё в середине XVIII века английские коллекционеры бабочек называли себя аурелианами — от лат. aureus — «золотой» (намёк на золотистую окраску куколок некоторых бабочек). В середине XIX века в интерьере викторианских домов непременно присутствует застеклённый шкаф, где наряду с раковинами морских моллюсков, окаменелостями и минералами выставлялись и засушенные бабочки[55].
Как и все энтомологические коллекции, их стоит разделять на частные и научные. Частные коллекции, в большей мере носят любительский характер и ориентируются на эстетичность, разнообразие и эффектность собранных экземпляров. Бабочки в такие коллекции могут подбираться по цветовой гамме, размерам, личным предпочтениям и материальным возможностям коллекционеров. В последнее время многие частные коллекции становятся выставочными, принося дополнительный доход их владельцам.
Научные коллекции представлены фондами музеев, университетов, научно-исследовательских институтов и так далее. Такие коллекции собираются научными работниками, которые проводят специализированные фаунистические, эволюционные и экологические исследования бабочек. Научные коллекции принято разделять на выставочные — предназначенные для всеобщего обзора — и фондовые, доступ к которым имеют лишь научные сотрудники.
В советское время сообщалось об одной из крупных частных коллекций бабочек в Харькове: «Александр Лисецкий, заведующий кафедрой позвоночных животных биологического факультета Харьковского университета, коллекционирует бабочек. В его коллекции насчитывается более 300 тысяч экземпляров, в том числе виды, впервые обнаруженные на территории СССР. Из них 15 тысяч уже систематизированы»[120].
В России самые крупные коллекции бабочек хранятся в Зоологическом музее Московского государственного университета имени М. В. Ломоносова, в Зоологическом институте Российской академии наук в Санкт-Петербурге, в Дарвинском музее в Москве и в зоомузее[121] Института систематики и экологии животных Сибирского отделения РАН в Новосибирске, в Биолого-почвенном институте Дальневосточного отделения РАН во Владивостоке. Основу фондов этих музеев, составляют частные коллекции, завещанные коллекционерами или приобретённые у их наследников. Крупнейшая коллекция, поступившая в Зоологический музей МГУ, насчитывающая около 84 000 экземпляров, была собрана знаменитым коллекционером Анатолием Васильевичем Цветаевым[11].
Коллекционированием бабочек занимались многие знаменитые люди: князь Николай Михайлович Романов, Владимир Набоков (он также опубликовал ряд научных статей по лепидоптерологии, в частности, описал новый род и ряд новых видов[122]), Михаил Булгаков, Николай Бухарин. Но пальма первенства принадлежит банкиру и финансисту Уолтеру Ротшильду, собравшему за свою жизнь самую крупную частную коллекцию в мире — около 2 миллионов 250 тысяч экземпляров бабочек[55].
Для коллекционеров ежегодно в Праге, Франкфурте-на-Майне, Париже проводятся международные выставки-продажи бабочек и других насекомых.
Самой дорогой коллекционной бабочкой за всю историю стал экземпляр мозаичного гинандроморфа вида Ornithoptera goliath, проданный одному из тайваньских дилеров в июле 2006 года по цене 28 000 долларов США[123].
Примечания
- Каабак Л. В. Бабочки-чемпионы // Наука и жизнь : журнал. — М., 2002. — Вып. 1, № 7.
- Richards O. W., Davies R. G. A General Textbook of Entomology. — 9th edn. — N. Y., NY, USA: E. P. Dutton, 1960. (англ.)
- Moucha J. Beautiful Moths. — L., UK: Drury House, 1966. (англ.)
- Герасимов А. М. Насекомые чешуекрылые. Гусеницы. — 2-е изд. — М.—Л.: Изд-во АН СССР, 1952. — Т. 1. — (Фауна СССР. Т. 56).
- Фасмер М. Этимологический словарь русского языка. — М.: Прогресс, 1964—1973. — Т. 1. — С. 100.
- Этимологический словарь славянских языков, 1974, т. 1, с. 107.
- Этимологический словарь славянских языков, 1993, т. 18, с. 137.
- Этимологический словарь славянских языков, 1994, т. 20, с. 84—85.
- Этимологический словарь славянских языков, 1992, т. 19, с. 203—205.
- Zhang Z.-Q. Phylum Athropoda // Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013) / Z.-Q. Zhang (ed.) (англ.) // Zootaxa / Z.-Q. Zhang (Chief Editor & Founder). — Auckland, New Zealand: Magnolia Press, 2013. — Vol. 3703, no. 1. — P. 17—26. — ISBN 978-1-77557-248-0 (paperback) ISBN 978-1-77557-249-7 (online edition). — ISSN 1175-5326.
- Каабак Л. В., Сочивко А. В. Бабочки мира. — М.: Аванта+, 2003. — ISBN 5-94623-008-5.
- The Lepidoptera Taxome Project Draft Proposals and Information (англ.) (недоступная ссылка). Centre for Ecology and Evolution, University College London. Дата обращения: 5 ноября 2009. Архивировано 9 ноября 2009 года.
- Кузнецов В. И., Стекольников А. А. Новые подходы к систематике чешуекрылых мировой фауны. — СПб.: Наука, 2001. — Т. 282. — С. 5. — 465 с. — (Труды ЗИН). — 655 экз.
- Каталог чешуекрылых (Lepidoptera) России. Изд. 2-е. / Под ред. С. Ю. Синева. — СПб.: Зоологический институт РАН, 2019. — 448 с. — ISBN 978-5-98092-068-5.
- Аннотированный каталог насекомых Дальнего Востока России. Том II. Чешуекрылые / Лелей А. С. (гл. ред.) и др. — Владивосток: Дальнаука, 2016. — С. 13. — 812 с. — 300 экз. — ISBN 978–5–8044–1576–2.
- Ландман В. Бабочки. Иллюстрированная энциклопедия. — М.: Лабиринт Пресс, 2002. — 272 с. — (Иллюстрированная энциклопедия). — ISBN 5-9287-0274-4.
- Frost S. W. Insect Life and Natural History. — 2nd edn. — N. Y.: Dover Publications, 1959. (англ.)
- Kons H., Jr. Chapter 32 — Largest Lepidopteran Wing Span (англ.) (недоступная ссылка). Book of Insect Records. University of Florida (17 мая 1998). Дата обращения: 20 апреля 2017. Архивировано 18 августа 2011 года.
- Определитель насекомых Дальнего Востока России. Т. V. Ручейники и чешуекрылые Ч. 1 / под общ. ред. П. А. Лера. — Владивосток: Дальнаука, 1997. — 540 с. — 500 экз. — ISBN 5-7442-0986-7.
- Review Of The Subgenus Trifurcula (Levarchama), With Two New Species (Lepidoptera: Nepticulidae). (англ.)
- Ефетов К. А., Будашкин Ю. И. Бабочки Крыма: (Высшие разноусые чешуекрылые). — Симферополь: Таврия, 1990. — 112 с. — (Охрана окружающей среды).
- Райххолф-Рим Х. Бабочки. Самый популярный справочник. — М.: Астрель, 2002.
- Корнелио М. П. Школьный атлас-определитель бабочек. — М.: Просвещение, 1986. — 256 с.
- Акимушкин И. И. Мир животных. — М.: Мысль, 1993. — Т. 3. — ISBN 5-244-00444-1.
- Стерри П. Бабочки и мотыльки. — Минск: Белфакс, 1995. — 80 с. — (Мир животных). — ISBN 985-407-011-5.
- Бродский А. К. Механика полёта насекомых и эволюция их крылового аппарата / ЛГУ им. А. А. Жданова. — Л.: Изд-во ЛГУ, 1988. — 206 с., илл.
- Smart P. The Illustrated Encyclopedia of the Butterfly World. — Transworld Publishers Ltd, 1981. — 274 p. — ISBN 0-552-98206-7. (англ.)
- Ламперт К. Атлас бабочек и гусениц. — Минск: Харвест, 2003. — 735 с. — 5000 экз. — ISBN 985-13-1664-4.
- Беккер Дж. А. Энциклопедия «Кругосвет» (недоступная ссылка). — Является дополненным и исправленным изданием в переводе на русский язык «Энциклопедии Кольера» («Collier’s Encyclopedia»), выходившей в США в 1952—1998 годах. Дата обращения: 16 августа 2009. Архивировано 13 августа 2009 года.
- Лапшин Д. Н. Эхолокационная система бабочек. — М.: Наука, 2005. — 179 с.
- Захваткин Ю. А. Эмбриология насекомых. — М.: Высшая школа, 1975.
- Wilts B. D., Matsushita A., Arikawa K., Stavenga D. G. Spectrally tuned structural and pigmentary coloration of birdwing butterfly wing scales (англ.) // Journal of the Royal Society Interface : журнал. — L.: The Royal Society, 2015. — Vol. 12, no. 111. — P. 20150717. — ISSN 1742-5662. — doi:10.1098/rsif.2015.0717.
- Яхонтов А. А. Дневные бабочки пособие для определения и биологического изучения Lepidoptera, Rhopalocera европейской части СССР. — К.: Радянська школа, 1939. — 92 с.
- Перевозчиков А. П. Влияние факторов внешней среды на развитие животных.
- Тыкач Я. Маленький атлас бабочек. — Прага: Госиздат, 1959. — 98 с.
- Моргун Д. В., Довгайло К. Е., Рубин Н. И., Солодовников И. А., Плющ И. Г. Дневные бабочки (Hesperioidea и Papilionoidea, Lepidoptera) Восточной Европы. CD определитель, база данных и пакет программ «Lysandra». — Мн., К., М., 2005.
- Эд А., Виар М. Бабочки мира. — Изд. 2-е, стереотипное. — Интербук-бизнес, 2001. — 193 с. — ISBN 5-89164-090-2.
- Мурзин В. Безмолвный язык любви // Наука и жизнь : журнал. — М., 1998. — № 10. Архивировано 15 апреля 2010 года.
- Эволюция жизни. Факторы эволюции (недоступная ссылка). Дата обращения: 14 октября 2009. Архивировано 25 мая 2009 года.
- Формы естественного отбора (недоступная ссылка — история ). Дата обращения: 14 октября 2009. (недоступная ссылка) Проверено Июль 2018.
- Majerus M. E. N. Industrial Melanism in the Peppered Moth, Biston betularia: An Excellent Teaching Example of Darwinian Evolution in Action (англ.) (недоступная ссылка) (6 декабря 2008). Дата обращения: 14 декабря 2017. Архивировано 15 декабря 2017 года.
- Beck J., Mühlenberg E., Fiedler K. Mud-puddling behavior in tropical butterflies: In search of proteins or minerals? // Oecologia. — 1999. — № 119. — P. 140—148.
- Кононенко В. С. Сем. Noctuidae — Совки, или ночницы. Введение // Определитель насекомых Дальнего Востока России. Т. V. Ручейники и чешуекрылые. Ч. 4. / под общ. ред. П. А. Лера. — Владивосток: Дальнаука, 2003. — С. 11. — 688 с. — 500 экз. — ISBN 5-8044-0343-5.
- Hilgartner R., Raoilison M., Büttiker W., Lees D. C., Krenn H. W. Malagasy birds as hosts for eye-frequenting moths (англ.) // Biology Letters. — Royal Society Publishing, 2007. — Vol. 3. — P. 117—120. (англ.)
- Heinrich B The Hot-Blooded Insects: Strategies and Mechanisms of Thermoregulation (англ.). — Berlin, Heidelberg: Springer-Verlag, 1993. — P. 601. — ISBN 978-3-662-10342-5. — doi:10.1007/978-3-662-10340-1. (англ.) (Дата обращения: 25 февраля 2017) Архивированная копия. Дата обращения: 14 декабря 2017. Архивировано 17 апреля 2019 года.
- Heinrich B., ed. (1981), Insect thermoregulation, N. Y.: John Wiley & Sons, Inc., с. 328, ISBN 0-471-05144-6
- Traut W., Sahara K., Marec F. Sex chromosomes and sex determination in Lepidoptera (англ.) // Sexual Development. — 2007. — Vol. 1, no. 6. — P. 332—346. — ISSN 1661-5425. — doi:10.1159/000111765. (Дата обращения: 4 апреля 2013) Архивированная копия. Дата обращения: 1 июля 2019. Архивировано 17 апреля 2019 года.
- Cockayne E. A. The genetics of sex in Lepidoptera (англ.) // Biological Reviews. — 1938. — April (vol. 13, no. 2). — P. 107—132. — ISSN 1464-7931. — doi:10.1111/j.1469-185X.1938.tb00509.x. (Дата обращения: 26 мая 2012) Архивированная копия. Дата обращения: 1 июля 2019. Архивировано 18 апреля 2019 года.
- Clarke C., Ford E. B. Intersexuality in Lymantria dispar (L.): a further reassessment // Proc. R. Soc. Lond. B. — 1982. — Vol. 214. — No. 1195. — P. 285—288. (англ.)
- Narita S., Kageyama D., Nomura M., Fukatsu T. Unexpected mechanism of symbiont-induced reversal of insect sex: feminizing Wolbachia continuously acts on the butterfly Eurema hecabe during larval development // Applied and Environmental Microbiology. — 2007. — Vol. 73. — No. 13. — P. 4332—4341. (англ.)
- Kageyama D., Traut W. Opposite sex-specific effects of Wolbachia and interference with the sex determination of its host Ostrinia scapulalis // Proc. R. Soc. Lond. B. — 2004. — Vol. 271. — P. 251—258. (англ.)
- Monteiro A., Glaser G., Stockslager S., Glansdorp N., Ramos D. Comparative insights into questions of lepidopteran wing pattern homology (англ.) // BMC Developmental Biology. — 2006. — Vol. 6, no. 1. — P. 52. — ISSN 1471-213X. — doi:10.1186/1471-213X-6-52. (Дата обращения: 4 декабря 2009) Архивированная копия. Дата обращения: 4 декабря 2009. Архивировано 26 ноября 2012 года.
- d’Abrera B. World Butterflies. — Hill House Publishers, 2006. — 272 p. — ISBN 0-947352-46-5. (англ.)
- Leraut P. Moths of Europe: Saturnids, Lasiocampids, Hawkmoths, Tiger Moths. — 2006. — Vol. 1. — 387 p. — (Moths of Europe). — ISBN 2-913688-07-1. (англ.)
- Рассел Ш. Э. Роман с бабочками. Как человек влюбился в насекомое. — М.: Колибри, 2005.
- Глущенко Ю. Н., Тарасов А. А., Чистяков Ю. А. Биология эпикопеи изменчивой (Epicopeia mencia Moore) (Lepidoptera, Epicopeidae) в Приморье и необходимые меры по её охране // Чтения памяти Алексея Ивановича Куренцова. — Владивосток: ДВО РАН, 1992. — Вып. I—II. — С. 102—116.
- Савчук В. В. Атлас бабочек и гусениц Крыма. — Симферополь: Бизнес—Информ, 2013. — 296 с. — ISBN 978-966-648-331-0.
- Горбунов О. Г. Биоразнообразие, зоогеография и история фауны бабочек-стеклянниц (Lepidoptera, Sesiidae) России и сопредельных территорий // Известия АН. Серия биологическая. — 1998. — Т. 4. — С. 453—457.
- Laštuvka Z., Laštuvka A. The Sesiidae of Europe. — Stenstrup: Apollo Books, 2001. — P. 245.
- Райков Б. Е., Римский-Корсаков М. Н. Зоологические экскурсии. — 7-е изд. — М.: Топикал, 1994. — С. 640. — 30 000 экз.
- Котт К. Приспособительная окраска животных = Adaptive Coloration in Animals. — М.: Изд-во ин. лит-ры, 1950. Архивированная копия (недоступная ссылка). Дата обращения: 19 августа 2009. Архивировано 11 декабря 2015 года.
- Morewood W. D., Ring R. A. Revision of the life history of the High Arctic moth Gynaephora groenlandica (Wocke) (Lepidoptera: Lymantriidae) (англ.) // Canadian Journal of Zoology. — 1998. — July (vol. 76, no. 7). — P. 1371—1381. — ISSN 0008-4301. — doi:10.1139/z98-085.
- Черешнев И. А., Андреев А. В., Берман Д. И., Докучаев Н. Е., Кашин В. А. Животные // Красная книга Чукотского автономного округа / Отв. ред. И. А. Черешнев. — Дикий Север, 2008. — Т. 1. — 235 с.
- Kluge N. J. Larval/pupal leg transformation and a new diagnosis for the taxon Metabola Burmeister 1832=Oligoneoptera Martynov 1923 // Russian Entomol. J. — 2004. — Vol. 13, no. 4. — P. 189—229. PDF (англ.). [Русский перевод: Трансформация ног при превращении личинки в куколку и новый диагноз таксона Metabola.]
- Коршунов, 2002, с. 12
- Тыщенко В. П. Физиология насекомых. — М.: Высшая школа, 1988. — 299 с.
- Tabashnik B.E. Population structure of pierid butterflies. III. Pest population of Colias philodice eryphile // Oecologia. — 1980. — № 47. — P. 175—183.
- Крыжановский О. Л. Состав и распространение энтомофаун земного шара. — М.: Товарищество научных изданий КМК, 2002. — 237 с. — ISBN 5-87317-116-5 (0).
- Tuzov V. K. Guide to the Butterflies of Russia and Adjacent Territories: Hesperiidae, Papilionidae, Pieridae, Satyridae. — Sofia—Moscow, 1997. — Vol. 1. — 480 p., ill. (англ.)
- Mani M. S. Ecology and Biogeography of High Altitude Insects (англ.). — Springer, 1968. — P. 530. — (Volume 4 of Series entomologica). — ISBN 978-90-6193-114-0. (англ.)
- Mølgaard P., Morewood D. ITEX insect: Gynaephora groenlandica/G. rossii. ITEX Manual / U. Molau, P. Mølgaard (ed.). — Copenhagen, Denmark: Danish Polar Center, 1996. — P. 34—36. PDF (недоступная ссылка) (англ.)
- Тинберген Н. Поведение животных. — М.: Мир, 1978. — 192 с., илл. с.
- Williams C. B. The Migration of Butterflies. — Edinburgh, UK: Oliver & Boyd, 1930. (англ.)
- Senthilmurugan B.. Mukurthi National Park: A migratory route for butterflies (англ.) // Journal of the Bombay Natural History Society : journal. — Bombay Natural History Society, 2005. — Vol. 102, no. 2. — P. 241—242. (англ.)
- Nature Australia : журнал. — 2003. — Вып. 27, № 10. (англ.)
- Pierce et al. Annual Review of Entomology. — 2002. — Vol. 47. — P. 47, 733—771. (англ.)
- Fiedler K., Holldobler B., Seufert P. Butterflies and ants: «The communicative domain» // Experientia. — 1996. — Vol. 52, no. 1. — P. 14—24. (англ.)
- Kistner D. H. The Social Insects' Bestiary // In: Social Insects. Vol 3. (Eds. Henry Hermann) (англ.). — New York, London: Academic Press, 1982. — P. 1—244 (Lepidoptera: 162—177). — 459 p. — ISBN 0-12-342203-5.
- Gilmore D. P., Da Costa C. P., Duarte D. P. F. Sloth biology: an update on their physiological ecology, behavior and role as vectors of arthropods and arboviruses (англ.) // Brazilian Journal of Medical and Biological Research. — 2001. — January (vol. 34, no. 1). — P. 9—25. — ISSN 0100-879X. — doi:10.1590/S0100-879X2001000100002. — PMID 11151024.
- Некрутенко Ю. П., Чиколовець В. Денні метелики України. — К.: Видавництво Раєвського, 2005. — 232 с. — (Природа України). — ISBN 966-7016-17-X.
- Защита растений на микробиологическом уровне. Вирусные болезни лесных насекомых (недоступная ссылка). Дата обращения: 7 сентября 2012. Архивировано 13 декабря 2013 года.
- Dodd F. P. Some remarkable ant-friend Lepidoptera (недоступная ссылка). Дата обращения: 20 августа 2016. Архивировано 18 августа 2016 года. // Transactions of the Entomological Society of London. — 1911. — Vol. 577—589, plate XLVIII. (англ.)
- Rau, Phil. Observations on certain lepidopterous and hymenopterous parasites of Polistes wasps (англ.) // Annals of the Entomological Society of America. — 1941. — June (vol. 34, no. 2). — P. 355—366.
- Rau P. Observations on certain lepidopterous and hymenopterous parasites of Polistes wasps (англ.) // Annals of the Entomological Society of America : journal. — 1941. — Vol. 34, no. 12. — P. 355—366. (англ.) (недоступная ссылка) Проверено 22 апреля 2018.
- Белокобыльский С. А., Тобиас В. И. Сем. Braconidae — Бракониды. 9. Подсем. Alysiinae. Группа родов, близких к Aspilota // Определитель насекомых Дальнего Востока России. Сетчатокрылообразные, скорпионницы, перепончатокрылые. — Владивосток: Дальнаука, 2007. — Т. 4, ч. 5. — С. 9—133.
- Тобиас В. И. [ред. и автор или первый автор]. Отряд Hymenoptera — Перепончатокрылые. Семейство Braconidae — Бракониды // Определитель насекомых европейской части СССР. — Л.: Наука, 1986. — Т. 3. — Ч. 4. — 500 с.; Ч. 5. — С. 1—231, 284—307; Сем. Aphidiidae — Афидииды // Ч. 5. — С. 232—283, 308.
- Лелей А. С. Гл. 5 // Определитель насекомых Дальнего Востока России. — Владивосток: Дальнаука, 2007. — Т. IV. — С. 255. (Дата обращения: 27 августа 2009)
- Каспарян Д. Р. Отряд Hymenoptera — Перепончатокрылые. Семейство Ichneumonidae — Ихневмониды, Введение // Определитель насекомых европейской части СССР. — Л.: Наука, 1981. — Т. 3. — Ч. 3. — 688 с.
- Савковский П. П. Атлас вредителей плодовых и ягодных культур. — 5-е изд., доп. и перераб. — К.: Урожай, 1990. — 96 с., цв. табл., илл. с.
- Grimaldi D., Engel M. S. Evolution of the Insects. — Cambridge University Press, 2005. — 755 p. — (Cambridge Evolution Series). — ISBN 0-521-82149-5, ISBN 978-0-521-82149-0. (англ.)
- Whalley P. E. S. The systematics and palaeogeography of the Lower Jurassic insects of Dorset, England // Bulletin of the British Museum of Natural History (Geology). — 1985. — Vol. 39, no. 3. — P. 107—189. (англ.)
- Tree of Life (англ.) (недоступная ссылка) (2007). Дата обращения: 29 июня 2007. Архивировано 22 июня 2007 года.
- Родендорф Б. Б., Расницын А. П. Историческое развитие класса насекомых. — М.: Наука, 1980. — С. 181—184. — 256 с.
- Eid A., Viard M. Les Papillons du Monde. — Paris, France: Les Editions Hatier, 1996. (фр.)
- Korb S. K. A Catalogue of Butterflies of the ex-USSR, with Remarks on Systematics and Nomenclature. — Nizhny Novgorod: Nizhegorodskaya Radiolaboratoriya Press, 2005. — 156 p. Архивированная копия (недоступная ссылка). Дата обращения: 30 августа 2009. Архивировано 25 октября 2009 года. (англ.)
- Кочетова Н. И. Они должны жить. Бабочки. — М.: Агропромиздат, 1990. — 64 с.
- Lepidoptera: информация на сайте Красной книги МСОП (англ.)
- Gärdenfors U., Stattersfield A. J. 1996 IUCN Red List of Threatened Animals. — 1996. — P. 47—48. — 455 p. — ISBN 2-8317-0335-2.
- Никитский Н. Б., Свиридов А. В. Насекомые Красной книги СССР. — М.: Педагогика, 1987. — С. 9. — 176 с. — (Береги природу). — 180 000 экз.
- Тихонов А. Красная книга России. Животные и растения. — М.: РОСМЭН, 2002. — 414 с. — 10 000 экз. — ISBN 5-353-00500-7.
- Россияне, пытавшиеся вывезти насекомых из Перу, освобождены (недоступная ссылка). Полит.ру (12 декабря 2008). Дата обращения: 1 декабря 2009. Архивировано 14 мая 2013 года.
- Приложения CITES — Appendices I, II and III. Valid from 27 April 2011 (англ.). Дата обращения: 18 марта 2010. Архивировано 28 сентября 2006 года.
- Хопкинс Дж. Экстремальная кухня. Причудливые и удивительные блюда, которые едят люди. — М.: ФАИР-ПРЕСС, 2006. — 336 с. — ISBN 5-8183-1032-9. Архивированная копия (недоступная ссылка). Дата обращения: 7 сентября 2012. Архивировано 18 июля 2013 года.
- Cherry R. Use of insects by Australian aborigines (англ.) (недоступная ссылка). Cultural Entomology Digest — Insect Articles. Дата обращения: 7 сентября 2012. Архивировано 16 октября 2012 года.
- Мариковский П. И. Тайны мира насекомых. — Алма-Ата: Кайнар, 1966. — 164 с.
- Энтомодизайн (недоступная ссылка). Дата обращения: 5 декабря 2009. Архивировано 24 мая 2009 года.
- Хисматуллина Н. З. Апитерапия. — Пермь: Мобиле, 2005. — С. 71—76. — 296 с. — 10 000 экз. — ISBN 5-88187-263-0.
- Food and Agricultural Organization of United Nations: Economic And Social Department: The Statistical Devision (англ.) (недоступная ссылка). Дата обращения: 12 декабря 2009. Архивировано 24 сентября 2009 года.
- Jolly M. S., Sen S. K., Sonwalkar T. N. Prasad G. S. Non-mulberry Silks // Serv. Bull. — Rome, Italy: Food & Agriculture Organisation, United Nations, 1979. — No. 29. — P. xvii + 178 p. (англ.)
- Annecke D.P., Burger W.A., Coetzee H. Pest status of Cactoblastis cactorum (Berg) (Lepidoptera: Phycitidae) and Dactylopius opuntiae (Cockerell) (Coccoidea: Dactylopiidae) in spineless Opuntia plantations in South Africa // Journal of the Entomological Society of South Africa. — 1976. (англ.)
- Floyd J. Pest Alert: Cactus moth Cactoblastis cactorum (англ.) (недоступная ссылка). APHIS (2006). Дата обращения: 18 февраля 2007. Архивировано 26 ноября 2012 года.
- Алексеев В. Бабочки в мифах и легендах. — Дрофа, 2006. — 192 с. — (Исторический альбом). — ISBN 5-358-00249-9.
- Гура А. В. Символика животных в славянской народной традиции. — М., 1997. — С. 486—492.
- Этимология и история слов русского языка (недоступная ссылка). Дата обращения: 29 октября 2009. Архивировано 16 марта 2010 года.
- Harrison R. (ed.); van Gogh-Bonger J. (translator): Vincent van Gogh. Letter to Theo van Gogh. Written 22 May 1889 in Saint-Rémy (англ.) (недоступная ссылка). Van Gogh Letters. WebExhibits (funded in part by U.S. Department of Education, Fund for the Improvement of Postsecondary Education); Institute for Dynamic Educational Advancement (IDEA) (2011). Дата обращения: 18 апреля 2011. Архивировано 19 июля 2011 года.
- Emperor Moth, 1889 (англ.) (недоступная ссылка). Permanent Collection. Van Gogh Museum (2005—2011). Дата обращения: 15 апреля 2011. Архивировано 28 сентября 2011 года.
- Геральдика мира (недоступная ссылка). Дата обращения: 10 октября 2009. Архивировано 27 октября 2009 года.
- Гербы городов Российской Федерации (недоступная ссылка). Дата обращения: 10 октября 2009. Архивировано 25 октября 2009 года.
- О. Петрова. Все о татуировке. Татуировка от А до Я. — Владис, 2006. — 288 с. — 8000 экз. — ISBN 5-9567-0248-6.
- Мозаика // Филателия СССР. — 1975. — № 12. — С. 63.
- Сибирский Зоологический музей ИСиЭЖ СО РАН (недоступная ссылка). Дата обращения: 15 сентября 2012. Архивировано 19 ноября 2012 года.
- Nabokov V. A third species of Echinargus Nabokov (Lycaenidae, Lepidoptera) (англ.) // Psyche: A Journal of Entomology. — 1945. — 1 January (vol. 52, no. 3—4). — P. 193—193. — ISSN 0033-2615. — doi:10.1155/1945/47328. (Дата обращения: 26 мая 2012) Архивированная копия. Дата обращения: 26 мая 2012. Архивировано 25 января 2013 года.
- The World of Birdwing Butterflies (англ.) (недоступная ссылка). Дата обращения: 14 октября 2009. Архивировано 7 февраля 2009 года.
Литература для дополнительного чтения
- Бабочки // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- Коршунов Ю. П. Определители по флоре и фауне России // Булавоусые чешуекрылые Северной Азии. Выпуск 4. — М.: Товарищество научных изданий КМК, 2002. — 424 с. — ISBN 5-87317-115-7.
- Львовский А. Л., Моргун Д. В. Булавоусые чешуекрылые Восточной Европы. — М.: Товарищество науч. изд. КМК, 2007. — 442, [1] с., [4] л. ил.: ил. — (Определители по флоре и фауне России; вып. 8).
- Татаринов А. Г., Долгин М. М. Видовое разнообразие булавоусых чешуекрылых на европейском Северо-Востоке России. — М.: Наука, 2001. — 244 с. — ISBN 5-02-026164-5.
- Уэйли П. Бабочки. Eyewitness Guides — Butterfly and Moth. — Лондон, Нью-Йорк, Штутгарт, М.: Дорлинг Киндерсли (в сотрудничестве с Музеем естественной истории — Лондон), Слово, 2000. — ISBN 0-86318-319-0 (Великобритания); ISBN 5-85050-550-4 (Россия).
- Этимологический словарь славянских языков. — М.: Наука.
- Butterflies: Ecology and Evolution Taking Flight / Ed. by Carol L. Boggs, Ward B. Watt, Paul R. Ehrlich. — 2nd edn. — Chicago, L.: University of Chicago Press, 2003. — 736 p. — ISBN 0-226-06318-6. (англ.)
- Tuzov V. K. Guide to the Butterflies of Russia and Adjacent Territories: Libytheidae, Danaidae, Nymphalidae, Riodinidae, Lycaenidae. — Sofia—Moscow, 1997. — Vol. 2. — 480 p., ill. (англ.)
Ссылки
- Бабочки Европы (недоступная ссылка). Дата обращения: 8 октября 2017. Архивировано 2 октября 2017 года. (англ.) (нем.) (польск.) (рус.)
- Галерея лепидоптерологов России и сопредельных стран (недоступная ссылка). Дата обращения: 28 января 2011. Архивировано 19 августа 2011 года.
- Colour Atlas of the Siberian Lepidoptera (англ.) (недоступная ссылка). Дата обращения: 15 июля 2011. Архивировано 16 июня 2012 года.
- Бабочки Крыма — фотогалерея (недоступная ссылка). Дата обращения: 13 декабря 2008. Архивировано 1 декабря 2008 года.
- Бабочки Кавказа и юга России (недоступная ссылка). Дата обращения: 2 июля 2012. Архивировано 6 июля 2012 года.
- Mapování a ochrana motýlů České republiky — Бабочки Чехии (чешск.).
- Чешуекрылые (Macrolepidoptera) Палеарктики: наблюдения, поиск и содержание гусениц (недоступная ссылка). Дата обращения: 29 июля 2018. Архивировано 14 июля 2018 года.
- The Global Lepidoptera Names Index (англ.) (недоступная ссылка). Дата обращения: 5 сентября 2013. Архивировано 28 сентября 2013 года.
| Таксономия | |
|---|---|
| В библиографических каталогах |
