Крыло насекомых
Крылья насекомых — придатки экзоскелета насекомых, которые используются ими для полёта. Две пары крыльев расположены на втором и третьем сегментах груди (мезотораксе и метатораксе). Пластинка крыла армирована жилками. Крыло состоит из двух стенок — верхней и нижней. Каждая стенка образована слоем гиподермы, снаружи покрытой более-менее развитой кутикулой. Между стенками находится узкая щель (часть миксоцеля — смешанной полости тела), заполненная гемолимфой. Крыло имеет систему хитиновых трубочек-жилок. Жилки выполняют опорную функцию, в них содержится гемолимфа, проходят трахеи и нервные пучки.

Филогения
Существует три основные гипотезы возникновения крыльев насекомых:
- Согласно первой, крылья возникли из паранотальних лопастей грудных тергитов.
- Согласно второй, они представляют собой видоизмененные абдоминальные жабры, сходные с жабрами личинок современных подёнок.
- Согласно третей, крылья возникли из тазобедренной области ног.
Онтогенез
В начале своего онтогенеза крыло является сплюснутым в дорсовентральном направлении мешком, внутренняя полость которого является продолжением полости тела. Стенки, являясь продолжением стенок тела, состоят из гиподермальных клеток, под которыми располагается базальная мембрана. Первоначально полость крыла заполнена гемолимфой, позднее в неё врастают ответвления трахей и нервы.
Позднее делящиеся гиподермальные клетки образуют складки крыла, одновременно они образуют кутикулу, первоначально мягкую. Окончательное развитие крыла происходит после вылупления имаго из куколки. Под воздействием нагнетаемой гемолимфы, крыло растягивается приобретая окончательные размеры и форму, при этом гиподермальные клетки превращаются из цилиндрических в плоские. Происходит затвердение кутикулы и сближение дорсальной и вентральной поверхностей. Позже плазма клеток дегенерирует, и обе хитиновые пластинки соединяются (например, таракан)
Однако в местах залегания трахей и нервов процесс происходит по другому пути. Гиподерма и хитин образуют по ходу трахей выпуклости. Выпуклость спинной пластинки расположена напротив брюшной, и вместе они образуют трубку, полость которой заполнена гемолимфой, а внутри неё проходят трахея и нервы. Таким образом формируются жилки.
Описание
Крылья насекомых (alae) представляют собой пластинчатые выросты покровов, связанные с комплексом обслуживающих их мышц и преобразованных склеритов груди. В типичном случае их две пары: передняя, прикреплённая к среднегруди, и задняя — на заднегруди. Крылья состоят из тонкой крыловой пластинки (membrana), укреплённой на твёрдых жилках (venae), играющих роль прочной основы крыла. Жилки делятся на продольные и поперечные, образующие у более низкоорганизованных групп густую сеть и многочисленные ячейки (cellulae). Поперечные жилки у высших форм сокращаются в числе и иногда исчезают совсем, так как главную поддерживающую роль несут продольные, попеременно выпуклые и вогнутые жилки. Поэтому продольные жилки, хотя и претерпели у высших групп значительную эволюцию, всё же сохранили свою основу.
Наличие крыльев свойственно большинству насекомых из подкласса Pterygota, и лишь некоторые из них вторично утратили крылья в ходе регрессивной эволюции. У бескрылых насекомых Apterygota крылья вообще отсутствуют. Полностью функциональные крылья появляются в стадии развития — имаго. Одним исключением является отряд подёнок (Ephemeroptera) у которых крылья появляются в предпоследней стадии развития (субимаго) и «отпадают» у имаго. Низшие насекомые и личинки высших насекомых характеризуются первичным отсутствием крыльев.
Крылья могут присутствовать только у представителей одного пола — чаще у самца как, например, у ос-немок (Mutillidae) и веерокрылых (Strepsiptera). У рабочих особей некоторых социальных насекомых, таких как муравьи и термиты, крылья также отсутствуют. Реже встречается обратный пример, когда крылья имеются только у самки — например у ос из семейства Agaonidae.
Иногда крылья появляются лишь на отдельной стадии жизненного цикла, например, на стадии расселения у тлей. Кроме присутствия или отсутствия крыльев, разные морфы могут также отличаться по структуре и окраской крыльев, как, например, тли, мигрирующие стадии саранчи и полиморфные бабочки.
В состоянии покоя крылья могут быть распластаны, или сложены несколько раз (чаще всего складываются лишь задние крылья, но, например, у веспоидных ос составляется передняя пара).
Крылья, прикрепленные ко второму грудному сегменту, принято называть передней или верхней парой; прикреплённые к третьему грудному сегменту — задней или нижней парой.
У каждого крыла принято выделять 3 края:
- передний — идёт от основания крыла к его вершине
- внутренний — начинается от основания крыла, не достигает вершины и находится напротив переднего края
- задний (внешний) — проходит от вершины до свободного края внутреннего края
Угол между передним и внешним краем принято называть передним или вершинным углом, угол между внешним и внутренним краем — внутренним углом, а угол между задним крылом и телом — задним углом.
Иногда у костального края крыла, у его вершины, имеется плотный тёмноокрашенный участок, называемый птеростигма (крыловой глазок).
Жилкование
В местах залегания трахей и нервов на крыльях насекомых образуются трубчатые утолщения — жилки, являющиеся каркасом крыла. Жилкование крыльев играет важную роль в систематике на разных уровнях, от вида до подотряда.
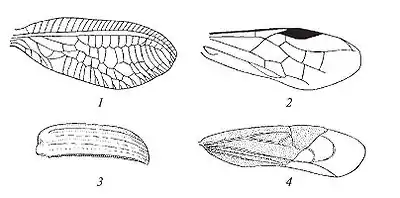
Выделяют два основных типа жилкования: сетчатое и перепончатое[1]. Сетчатое жилкование — густое, мелкоячеистое жилкование, образованное продольными и мелкими поперечными жилками, образующими (более 20) замкнутые ячейки. Данный тип жилкования встречается у отрядов Стрекозы, Прямокрылые, Сетчатокрылые и других. Перепончатое жилкование — негустое, с небольшим числом или отсутствием поперечных жилок, ячейки крупные, немногочисленные. Данный тип жилкования развит у большинства отрядов насекомых (Чешуекрылые, Перепончатокрылые, Двукрылые, Жесткокрылые и др.). Жилкование передних и задних крыльев насекомых всегда однотипно.
Названия жилкам и ячейкам дают согласно системе Комстока-Нидема (1898).
На крыльях имеются следующие продольные жилки:
- костальная (costa, обозначается С) — неветвящаяся, отходит от плечевой пластинки, как правило образует утолщенный передний край крыла.
- субкостальная (subcostata, обозначается Sc) — неветвящаяся, отходит от плечевой пластинки, располагается непосредственно за костальной, всегда располагаясь на дне желобка между костальной и радиальной жилками.
- радиальная (radius, обозначается R) — неветвящаяся, толстая, связана у основания с медиальной пластинкой.
- медиальная (media, обозначается М) — отходит от медиальной пластинки, дихотомически разделяется на переднюю и заднюю ветви — MA и MP — каждая из которых, в свою очередь, делятся ещё на две ветви:
- кубитальная (cubitus, обозначается Cu) — отходит от кубитальной пластинки, двухветвистая, сильно выступающая жилка, проходит вдоль гребня складки. Кубитальных жилок может быть две — передняя (CuA) и задняя (CuP)
- анальная (analis, обозначается A) — представляет собой пучок жилок с общим основанием, непосредственно связанное с третьим аксиллярным склеритом.
Эти жилки образуют основную схему. В процессе эволюции жилкование крыльев подверглось изменениям — упрощению или усложнению. Так могут появляться дополнительные ветви на субкостальной, радиальной и других жилках.
Главная задняя ветвь радиальной жилки часто называется радиальным сектором (RS). Также на задних крыльях, могут проходить югальные жилки (Ju), располагающиеся позади анальных и входящие в состав складывающейся задней части крыла.
Иногда у костального края крыла, у его вершины, имеется плотный тёмноокрашенный участок, называемый птеростигма (крыловой глазок).
От продольных жилок крыла отходят боковые ветви, образующие сеть жилок, разделяющих крыло на участки, называемые ячейками. В первую очередь выделяют срединную ячейку (cellula media), располагающуюся от корня до середины крыла. Обычно она объединяет 6-8 продольных жилок, идущих раздельно к краю крыла. Также от корня крыла отходят ещё две или три продольные жилки, находящиеся между передним и внешним краем и серединной ячейкой.
Близко к переднему краю передних крыльев проходит переднекрайняя жилка (I), за которой располагается срединная ячейка (MZ), ограниченная спереди субкостальной жилкой (Sс). Последняя даёт по направлению к переднему краю крыла пять ветвей (II, 1-5), четвёртая и пятая из которых имеют общий ствол. Кнаружи (по направлению к внешнему краю) серединная ячейка ограничена верхней, средней и нижней дискоидальными жилками (ODC, MDC, UDC), от которых отходят верхняя и нижняя радиальные жилки (III,1-2) к внешнему краю. Задняя граница серединной ячейки образует серединную жилку (М), от которой отходят три ветви к задней части внешнего края передних крыльев (V, 1-3). Позади лежит подсерединная жилка (SM)- заканчивающаяся между внешним и задним краем, от неё отходит одноимённая ветвь (V, 1). Между третьей и четвёртой субкостальной ветвью находится передняя вилообразная ячейка (VGZ), а между четвёртой и пятой — главная вилообразная ячейка (GZ). На боковом крае сзади располагаются семь краевых ячеек.
Заднее крыло также имеет костальную жилку (I), от которой отходит по направлению к внутреннему краю крыла маленькая ветвь — предкостальная жилка (I, 1). Далее идет соединенная с костальной жилкой маленькой ветвью неразветвленная субкостальная жилка. Как и на переднем крыле, имеются верхняя, средняя и нижняя дискоидальные жилки (ODC, MDC, UDC) от которых отходят верхняя и нижняя радиальные жилки. От серединной жилки направляются, три ветви (IV, 1,2,3); третья из них заканчивается в хвостовом конце заднего крыла. Подсерединная жилка (SM) не имеет ветвей. Из ячеек в заднем крыле выделяют серединную и девять краевых.
Описанное жилкование свойственно для высших отрядов насекомых. Ряд примитивных форм, такие как прямокрылые, подёнки, стрекозы, сетчатокрылые, имеют архедиктион — продольные стволы жилок, соединенные множеством поперечных. Считается, что данные поперечные жилки имеют вторичное происхождение, так как лишены трахейной основы. Поперечные жилки высших насекомых также вероятно являются остатками архедиктиона.
Различают 3 типа жилкования заднего крыла у жесткокрылых: карабоидный, стафилиноидный и кантаридоидный. Первый тип характерен примитивным семействам. Стафилиноидный тип характеризуется полным отсутствием поперечных жилок.
В исходном состоянии различают следующие продольные жилки: костальная (costa или C); субкостальная (subcosta, Sc); радиальная (radius, R), снабжённая сзади ветвью — радиальным сектором (radii sector, RS); срединная (media, M), иногда делится на переднюю (MA) и заднюю (MP) ветви; кубитальная (cubitus, Cu), в числе двух, нередко сближенных жилок — передней (CuA) и задней (CuP); анальные (analis, A) в числе 1—4. В некоторых случаях на задних крыльях позади анальных располагается ещё серия югальных (jugalis, Ju) жилок, входящих вместе с анальными в состав складывающегося веера (vannus) крыла. Между продольными жилками располагается серия полей (campo), называемых по пробегающей спереди жилке. Более крупными подразделениями являются области (regio) крыла: главная — спереди от анальных жилок, анальная — между анальными жилками, югальная — между югальными.
Этот исходный план жилкования подвергается существенным модификациям путём образования ряда ветвей или выпадения отдельных жилок, их перемещения, слияния и т. д. Наиболее существенно в эволюции крыльев появление у новокрылых насекомых югальной области, обеспечившей особое складывание крыльев вдоль тела. Однако, в процессе дальнейшей эволюции югальная область и способность складывать крылья вдоль тела неоднократно утрачивались в разных группах. Также отмечается процесс костализации крыла, то есть смещения всех продольных жилок к переднему краю крыла, укреплявшего механическую основу и повышавшего аэродинамические характеристики. Наивысшей степени костализация достигла у перепончатокрылых и двукрылых. С совершенствованием полёта от относительной независимости крылья насекомых «переходили» к сцеплению друг с другом различными способами, функционируя как единый орган с перемещением основной нагрузки на первую пару. С дальнейшей эволюцией задняя пара сокращается в размерах, а затем и утрачивается. Возникает высший этап полёта насекомых — морфологическая двукрылость.
Модификации жилкования
У наиболее архаичных форм отряда Mecoptera из пермских отложений описано округлое, с широким основанием, почти симметричное крыло. Жилки которого располагаются веерообразно, радиальная жилка проходит почти по срединной линии, ветвления немногочисленные, поперечных жилок мало. Данный тип жилкования считается наиболее примитивным. В дальнейшем, в ходе процесса эволюции происходили множественные его модификации[2].
Первой из них является развитие архедиктиона — продольных стволов жилок, соединенных множеством поперечных — который был широко распространен среди ископаемых насекомых. В настоящее время архедиктион сохранился только у некоторых прямокрылых, сетчатокрылых, стрекоз, подёнок, тараканов и, возможно, в надкрыльях некоторых жесткокрылых.
Вторая модификация — многократное разветвление продольных жилок, характерное для современных тараканов.
Третья модификация — костализация (костальная концентрация жилок). При этом радиальные жилки тесно располагаются ближе к костальному краю крыла, в результате чего происходит расширение медиальной ячейки (чешуекрылые, двукрылые). Функциональное значение такой модификации заключается в укреплении костального края крыла для полёта. Также костализация приводит к несимметричности формы крыла. Костальная концентрация сильно варьирует в пределах одного и того же отряда.
Четвёртая модификация — дифференциация ячеек в результате смещения жилок и их вторичных сращений (высшие двукрылые, перепончатокрылые).
Сочленение крыла с телом
Сочленение крыла является одним из наиболее совершенных механизмов в теле насекомых. Задача сочленения: обеспечить крылу максимальную быстроту взмаха и ограничить его движение точными пределами. По способу прикрепления к телу различают древнекрылых и новокрылых насекомых. Древнекрылые движутся в одной плоскости. Новокрылые при полёте вращаются вокруг оси (пчёлы, шмели).
Крыловая мускулатура
Продольные спинные мышцы. Мышца соединяет фрагмы и развита у летающих форм. Сокращения данной мышцы приводят к выгибанию тергита вверх, благодаря чему гребная часть крыла опускается. В переднегруди её передний конец крепится к постмаксиллярной складке и участвует также в движении головы. Вторая продольная мышца располагается сбоку от первой и проходит в каждом сегменте от задней фрагмы к скутуму. Основная функция — вспомогательное опускание крыла.
Плейральные мышцы. Целая группа мышц. Наибольшее значение имеют три из них. Первая и вторая лежащие в пределах эпистерны и сходящиеся кверху к базаларной пластинке. Данная пластинка лежит в мембране, соединяющей крыло с плейритом. При сокращении этих мышц передний край крыла опускается и изменяется наклон крыла. Также данные мышцы служат для расправления крыла и двигают коксу и таким образом, обладая тройной функцией. Третья мышца является антагонистом первых двух и располагается в пределах эпимеры.
Продольные вентральные мышцы. Первая обычно сохраняется у личинок, и отсутствует у имаго. Вторая соединяет фурки стернитов. От переднегрудной фурки идет к тенториуму и осуществляет движение головы. От каждой ветви фурки проходят ещё две мышцы. Первая из них идет к передней спинальной складке, вторая — к задней. Основная функция данной группы мышц — притягивание сегментов друг к другу.
Дорсо-вентральные мышцы. Данные мышцы являются производными кольцевой мускулатуры аннелид. В группу входят 6 мышц, идущих от тергита к стерниту, либо к частям ноги. Первая мышца является терго-стернальной; её функция — притягивание тергита к стерниту, что приводит к поднятию гребной части крыла. Данная мышца наиболее развита у летающих форм. Вторая мышца обладая той же функцией, располагается от передней спинной мышцы коксоподита. Третья и четвёртая мышцы начинаясь на тергите, выполняют двойную функцию: при неподвижной ноге выполняют функцию вспомогательных поднимателей крыла, а также приводят в движение ногу. Пятая мышца располагается от тергита к внутреннему краю вертлуга, и соединена с ним специальным сухожилием. Сокращаясь, данная мышца опускает ногу, и служит вспомогательным поднимателем крыла. Шестая мышца идет от тергита к мерону, расположенному рядом с коксой и также обладает двойной функцией.
Участие крыльев в полёте

По участию в полете крыльев и крыловой мускулатуры различаются три группы насекомых:
- бимоторные (например, стрекозы, чешуекрылые, Neuropteroidea)
- переднемоторные (перепончатокрылые, чешуекрылые, двукрылые)
- заднемоторные (жесткокрылые, полужесткокрылые)
Бимоторность — в одинаковой степени развитие обеих пар крыльев и приводящие их в движение мышц. Переднемоторность — объединение и содружественность в работе передних и задних крыльев. В ходе такого объединения возникает функциональная двукрылость — обе пары крыльев функционируют как одна. При этом, во время полёта основная нагрузка приходится на переднюю пару крыльев, что порой приводит к ослаблению задней пары. И у отдельных групп насекомых к их исчезновению — функциональная двукрылость, которая может переходить в морфологическую (двукрылые). Заднемоторность — передние крылья плотнее задних и выполняют защитную и покровную функцию.
Складывание крыльев
По А. В. Мартынову[3], все крылатые насекомые подразделяются на две группы: древнекрылые насекомые (Palaeoptera) и новокрылые насекомые (Neoptera).
Древнекрылые насекомые
Древнекрылые насекомые не могут складывать крылья вдоль тела в покое, и держат их распростёртыми или отведенными назад. Новокрылые способны компактно складывать крылья вдоль тела.
Новокрылые насекомые
Складывание крыльев у новокрылых насекомых обеспечивается благодаря особому строению системы склеритов в сочленении дорсальной поверхности крыла и тергита. В типичном случае аксиллярный аппарат новокрылых насекомых имеет два основных сочленения: первого аксиллярного склерита с передним и средним крыловыми отростками нотума и третьего аксиллярного склерита с задним крыловым выступом; при этом и первый, и третий аксиллярные склериты соединены со вторым. По линии, проходящей через эти два сочленения, при складывании крыла образуется складка. На вентральной стороне крыла особый отросток третьего аксиллярного склерита сходится с задним краем субалярного склерита. Кроме того, третий аксиллярный склерит сочленяется ещё и с основаниями анальных жилок крыла, благодаря чему, при складывании крыла образуется анальная складка. Таким образом, третий аксиллярный склерит и лежащий позади него участок крыла за анальной складкой переворачиваются, позволяя крылу повернуться вершиной назад. Переворачивающийся участок крыла был назван неала, а остальная часть крыла — палеала, что отражает представление о том, что нескладывающиеся крылья подёнок, стрекоз и других древних насекомых являются плезиоморфией, а способность к складыванию крыльев у новокрылых — апоморфией.
Адаптации и видоизменения крыльев
У некоторых отрядов насекомых крылья претерпели ряд изменений для выполнения специальных функций. Различия в форме и жилковании передних и задних крыльев практически отсутствуют у примитивных летающих форм, среди хорошо летающих — только у стрекоз. У чешуекрылых и перепончатокрылых заднее крыло отличается от переднего как функциональному значению и по размеру и площади, а у двукрылых превращается в жужжальца. Жилкование заднего крыла у данных групп насекомых примерно соответствует таковому на переднем крыле.
В других группах, наоборот, основная локомоторная функция возлагается на заднее крыло. Например, у веерокрылых переднее крыло также трансформируется в жужжальце. У жесткокрылых переднее крыло превращается в надкрылья, заднее крыло длиннее переднего и в покое складывается пополам, изменяется характер его жилкования.
Ориентация
- У двукрылых задняя пара крыльев превратилась в жужжальца, которые позволяют им лучше ориентироваться в пространстве и изменять траекторию движения в полёте, а также помогают поддерживать равновесие.
- У веерокрылых в жужжальца превратилась передняя пара крыльев. Самки бескрылые вовсе.
Защитная функция
- У жесткокрылых передние крылья приобрели твёрдость и эволюционировали в надкрылья для защиты тонких задних крыльев, скрытых в покое под ними.
- У полужесткокрылых (клопы) передние крылья также могут отвердевать, тем не менее не так выражено, как у жесткокрылых (твёрдой становится лишь передняя часть крыла — полунадкрылье).
- Передние крылья кожистокрылых (уховёртки), прямокрылых (кузнечики, сверчки), богомолов и тараканов стали кожистыми, и не принимают участия в полете.
- У некоторых насекомых из других отрядов передние крылья могут иногда преобразовываться в надкрылья, что часто сопровождается редукцией задних крыльев и способности летать. Также, некоторые особи из вышеприведенных рядов теряют задние крылья.
См. также
Источники
- В. В. Гриценко, Ю. М. Стройков, Н. Н. Третьяков. Вредители и болезни сельскохозяйственных культур. — М.: — «Академия», 2008
- Все о насекомых
- А. В. Мартынов. О двух основных типах крыльев насекомых и их значении для общей классификации насекомых. — Тр. I Всероссийского съезда зоологов, анатомов и гистологов в Петрограде 15-21.XII.1922 г., 1923
Литература
- Triplehorn, Charles A.; Johnson Norman F. (2005). Borror and DeLong’s introduction to the study of insects (7th edition ed.). Thomson Brooks/Cole. ISBN 0-03-096835-6.