Стрекозы
Стреко́зы (лат. Odonáta) — отряд древних летающих насекомых, насчитывающий в мировой фауне свыше 6650 видов. Это относительно крупные насекомые, с подвижной головой, большими глазами, короткими щетинковидными усиками, удлинённым стройным брюшком и четырьмя прозрачными крыльями с густой сетью жилок. Стрекозы — активные специализированные хищники, которые питаются насекомыми, пойманными на лету. Представители отряда широко распространены по миру, встречаясь на всех материках, исключая Антарктиду.
| Стрекозы | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Дозорщик-император (Anax imperator), одна из самых крупных стрекоз Европы | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Отряд: Стрекозы |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Odonata Fabricius, 1793 | |||||||||||||||||||||||||||||||||||||||||
| Подотряды | |||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 233 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Где гнутся над омутом лозы,
Где летнее солнце печет,Летают и пляшут стрекозы,
Веселый ведут хоровод.Дитя, подойди к нам поближе,
Тебя мы научим летать,
Дитя, подойди, подойди же,
Пока не проснулася мать!Под нами трепещут былинки,
Нам так хорошо и тепло,
У нас бирюзовые спинки,
А крылышки точно стекло!Мы песенок знаем так много,
Мы так тебя любим давно -
Смотри, какой берег отлогий,
Какое песчаное дно!
Гляжу: стрекозы
Над водой летают
И крылышками
Еле задевают
ПоверхностьОчарованной реки.
Все представители отряда ведут амфибионтный образ жизни — яйца и личинки развиваются в водной среде, а имаго (взрослые) обитают на суше, освоив воздушную среду и став прекрасными летунами. Взрослые стрекозы не ограничиваются в выборе места обитания одними только берегами водоёмов и могут улетать от них на значительные расстояния, встречаясь на лугах, опушках лесов и даже в населённых пунктах. Развитие с неполным превращением: имеются стадии яйца, личинки и имаго. Личинки (их называют также нимфами или наядами) развиваются в водоёмах различных типов — главным образом в стоячих озёрах, прудах и старицах рек, а также в ручьях, реках и болотистых водоёмах, вплоть до созданных человеком прудов и канав с водой. Как и взрослые стрекозы, личинки являются хищниками. Они охотятся на водных насекомых и прочих беспозвоночных.
Стрекозы имеют большое значение для человека. Велика их роль в регуляции численности кровососущих насекомых, ряда насекомых-вредителей сельского и лесного хозяйства. В некоторых случаях личинки стрекоз могут приносить вред, например, уничтожая мальков в рыбоводных прудах либо составляя им пищевую конкуренцию. Кроме того, личинки некоторых видов могут являться промежуточными хозяевами гельминтов.
Раздел энтомологии, посвящённый изучению стрекоз, называется одонатологией.
Этимология
Русское слово «стрекоза» образовано при помощи редкого суффикса -оз- (ср. «егоза») от глагола стрекать «прыгать» или омонимичного ему стрекать «жалить». По альтернативной этимологии, связано со «стрекотать». Впервые засвидетельствовано в конце XVII века в форме «строкоза»[1][2].
Термин Odonata, используемый в качестве научного латинского названия для обозначения отряда стрекозы, был впервые введён в 1793 году датским энтомологом Иоганном Христианом Фабрицием, который образовал его от греческого слова ὀδούς «зуб» (род. п. ὀδόντος), по-видимому, потому, что у стрекоз имеются зубцы на верхних челюстях, хотя и большинство других насекомых также имеют зубчатые мандибулы[3].
Количество видов и распространение

Стрекозы весьма разнообразны. При этом некоторые описанные виды известны по находкам из единственной местности, а для некоторых, известных по единичным экземплярам, неизвестна личиночная стадия. Биология части видов всё ещё остаётся неизученной[4].
Число описанных видов стрекоз, по данным на август 2013 года, — 6650, включая 608 ископаемых[5]. Ежегодно описывается несколько десятков новых видов, и всего за два года до этого указывалась цифра в 5899 современных видов[6].
Стрекозы распространены на всех континентах, за исключением Антарктиды, также отсутствуют на островах Ледовитого океана, в Исландии и Гренландии[7]. Северная граница глобального ареала стрекоз приблизительно совпадает с границей распространения безморозных территорий и областей, на которых продолжительность периода с устойчивой температурой воздуха ниже 0 °C не превышает 90 дней в году[7]. Широкое распространение стрекоз по миру определяется рядом особенностей, основными из которых являются древнее происхождение отряда, способность к активному передвижению, полное отсутствие пищевой специализации (в то время как многие группы насекомых связаны в своём распространении с растительными группами)[8].
Внутри отряда имеются как очень узко, так и очень широко распространённые группы и виды. Благодаря пластичности жизненных циклов стрекоз, позволяющей им выбирать оптимальные температурные и другие условия воздушной среды, многие виды сформировали обширные ареалы, занимающие разные широтные зоны и вертикальные пояса. Ареалы некоторых стрекоз охватывают несколько природных зон. Так, например, дозорщик-император и стрекоза Фонсколомба распространены на обширной территории от южной Африки до полярного круга в Северной Европе. Наиболее широко распространённым и высоко летающим видом стрекоз является бродяжка рыжая, встречающаяся на всех материках, за исключением Антарктиды[9]. В Гималаях она была зарегистрирована на высоте 6200 м. Этот вид стрекоз стал первым, заселившим атолл Бикини после проведения на нём ядерных испытаний (в 1946—1958 годах), и является единственным видом, обитающим на острове Пасхи[4].

В то же время другие виды имеют ограниченные ареалы либо являются эндемиками. Так, огнетелка Элизабеты известна только в 8 местах обитания на юге Балканского полуострова[10], а булавобрюх увенчанный является эндемиком гор Средней Азии[11]. Большинство современных видов обитает в тропиках и влажных субтропиках[12]. Стрекозы населяют также засушливые районы и пустыни, где развитие их личинок происходит в непересыхающих водоёмах и оазисах. Известен даже ряд видов-эндемиков, обитающих в Сахаре, пустыне Намиб, Аравийской и Сомалийской пустыне в Африке (Ischnura saharensis, Aeshna minuscula, Paragomphus sinaiticus, Agriocnemis sania, Enallagma somalicum, Urothemis thomasi и др.)[13][14]. В южноафриканских засушливых районах основные места обитания большинства видов находятся в саваннах[14].
В России стрекозы широко распространены по всей территории страны (за исключением засушливых областей), их фауна насчитывает около 150 видов[15]. Фауна Казахстана включает 87 видов стрекоз[16][17], фауна Украины — до 76 видов[18][19], а фауна Белоруссии — 68 видов[20][21]. На территории бывшего СССР всего было зарегистрировано 172 вида и 69 подвидов стрекоз, относимых к 52 родам из 10 семейств всех трёх подотрядов. Рекордная по числу видов фауна стрекоз отмечена в Бразилии — 828 видов[22].
Зоогеография
Наибольшим видовым разнообразием стрекоз отличаются индомалайская зона (1665 видов) и неотропика (1640). Далее следуют афротропика (889) и австралийская область (870). Меньше видов в областях с умеренным климатом: палеарктика (560) и неарктика (451)[23]. Распространение и разнообразие стрекоз по зоогеографическим областям (учтены 2942 современных вида и 39 родов для Zygoptera и 3012 видов и 348 родов для Anisoptera[24], а также 4 вида Anisozygoptera[25]):
Zygoptera
| Семейство | Ориентальная | Неотропика | Австралазия | Афротропика | Палеарктика | Неарктика | Океания | Мир |
|---|---|---|---|---|---|---|---|---|
| Hemiphlebiidae | 1 | 1 | ||||||
| Lestidae | 40 | 42 | 29 | 17 | 18 | 18 | 3 | 151 |
| Perilestidae | 19 | 19 | ||||||
| Synlestidae | 18 | 1 | 8 | 11 | 6 | 39 | ||
| Platystictidae | 136 | 43 | 44 | 1 | 1 | 224 | ||
| Amphipterygidae | 4 | 4 | ||||||
| Argiolestidae | 10 | 73 | 19 | 6 | 108 | |||
| Calopterygidae | 66 | 68 | 5 | 20 | 37 | 8 | 185 | |
| Chlorocyphidae | 86 | 17 | 42 | 3 | 144 | |||
| Devadattidae | 6 | 6 | ||||||
| Dicteriadidae | 2 | 2 | ||||||
| Euphaeidae | 65 | 1 | 11 | 68 | ||||
| Heteragrionidae | 51 | 51 | ||||||
| Hypolestidae | 2 | 2 | ||||||
| Lestoideidae | 9 | 9 | ||||||
| Megapodagrionidae | 29 | 29 | ||||||
| Pentaphlebiidae | 2 | 2 | ||||||
| Philogangidae | 4 | 2 | 4 | |||||
| Philogeniidae | 39 | 39 | ||||||
| Philosinidae | 12 | 12 | ||||||
| Polythoridae | 59 | 59 | ||||||
| Pseudolestidae | 1 | 1 | ||||||
| Rimanellidae | 1 | 1 | ||||||
| Thaumatoneuridae | 5 | 5 | ||||||
| Incertae sedis | 25 | 11 | 19 | 9 | 61 | |||
| Coenagrionidae | 193 | 554 | 152 | 202 | 96 | 103 | 91 | 1266 |
| Isostictidae | 41 | 46 | ||||||
| Platycnemididae | 199 | 122 | 70 | 22 | 404 | |||
Anisoptera
| Семейство | Ориентальная | Неотропика | Австралазия | Афротропика | Палеарктика | Неарктика | Океания | Мир |
|---|---|---|---|---|---|---|---|---|
| Aeshnidae | 149 | 129 | 78 | 44 | 58 | 40 | 13 | 456 |
| Austropetaliidae | 7 | 4 | 11 | |||||
| Petaluridae | 1 | 6 | 1 | 2 | 10 | |||
| Gomphidae | 364 | 277 | 42 | 152 | 127 | 101 | 980 | |
| Chlorogomphidae | 46 | 5 | 47 | |||||
| Cordulegastridae | 23 | 1 | 18 | 46 | ||||
| Neopetaliidae | 1 | 1 | ||||||
| Corduliidae | 23 | 20 | 33 | 6 | 18 | 51 | 12 | 154 |
| Libellulidae | 192 | 354 | 184 | 251 | 120 | 105 | 31 | 1037 |
| Macromiidae | 50 | 2 | 17 | 37 | 7 | 10 | 125 | |
| Synthemistidae | 37 | 9 | 46 | |||||
| Incertae sedis | 37 | 24 | 21 | 15 | 2 | 99 | ||
Anisozygoptera
| Семейство | Ориентальная | Неотропика | Австралазия | Афротропика | Палеарктика | Неарктика | Океания | Мир |
|---|---|---|---|---|---|---|---|---|
| Epiophlebiidae | — | — | — | — | 4 | — | — | 4 |
Систематика
Ввиду высокой морфологической специализации стрекозы занимают особое положение в классе насекомых и относятся к инфраклассу древнекрылых, в то же время некоторые энтомологи выделяют их в самостоятельный отдел или инфракласс Libellulones[26].
В настоящее время отсутствует единое мнение о рангах высших таксонов, относимых к стрекозам, их объёмах и родственных связях. Не оспаривается только монофилия, происхождение всех вымерших и современных стрекоз от гипотетического общего предка. Однако обоснование монофилии ни одним автором не приводится[26].
Представителей отряда стрекоз (Odonata) и вымершие группы крылатых стрекозоподобных насекомых, живших в каменноугольном и пермском периодах, обычно объединяют вместе в составе надотряда или клады Odonatoptera Martynov, 1932 sensu Grimaldi & Engel, 2005[27][28][29]. Состав и филогенетические взаимоотношения Odonatoptera являются предметом дискуссии — разные авторы придерживаются различных взглядов на них, что отражается в различиях в опубликованных работах. В состав Odonatoptera включаются: Geroptera, Erasipteridae, Protodonata (Meganisoptera), Campylopteridae, Lapeyriidae, Protanisoptera, Triadophlebioptera, Protozygoptera (включая Archizygoptera), собственно отряд Odonata. Признано, что Odonatoptera представляют собой отдельную от всех Neoptera кладу крылатых насекомых. Существует несколько взглядов на взаимоотношения этой группы с близкими. По мнению одних исследователей, Odonatoptera являются сестринским таксоном к Ephemeropteroidea (подёнки и другие), а Palaeodictyopteroidea образуют сестринскую или базальную группу внутри монофилетической группы Palaeoptera. Согласно взглядам других учёных, Odonatoptera — сестринский таксон по отношению ко всем Neoptera, а группа «Palaeoptera» является парафилетической[30][31].
Иногда Odonata рассматриваются в более широком объёме со включением в эту группу почти всех вышеназванных таксонов (Odonata sensu lato)[32], за исключением «Erasipteridae», Geroptera и Protodonata; эти группы трактуются в качестве безранговой клады Odonatoclada[29][33].
Классификация
Стрекоз делят на три подотряда: равнокрылых (Zygoptera), разнокрылых (Anisoptera) и Anisozygoptera (4 современных вида в Японии и Индии). В настоящее время учёным известно более 6650 видов стрекоз, группируемых в примерно 40 семейств[34][35].
Отряд Odonata — Стрекозы
- Подотряд Anisoptera — Разнокрылые стрекозы
- Надсемейство Aeshnoidea
- Семейство Aeshnidae — Коромысла, 456 видов
- Семейство Austropetaliidae — 11 видов
- Надсемейство Petaluroidea
- Семейство Petaluridae — 10 видов
- Надсемейство Gomphoidea
- Надсемейство Cordulegastroidea
- Семейство Chlorogomphidae — 47 видов
- Семейство Cordulegastridae — Булавобрюхи, 46 видов
- Семейство Neopetaliidae — 1 вид
- Надсемейство Libelluloidea
- Семейство Synthemistidae — 46 видов
- Семейство Macromiidae — 125 видов
- Семейство Corduliidae — Бабки, 154 вида
- Семейство Libellulidae — Настоящие стрекозы, 1037 видов
- Роды incertae sedis — 19 родов с 98 видами
- Надсемейство Aeshnoidea
- Подотряд Anisozygoptera
- Семейство Epiophlebiidae — 4 вида
- Подотряд Zygoptera — Равнокрылые стрекозы
- Надсемейство Lestoidea
- Семейство Hemiphlebiidae — 1 вид
- Семейство Perilestidae — 19 видов
- Семейство Synlestidae — 38 видов
- Семейство Lestidae — Лютки, 153 вида
- Надсемейство Platystictoidea
- Семейство Platystictidae — 224 вида
- Надсемейство Calopterygoidea
- Семейство Amphipterygidae — 14 видов
- Семейство Argiolestidae
- Семейство Calopterygidae — Красотки, 185 видов
- Семейство Chlorocyphidae — 144 вида
- Семейство Devadattidae
- Семейство Dicteriadidae — 2 вида
- Семейство Euphaeidae — Ложнокрасотки, 68 видов
- Семейство Heteragrionidae
- Семейство Hypolestidae
- Семейство Lestoideidae
- Семейство Megapodagrionidae — 30 видов
- Семейство Pentaphlebiidae
- Семейство Philogangidae
- Семейство Philogeniidae
- Семейство Philosinidae
- Семейство Polythoridae
- Семейство Pseudolestidae
- Семейство Rimanellidae
- Семейство Thaumatoneuridae — 1 вид
- Надсемейство Coenagrionoidea
- Семейство Isostictidae
- Семейство Platycnemididae — Плосконожки, 404 вида
- Семейство Coenagrionidae — Стрелки, примерно 1267 видов
- Надсемейство Lestoidea
Эволюция стрекозоподобных насекомых
Стрекозы — одни из наиболее древних групп насекомых. Данная группа отделилась от примитивных скарабеоновых насекомых (Scarabaeones) ещё в раннем каменноугольном периоде[26]. В прошлом стрекозы также были многочисленны и разнообразны, о чём свидетельствуют довольно частые находки их остатков в отложениях разных геологических возрастов. В то же время в геологической истории стрекоз ещё остаётся немало пробелов[8].

В составе группы примитивными считаются гигантские стрекозоподобные насекомые Meganeurina ввиду отсутствия у них структур, характерных для крыла современных стрекоз, таких как узелок и птеростигма. Эти насекомые появились в конце раннего каменноугольного периода. Основное направление эволюции меганевриновых заключалось в увеличении размеров, самые древние и примитивные из них (представители семейства Eugeropteridae) были невелики — длина крыльев не превышала 4—5 см. В раннем пермском периоде среди этих стрекоз появились настоящие гиганты: у Meganeuropsis размах крыльев превышал 70 см[8], что делает их наиболее крупными из всех известных насекомых. В позднем каменноугольном — раннем пермском периодах меганеврины достигли своего расцвета, широко расселившись в тропических болотистых лесах. К концу пермского периода их разнообразие снизилось, и в триасовый период перешло только одно семейство Triadotypidae. Во второй половине перми меганевриновые стрекозы дали начало двум группам потомков: Ditaxineurina и Kennedyina (инфраотряд Kennedyomorpha) были преимущественно пермскими, а Triadophlebiina со стебельчатыми крыльями — эндемичной для триасового периода[8]. К подотряду Kennedyina принадлежали мелкие и средней величины насекомые с резко стебельчатыми крыльями длиной до 9 см, которые в покое косо складывались над брюшком дорсальными поверхностями внутрь. У всех представителей Kennedyina имелась птеростигма и был развит узелок, а «ловчая корзина», образуемая ногами и используемая для хватания добычи, была не выражена. Судя по строению ног, эти насекомые охотились, хватая сидящих насекомых[26]. Наиболее близкими к гигантским меганевриновым стрекозам были дитаксиневриновые, чьи ноги уже образовывали «ловчую корзину», а на крыльях появились узелок и птеростигма. Дитаксиневриновые достигли расцвета в раннем пермском периоде и вымерли с его окончанием[26].
.jpg.webp)

В юрском периоде на смену палеозойским группам пришли мезо-кайнозойские группы: Libellulomorpha и Lestomorpha, имевшие водных личинок, а также обладавшие более совершенным летательным аппаратом[26]. Первые представители современной группы Libellulida известны по немногочисленным находкам из триасовых отложений в Австралии, Африке и Южной Америке, где меганевриновые к тому времени уже практически вымерли. В фауне Евразии тех времён всё ещё преобладали меганевриновые, сильно изменившиеся, но ещё многочисленные и разнообразные. Быстрое развитие подотряда началось во второй половине триасового периода, но его распространение на то время всё ещё ограничивалось континентами Южного полушария. С наступлением раннего юрского периода они достигли вершины своей эволюции и заняли господствующее положение среди стрекоз Евразии[8]. В отличие от своих пермских аналогов стебельчатокрылые Libellulida унаследовали от предков «ловчую корзину», а с ней вместе и способность хватать насекомых как на субстрате, так и в воздухе. Водное дыхание личинок совершенствовалось иначе, чем у представителей предыдущей группы. Частично газообмен осуществлялся, как и у предков, дыхательным эпителием ректума, но его также дополняли наружные пластинчатые жабры — производные от эпи- и парапроктов. Позднее, при колонизации текучих водоёмов, первичные жабры видоизменялись, утратили способность к газообмену, и их заменили наружные жабры другого происхождения и строения[8].
В это же время предки калоптеригиновых (Calopterygoidea) гораздо дальше эволюционно продвинулись от своего исходного состояния. Их крылья сузились и стали стебельчатыми, у них исчезли отличия в форме обеих пар крыльев, а их жилкование в большинстве таксонов частично редуцировалось. Помимо них, в юрском периоде широкое распространение получают стрекозы подотряда Anisozygoptera, в меловом периоде уже гораздо менее многочисленные. Стрекозы подотряда Anisoptera в этот период представлены обособленным вымершим семейством, близким к современным реофильным Cordulegastridae. Почти совершенно не встречаются в мелу стрекозы Zygoptera. Для позднемелового-палеогенового периода характерными являются многочисленные Zygoptera, особенно Agrionoidea, однако их личинки редки, что указывает на развитие их в основном вне озёр. На протяжении всего палеогена довольно широко распространены Aeshnidae, но почти отсутствуют Lestidae, Coenagrionidae, Libelluloidea; редки Chironominae; совершенно отсутствуют Limnephilidae и прочие. Кайнозойская фауна стрекоз в общих чертах соответствует современной. К началу неогена энтомофауна приобретает в целом современный облик. Количество Zygoptera резко сокращается, Lestidae и Coenagrionidae, наоборот, становятся более многочисленными, а другие группы (прежде всего Agrionoidea) — редкими[36].
Филогенетическое дерево разных групп стрекоз по данным Bechly (2003) с дополнением по Petrulevicius и Gutiérrez (2016)[37][38]:
| Odonatoptera |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Размеры


Размеры представителей группы колеблются в довольно широких пределах. Одними из самых мелких стрекоз являются азиатские виды — Nannophya pygmaea при длине тела 15 мм и размахе крыльев 20 мм[39][40] и Agriocnemis femina с размахом крыльев 17—20 мм[41].
Крупнейшей современной стрекозой является Megaloprepus caerulatus, обитающая в Центральной и Южной Америке, с длиной тела до 100 мм и с размахом крыльев до 191 мм[42].
К крупнейшим представителям отряда по размаху крыльев также относятся обитающая на Борнео и в Малайзии Tetracanthagyna plagiata[43] с размахом крыльев до 163 мм при длине тела до 100 мм[42][44]; австралийская Petalura ingentissima с длиной тела до 116—120 мм при размахе крыльев до 165 мм[42] и эндемик Гавайских островов Anax strenuus — до 150 мм[45][46]. Самыми тяжёлыми среди всех стрекоз признаны самки Tetracanthagyna plagiata[47].
Самым длинным телом среди стрекоз обладают виды, обитающие в Центральной и Южной Америке: Mecistogaster linearis и Pseudostigma accedens[42], чья длина достигает 135 мм[48], Pseudostigma aberrans — до 144 мм[42], Mecistogaster lucretia — до 150 мм при размахе крыльев до 125 мм[42][44].
Морфология
К отряду относятся крупные и очень крупные, хорошо летающие, обычно ярко окрашенные насекомые, сочетающие в себе общую примитивность строения со многими чертами специализации[49][50].
Внешнее строение
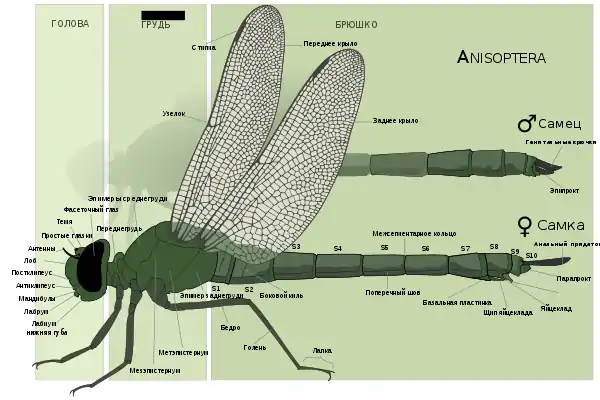
Как у всех насекомых, тело стрекоз состоит из трёх главных отделов: головы, груди и брюшка. Снаружи тело защищено твёрдым хитиновым покровом, образующим наружный скелет.
Строение головы

Голова крупная, шире всех остальных отделов тела. Она обособлена от переднегруди и сочленена с ней чрезвычайно подвижно, благодаря чему способна поворачиваться вокруг продольной оси тела на 180°. Бо́льшую часть головы занимают очень крупные полушаровидные сложные фасеточные глаза. У равнокрылых стрекоз они располагаются строго по бокам головы, отчего её форма поперечно вытянутая; разделяются они промежутком, превышающим по ширине диаметр глаза. У разнокрылых стрекоз глаза более или менее смещены ко лбу. У Gomphidae они разделены промежутком, меньшим, чем диаметр глаз. У булавобрюхов глаза соприкасаются в одной точке, а у настоящих стрекоз и коромысел — на протяжении некоторого отрезка[50].
Зрение играет большую роль в жизни стрекоз. Его особенности обусловлены фасеточным строением глаз, которые состоят из большого числа омматидиев. Наибольшее их количество у стрекоз достигает 10—28 тысяч[51]. При этом нижние омматидии способны различать только цвета, а верхние — только форму предметов. Подобная особенность помогает стрекозам ориентироваться в пространстве и находить подходящую добычу. Темя вздутой формы, располагается между глазами, если глаза разделены, или спереди от них, если глаза соприкасаются. Детали его строения различаются у представителей разных групп. На темени всегда располагается три простых глазка (ocelli). У большинства стрекоз они образуют равносторонний треугольник, у Gomphidae и Aeshnidae располагаются практически на одной прямой, а у остальных стрекоз — по краю теменного вздутия. Впереди, книзу от темени, последовательно расположены лоб, наличник и верхняя губа, прикрывающая сверху ротовой аппарат. Усики малозаметные — короткие и тонкие, шиловидные, образованы 4—7-члениками. Их базальный членик наиболее короткий и толстый, далее членики постепенно утончаются. Ротовой аппарат мощный, хорошо развитый, грызущего типа. Он образован непарными верхней и нижней губами, парными острозубчатыми верхними (мандибулы) и нижними (максиллы) челюстями. Верхняя губа прикрывает сверху челюсти, она в виде короткой пластинки, вытянутой в поперечном направлении, с полукруглым внешним краем. Нижняя губа из трёх лопастей (средней и двух боковых) покрывает собой челюсти снизу. Она крупнее верхней, наиболее развита у разнокрылых стрекоз, которые жуют добычу на лету. Парные челюсти размельчают пищу, двигаясь при этом в горизонтальной плоскости. Челюсти стрекоз чрезвычайно прочные и характеризуются острыми твёрдыми зубцами. На максиллах также имеются щупики, выполняющие осязательную роль[50].
.jpg.webp)
Строение груди
Грудь хорошо развитая; как и у всех насекомых, состоит из переднегруди, среднегруди и заднегруди. Каждая из частей несёт на себе одну пару конечностей, а среднегрудь и заднегрудь ещё и по паре крыльев каждая. Целый ряд особенностей строения груди стрекоз является своеобразным. Переднегрудь отделена от среднегруди, сильно редуцирована (иногда практически целиком помещается в углублении задней стороны головы). Задне- и среднегрудь хорошо развитые и плотно сросшиеся между собой, образуют синторакальный комплекс, или синторакс, который воспринимается как собственно грудь и часто так и называется. Склериты каждого сегмента отделены друг от друга швами. Боковой шов между средне- и заднегрудью развит только у представителей семейства Calopterygidae[50].
Средне- и заднегрудь характеризуются чрезвычайно сильно развитыми мезо- и метаплевритами. Их тергиты и стерниты сильно уменьшены, а плевриты — наоборот, чрезвычайно развиты. Из-за этого вся грудь сплюснута с боков. Помимо этого, в отличие от всех остальных насекомых, спинная часть груди у стрекоз косо отодвинута назад, при этом среднегрудь иногда расположена выше заднегруди. Из-за этого крылья оказываются прикреплёнными позади ног. У многих равнокрылых стрекоз передняя пара крыльев располагается даже скорее прямо над задней парой, нежели впереди неё. Верхняя часть переднегруди (тергит) называется переднеспинкой. Она разделена на три следующие друг за другом доли — переднюю, среднюю и заднюю. Средняя обычно самая крупная и имеет продольную вмятину[50][52].
.jpg.webp)
Крылоносные сегменты представлены гипертрофированными плейритами. Таким образом, движения крыльев, в отличие от всех других насекомых, у стрекоз обеспечиваются мышцами прямого действия, которые позволяют им совершать несинхронные взмахи. Эти мышцы, будучи непосредственно связанными с крыловыми пластинками, придают их движениям особую силу. Независимость в работе передних и задних крыльев обусловливает манёвренность полёта, по скорости которого многие виды крупных стрекоз превосходят бражников и слепней[51].
Строение крыльев
Крыльев две пары. Они очень крупные, их длина близка к общей длине тела[49].
У представителей Zygoptera и Anisozygoptera обе пары крыльев одинаковые по форме и жилкованию (передние чуть длиннее) и прикрепляются к синтораксу всем своим основанием, стебельчато суженным[52]. У Anisoptera крылья заметно различны, задние крылья отличаются по форме от передних — они шире у основания[52]. К телу передние крылья прикрепляются всем основанием, а задние крылья в своей базальной части несколько шире передних и прикрепляются только верхней, стебельчато суженной частью расширенного основания[50].
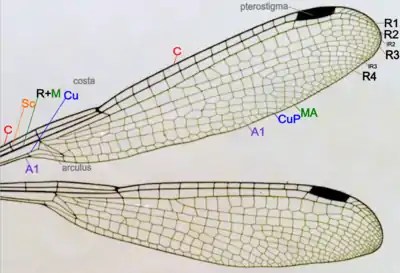
Крыло стрекоз образовано двумя хитиновыми слоями, каждый из которых имеет свою собственную сеть жилок. Жилки накладываются одни на другие, поэтому вся сеть жилкования внешне выглядит единой. Жилки крыла представляют собой трахеи; более крупные из них заполнены гемолимфой, более мелкие лишены просвета. Жилкование крыльев очень густое и сложное. У стрекоз почти все промежутки между основными жилками крыла заняты сетью более мелких жилок и анастомозов (первичная сеть — архедиктион)[51], между которыми образуются ячейки и поля.
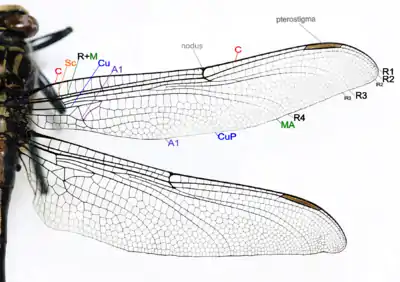 |
 | |
Общий план строения крыльев разнокрылой стрекозы |
Общий план строения крыльев равнокрылой стрекозы |
На каждом крыле имеется шесть основных продольных жилок[49][52]:
- Костальная жилка (costa, С) — образует собой весь передний (костальный) край крыла.
- Субкостальная жилка (subcosta, Sc) — располагается параллельно костальной жилке на расстоянии одного ряда ячеек, но короче неё.
- Радиальная жилка (radius, R) — представляет собой целую систему ветвей, называемых секторами. От основания крыла радиальная жилка слита со следующей, медиальной жилкой (medius, M); этот отрезок обозначается как R+M.
- Кубитальная жилка (cubitus, Си) — не ветвясь, идёт от основания крыла до средней части его заднего края, параллельно медиальной жилке, лишь в конце загибаясь назад.
- Анальная жилка (А) — проходит более или менее параллельно кубитальной жилке и заканчивается на заднем крае крыла.
Представители различных подотрядов стрекоз имеют принципиально одинаковую схему жилкования и отличаются только деталями расположения и ветвления жилок, а также формой отдельных ячеек и полей. В основании крыла, между медиальной и кубитальной жилками, у равнокрылых стрекоз находится поле, носящее название четырёхугольника. У разнокрылых стрекоз расположенное в этом месте поле называется треугольником. Непрозрачное поле в основании крыла, свойственное большинству разнокрылых стрекоз, получило название перепоночки[52]. Важной особенностью строения крыла стрекоз является наличие птеростигмы и узелка. Птеростигма — утолщение у переднего края крыла, расположенное ближе к его вершине. Она состоит из утолщённых (иногда дополнительно пигментированных) жилок и ячеек и внешне выглядит как плотный тёмноокрашенный участок края крыла. Птеростигма увеличивает амплитуду размаха крыла за счёт утяжеления его вершины и, вероятно, способна гасить вредные колебания (флаттер) крыла[53]. Среди поперечных жилок следует отметить узелок (nodulus) — сильно утолщённую короткую жилку в средней части переднего края крыла, соединяющую костальную и первую радиальную жилки. Он играет роль шарнира, позволяющего крылу скручиваться вдоль продольной оси[50].
Складывание крыльев с изменением их формы у стрекоз невозможно. В покое крылья распростёрты (иногда почти опущены вниз) либо приподняты и прижаты друг к другу вдоль брюшка либо над ним. Стрекозы способны к автономным движениям каждого крыла. Многие стрекозы могут совершать крыльями до 10—15 взмахов в секунду[51].
Строение конечностей

Конечности стрекоз (ноги) состоят из 5 основных частей — тазика, вертлуга, бедра, голени и лапки, которые соединены подвижными суставами. Неподвижно соединены лишь дистальный конец вертлуга и проксимальный конец бедра, сросшиеся друг с другом. Форма ног практически постоянна между представителями; их длина увеличивается от передней пары к задней. Расстояния между парами ног несколько неравные — средняя пара ближе к задней, чем к передней[50].
Тазик толще и короче всех остальных частей ноги. Вертлуги, напротив, длинные, с перетяжкой в средней части, гладкие (крайне редко несут шипики). Они сочленены с бедром косо. Бёдра и голени ещё длиннее, но первые несколько толще и короче вторых. Те и другие снабжены по бокам двумя рядами шипиков, направленных книзу. Один ряд разделяет переднюю и нижнюю поверхности голени, а второй — заднюю и нижнюю поверхности бёдер. Длина и густота расположения шипиков у представителей различных групп стрекоз различны. Так, у красоток шипики на бёдрах и голенях равные, а у коромысел и плоскобрюхов первые несколько длиннее последних. Помимо этого, длина и густота шипиков могут различаться на разных парах ног, что служит таксономически важным признаком. Иногда ряды шипиков могут быть двойными (у Cordulegaster) либо даже тройными. Лапка всегда трёхчлениковая, длина её сегментов увеличивается от проксимального к дистальному. На каждом членике располагается по два ряда шипиков. Дистальный членик лапки несёт пару острых раздвоенных коготков с острым зубцом на нижней стороне. Между основаниями коготков имеется небольшой отросток. Все сегменты лапки подвижно сочленены, однако их подвижность ограничивается хитиновыми пластинками и отростками[50].
Конечности практически не используются стрекозами для передвижения и служат им только для посадки и взлёта. При этом передняя пара ног служит в первую очередь для схватывания и удерживания добычи[50][52].
Строение брюшка
_(4).JPG.webp)

Брюшко очень длинное, палочкообразное. Чаще оно в сечении округлое, тонкое, реже — широкое и плоское[52]. На его долю приходится до 90 % от всей длины тела. Брюшко состоит из десяти полных сегментов и одиннадцатого рудиментарного сегмента (тельсон). Каждый сегмент образован двумя хитиновыми щитками. Верхний щиток (тергит) дуговидный в поперечном сечении, заходя на брюшную сторону, частично перекрывает нижний (стернит). Последний, в отличие от тергита, имеет слабо выгнутую форму и покрывает брюшко только снизу. По бокам между тергитами и стернитами располагаются растяжимые плевральные перепонки, позволяющие стрекозе изгибать брюшко, а также сжимать и разжимать его при дыхании и откладывании яиц. С каждой стороны на каждом сегменте (кроме IX и X) на плевральной перепонке находится по одному дыхальцу (стигме). Характерная черта строения брюшка самцов большинства видов коромысел и булавобрюхов — наличие продольных выростов на каждой стороне нижней поверхности второго тергита — так называемых ушек (auriculae). Рудиментарные ушки имеются и у самок многих представителей семейства Gomphidae[50].
Ряд деталей строения II, VIII и IX сегментов брюшка у стрекоз несколько специфичен, что связано с половыми различиями. У самок на заднем крае стернита VIII сегмента располагается половое отверстие, а на II и IX сегментах любые отверстия и дополнительные структуры отсутствуют. У самцов же VIII сегмент лишён отверстий; на конце стернита IX сегмента имеется первичное половое отверстие, а на стерните II сегмента — вторичный копулятивный орган[50].
На конце брюшка располагаются нечленистые анальные придатки, число которых у самок постоянно и равно 2, а у самцов стрекоз разных таксономических групп — либо 4 (по паре верхних и нижних), либо 3 (пара верхних и непарный нижний). Генитальные органы у самок располагаются на конце брюшка. У самцов копулятивный орган — так называемый вторичный копулятивный аппарат[49] — располагается на брюшной стороне второго сегмента брюшка, при этом первичное спермовыводящее отверстие находится на стерните десятого сегмента[50].
Органы размножения
Строение репродуктивных органов имеет большое значение в систематике стрекоз. Таксономическую принадлежность представителей многих групп возможно точно определить только при использовании данных признаков[50].
Самцы
Строение системы репродуктивных органов у самцов стрекоз уникально и не имеет аналогов ни в одной другой группе насекомых. Первичное спермовыводящее отверстие, в которое открываются протоки семенников, расположено у самцов на задней линии IX сегмента и прикрыто парными хитиновыми клапанами. Однако в процессе спаривания данное отверстие непосредственного участия не принимает. Для этого предназначен особый вторичный копулятивный аппарат, расположенный в генитальном кармане на стерните II сегмента. Сперма перекачивается сюда из первичного отверстия перед началом процесса спаривания, для чего самец пригибает конец брюшка к его II сегменту. Вторичные половые органы у самцов образованы пенисом, пузырьком, находящимся позади него, и расположенными по бокам от него парными придатками — генитальными крючочками. У представителей Aeshnoidea и Cordulegastroidea крючочков две пары, у Libelluloidea — одна, гомологичная задней паре у первых двух групп. Генитальные крючочки служат для удерживания конца брюшка самки во время спаривания. Строение крючочков весьма разнообразно и имеет важное значение для систематики стрекоз. Также у Aeshnoidea и Cordulegastroidea между крючочками, впереди от пениса, имеется непарная защитная шапочковидная хитиновая пластинка. У Libelluloidea она отсутствует, и вместо неё имеется пара направленных вниз и назад пластинок[50].
Самки
_male_lateral.jpg.webp)
У самок стрекоз наружное половое отверстие располагается на заднем конце VIII стернита (фактически на границе VIII и IX стернитов). Оно может быть открытым, но чаще всего прикрыто выступом или отростком заднего края стернита, носящего название генитальной пластинки. У всех Zygoptera, а также у Aeschnidae, яйцеклад самок имеет вид трубки, образованной желобковидными придатками генитальной пластинки, заострёнными на концах и прижатыми друг к другу своими вогнутыми сторонами. Каждый из придатков образован двумя изогнутыми хитиновыми полосками, называемыми створками яйцеклада. У некоторых групп имеется также вторая и третья пара створок яйцеклада. Эти две пары получили название генитальных клапанов. Створки третьей пары на конце имеют тонкий генитальный отросток (грифелёк), который у Anisoptera несёт небольшой пучок волосков, у Zygoptera отсутствующий. У представителей многих групп Anisoptera яйцеклад подвергся значительной (в ряде случаев — полной) редукции. Так, у Libellulidae и Gomphidae вторые и третьи створки почти или полностью отсутствуют, а первые сильно уменьшены. У булавобрюхов яйцеклад образован только первой парой створок и является весьма длинным, сильно выдаваясь за конец брюшка. Однако у них он не функционирует по прямому назначению, а служит для проделывания отверстий в субстрате, куда откладываются яйца, переносимые туда непосредственно из полового отверстия[50].
Окраска
Окраска тела и крыльев представителей отряда весьма разнообразна; часто она является характерной для отдельных видов или иных таксономических групп.
У стрекоз, как и других групп насекомых, различают структурную и пигментную окраску[51]. У некоторых видов отмечается сочетание этих двух типов окраски. Структурная окраска связана с некоторыми специфическими особенностями поверхностной структуры кутикулы (микроскопическими рёбрышками, пластинками и т. п.), которые создают эффекты интерференции, дифракции и дисперсии света, результатом чего являются металлически блестящие оттенки в окраске[54].
Пигментная окраска обусловлена пигментами, которые наиболее часто содержатся в экзокутикуле либо клетках гиподермы. После гибели насекомого гиподерма разрушается, что приводит к изменению цвета[54]. Наиболее распространённые пигменты — меланины, дающие тёмно-коричневую, коричнево-красную или чёрную окраску. Также распространены каротиноиды, обеспечивающие жёлтый, оранжевый, красный цвета; флавоноиды, дающие жёлтый, белый, красный и коричневый цвета. Некоторые пигменты (особенно чёрные и бурые) отличаются большой стойкостью, в то время как красные, синие и зелёные неметаллические пигменты обычно являются менее стойкими и после гибели стрекоз часто изменяют свой цвет на коричневый, жёлтый, буро-жёлтый, рыжий. К гиподермальным относят голубые и зелёные участки на теле стрекоз, которые посмертно становятся серо-бурыми[54].
Окраска тела стрекоз может быть одноцветной либо представлять собой комбинацию из нескольких цветов (обычно из двух-трёх, реже — большего числа цветов). На общем фоне тела часто имеются полосы и (или) пятна другого цвета. У большинства стрекоз крылья прозрачные и бесцветные, но у целого ряда видов они могут быть полностью или частично (наиболее часто — в области у основания) окрашенными либо же затемнёнными. Полупрозрачная окраска может быть различной: жёлтой, оранжевой, рыжей, зелёной, голубой. Сочетания различных цветов на одном крыле встречаются крайне редко. У других стрекоз крылья могут иметь непрозрачные участки в виде полос и пятен, как правило — чёрные или бурые, реже — ярких тонов[50].
 Пример структурной окраски. Самец красотки-девушки
Пример структурной окраски. Самец красотки-девушки Пример пигментной окраски. Самец Trithemis aurora
Пример пигментной окраски. Самец Trithemis aurora.jpg.webp) Rhyothemis resplendens — один из видов с непрозрачными участками на крыльях
Rhyothemis resplendens — один из видов с непрозрачными участками на крыльях
Половой диморфизм и полиморфизм
Половой диморфизм — различия во внешнем виде и окраске тела самца и самки — широко распространён и часто весьма заметен у стрекоз. Он играет определённую роль в их репродуктивном поведении. Окраска стрекоз изменяется с возрастом и окончательно устанавливается только к моменту половой зрелости.
У стрекоз отмечены разные варианты полового диморфизма. У одних видов самки крупнее самцов (соответственно 6,1 и 5,7 см у Dromogomphus spinosus; 3,1 и 2,7 см у Enallagma antennatum), у других, наоборот, самки мельче самцов (11 и 14 см у Pseudostigma aberrans; 4,6 и 5,1 см у Orthemis ferruginea; 3,4 и 4,1 см у Hetaerina americana)[55]. На уровне семейств также наблюдаются разные тенденции: у красоток (Calopterygidae) самцы в среднем крупнее, чем самки, а у стрелок (Coenagrionidae) самки крупнее самцов[56]. Однако в первую очередь половой диморфизм у стрекоз выражается в различной окраске. Крыловой узор также, вероятно, способствует видовому распознаванию[8]. У бродяжки рыжей половой диморфизм по окраске наблюдается в континентальных популяциях, но не выражен в островных, однако островные популяции демонстрируют больший половой диморфизм, чем континентальные популяции, по иным морфологическим характеристикам: по длине крыльев, размерам брюшка и ног[57].
В ряде случаев у стрекоз наблюдается полиморфизм — наличие нескольких внешне отличающихся по окраске форм одного вида, не связанных с половыми различиями. Так, для самок тонкохвоста изящного описан полиморфизм окраски, включающий три различные цветовые морфы, одна из которых сходна по окраске с самцами[58].
 Самец Libellula depressa
Самец Libellula depressa Самка Libellula depressa
Самка Libellula depressa Самец (слева) и самка красотки-девушки
Самец (слева) и самка красотки-девушки
Морфология преимагиальных стадий
Яйца
Форма яиц бывает овальной (соотношение длины и ширины 1,5:1, 2:1) или длинной и узкой (5:1). Удлинённые яйца характерны для равнокрылых стрекоз и коромысел (Aeshnidae), а округлые и овальные — для остальных разнокрылых. Яйца могут быть окрашены различно — оттенками от светло-жёлтых до тёмно-бурых. У некоторых видов яйца окружает студенистая масса, а у стрекозы двупятнистой и других представителей семейства бабки комки яиц собраны в студенистые жгуты[59].
Личинки

Личинок стрекоз часто называют нимфами либо наядами[49]. По внешней морфологии они отличаются от взрослых стрекоз в большей степени, чем личинки в других группах насекомых с неполным превращением, в первую очередь по строению дыхательной системы и ротового аппарата. Это объясняется их водным образом жизни, а не наземным, как у половозрелой стадии развития[49].
Тело у личинок имеет различную форму. У представителей одних групп оно очень короткое и толстое, совершенно не похоже на тело взрослой стрекозы. В других группах — более удлинённое и стройное, больше напоминает тело имаго, но всегда толще и короче его. Голова у личинок стрекоз крупная и широкая и, за исключением ротового аппарата, устроена так же, как у имаго. Однако она плотно сращена с грудью и не обладает такой подвижностью, как у взрослых стрекоз. На голове выделяются сложные глаза, простые глазки, антенны, лоб, наличник, щёки, темя, затылок, виски, ротовые органы[60]. Сложные глаза всегда располагаются по бокам головы. Они несколько мельче, чем у имаго, но по строению и функции не отличаются от них[60]. Простые глазки у личинок разнокрылых стрекоз появляются после первых линек, а у личинок равнокрылых стрекоз — только на последних стадиях личиночного развития[50]. Органы чувств у личинок развиты так же хорошо, как и у взрослых стрекоз. Наиболее развиты органы зрения, вкуса и осязания. Органы обоняния и слуха (хордотональные органы) развиты слабо. Последние располагаются на ногах — на вертлуге и у основания голени. К слуховым органам также относится тимпанальный орган, расположенный на антеннах[61].
Характерной чертой строения головы личинок стрекоз является ротовой аппарат, состоящий из непарной верхней губы, парных верхних челюстей, парных нижних челюстей и непарной нижней губы. Нижняя губа имеет очень своеобразное строение и преобразована в специальное хватательное приспособление для ловли добычи — маску, являющуюся уникальным органом личинок стрекоз[50][62][60]. При захвате добычи она стремительно раскрывается и выбрасывается вперёд, при этом зубцы на её переднем конце глубоко вонзаются в жертву. Когда маска складывается, добыча подтягивается ко рту и спокойно пережёвывается[61]. В состоянии покоя маска прикрывает голову снизу и (или) с боков[62]. Особенности строения маски, наличие или отсутствие на ней щетинок, а также их количество, имеют большое значение в систематике при определении таксономической принадлежности личинок[61]. Количество щетинок в целом варьирует от 1 до 10 и более на каждой стороне подбородка (у Aeschnidae и Gomphidae они отсутствуют)[50]. Маска состоит из подподбородка (submentum), подбородка (mentum), дистальной лопасти, слитой с подбородком, и губных щупиков, преобразованных в боковые лопасти. Подбородок своей вершиной подвижно сочленяется с пластинкой подподбородка, который в свою очередь подвижно соединяется с головой личинки, вследствие чего маска может выдвигаться вперёд и затем складываться[62].
.jpg.webp)
Существует два типа масок: плоская и выпуклая (шлемовидная)[60]. Плоская маска в состоянии покоя прикрывает рот личинки только снизу (её средняя пластинка более или менее плоская, боковые лопасти лежат вместе с ней в одной плоскости), а шлемовидная маска прикрывает одновременно сверху, спереди и снизу, как забрало или черпак. Этот тип характерен, например, для представителей родов коромысла и дозорщики, которые хватают добычу большими подвижными зубцами на её боковых лопастях. Второй тип масок характерен для личинок, обитающих в иле, например, у таких родов, как плоскобрюхи и булавобрюхи. Такие личинки захватывают добычу вместе с илом, а затем мелкие частицы просеиваются через своеобразное «сито», образованное длинными средними и боковыми щетинками на маске[60][61].
Грудь у личинок стрекоз принципиально устроена также, как и у имаго. Задне- и среднегрудь образуют единый комплекс (синторакс), а переднегрудь обособлена от них. Конечности прикрепляются к отделам груди по тому же принципу, что и у взрослых стрекоз. К соответствующим отделам синторакса также прикреплены крыловые чехлики: передние — к среднегруди, задние — к заднегруди. На верхней стороне всех сегментов груди часто имеются бугорки, шипы либо волоски. Имеются обе пары дыхалец, характерные для взрослых стрекоз, которые, однако, не функционируют до самой последней стадии развития[50]. Ноги личинок принципиально не отличаются от конечностей взрослых стрекоз, но являются более длинными относительно тела. Особенно это характерно для только что вылупившихся личинок, имеющих паукообразный облик[60]. По мере роста личинки относительная длина конечностей уменьшается, но у личинок настоящих стрекоз и родственных им групп паукообразный вид сохраняется до конца личиночной стадии. Характерной особенностью конечностей личинок является строение вертлуга — его мышцы не входят в основание бедра, а прикрепляются к натянутой мембране, поддерживающей бедро. Если схватить личинку за ногу, мышцы вертлуга сокращаются, что вызывает разрыв мембраны, и бедро с лёгкостью отделяется от тазика. На ранних стадиях развития личинки могут регенерировать утраченную конечность, на поздних — нет[50].
.jpg.webp)
Зачатки крыльев у личинок появляются после третьей или четвёртой линьки и имеют вид коротких треугольных отростков. С возрастом они постепенно увеличиваются, достигая конца четвёртого или начала пятого сегмента брюшка у личинок последних возрастов. У большинства стрекоз они располагаются параллельно спинной стороне груди и тесно примыкают друг к другу, но у некоторых групп они являются расходящимися (Cordulegastridae и некоторые Libellulidae)[50].
Брюшко личинок весьма изменчиво по своему строению, форме и размерам, но всегда гораздо шире и короче, чем у взрослых стрекоз. Брюшко состоит из 12 сегментов (а не из 11, как у имаго), 10 из которых являются полностью развитыми, а 2 последних — рудиментарными[49]. Сегменты брюшка несут по бокам небольшие пластинчатые плевриты (у Anisoptera прямые, у Zygoptera изогнутые). На брюшной стороне I—VIII сегментов расположены парные дыхальца. В норме они не используются личинкой для дыхания, за исключением периода, предшествующего метаморфозу, и в редких случаях при высыхании водоёмов. У Anisoptera брюшко заканчивается анальной пирамидой, а у Zygoptera — тремя наружными хвостовыми жабрами и парой верхних анальных придатков (церков)[49]. Поверхность брюшка часто покрыта щетинками, волосками и бугорками. Органы размножения у личинок стрекоз внешне различимы уже на ранних стадиях развития. Самцы практически неотличимы от самок на ранних стадиях развития, позднее у них происходят характерные изменения, наружные половые придатки становятся хорошо развитыми на поздних стадиях развития[50].
Бо́льшую часть жизни личинки стрекоз не могут дышать атмосферным воздухом и для дыхания используют жабры, расположенные на конце тела, которые могут быть внутренними (ректальными) или наружными (каудальными). На брюшке личинок большинства равнокрылых стрекоз (Zygoptera) развиты хвостовые трахейные жабры, которые могут иметь вид листовидных прозрачных пластинок или овальных капсуловидных (пузыревидных) придатков. У личинок разнокрылых стрекоз (Anisoptera) трахейные жабры расположены в специальной камере в прямой кишке (ректальные жабры), которая периодически заполняется водой[50].
Кровеносная система состоит из спинного сосуда, который делится на аорту, переднее и заднее сердца. Переднее сердце состоит из шести камер, располагается в II—VII сегментах брюшка (переходящее во II сегменте в аорту). Заднее сердце состоит из двух камер и находится в VIII и IX сегментах[61].
Развитие и размножение
Жизненный цикл
.jpg.webp)
Стрекозы развиваются с неполным превращением: они проходят стадии яйца, личинки (наяды) и имаго. Многие стрекозы способны производить более одной генерации в год (вольтинизм). Показана обратная связь распространённости вольтинизма с географической широтой: это явление наблюдается чаще ближе к экватору (исключение — семейство лютки)[63]. Спаривание стрекоз происходит в воздухе, при этом процесс у представителей разных подотрядов различается. Самцы разнокрылых стрекоз захватывают самку своими анальными придатками за голову, а самцы равнокрылых удерживают партнёршу за переднегрудь. Совокупительные органы самцов располагаются на II сегменте брюшка (исключительный случай среди насекомых), а семявыводящие протоки на IX сегменте. Предварительно перекачав сперму из девятого во второй абдоминальный сегмент, самцы начинают гоняться за самками. Продолжительность спаривания различается у разных видов и может длиться от нескольких секунд до трёх часов[18].
После спаривания оплодотворённые самки откладывают яйца прямо на воду, на водоросли, прибрежный ил, мох, гниющие растительные остатки (равнокрылые стрекозы и разнокрылые семейства Aeshnidae) либо в специальные углубления в тканях растений[18][64]. При этом самки, окружённые воздушным пузырём (благодаря которому они остаются сухими), могут погружаться в воду, у некоторых групп — вместе с присоединившимся самцом (лютки из подсемейств Lestinae и Agrioninae). Некоторые самки красоток без самцов погружаются глубоко под воду, например, у красотки-девушки — глубже, чем на метр[52][59].
Развитие из неоплодотворённых яиц (партеногенез) для стрекоз нехарактерно и обнаружено в природе только у стрекозы Ischnura hastata (Coenagrionidae) и только в одной из популяций на Азорских островах[24][65]. Другим исключением служит яйцеживорождение, обнаруженное у Heliocypha perforata (Chlorocyphidae) в Западном Китае, когда молодые личинки (предличинки, или пронимфы, вероятно, вылупившиеся из яиц внутри тела матери) откладывались самкой на частично погружённую ветвь дерева[66].


 Coenagrion puella
Coenagrion puella_mating_female_typica_3.jpg.webp)
Яйца
В среднем стрекозы откладывают от 250 до 500 яиц. По характеру откладывания яиц выделяют две группы стрекоз. Экзофилы характеризуются округлой формой яиц. К ним относятся большинство разнокрылых стрекоз, откладывающих яйца прямо в воду или на твёрдые водные объекты. У эндофилов отличаются яйца округлые, но удлинённые. Эндофилами являются равнокрылые стрекозы и коромысла. Они откладывают яйца в рыхлый субстрат или в ткани растений[52]. Одна самка сжатобрюха полосатого за всю свою жизнь (около 3 месяцев) откладывает до 1041 яйца, а у прямобрюха синеющего — до 3081 яйца за 2 месяца[67]. Смертность яиц у стрекоз достаточно высока, например, у огнетелки нимфальной (Pyrrhosoma nymphula) она достигает 25 %[68]. Их смертность повышается при экстремальных температурах, в случае загрязнения среды, наличия паразитов и хищников. Время эмбрионального развития и, следовательно, время вылупления личинок из яиц увеличивается при низких температурах. Кроме того, размер вылупляющихся личинок уменьшается с понижением температуры[69].
При благоприятных условиях личинки отрождаются уже спустя несколько дней, но у большинства видов в умеренной климатической зоне это происходит лишь на следующую весну. Время развития яиц зависит от климатических факторов и длится от нескольких недель у весенних видов до нескольких месяцев у стрекоз, откладывающих их осенью (тогда яйца зимуют). Короткие сроки развития яиц (до 20 дней) характерны для тропических групп и представителей умеренного климата, таких как красотки, стрелки настоящие, плоскобрюхи, белоносы, Cordulia и другие. Для родов с осенним типом развития (лютки, коромысла, стрекозы-каменушки) средний срок развития яиц достигает 9 месяцев[52], в разных группах обычно в пределах от 80 до 230 суток[70].
Личинки
Предличинка (пронимфа) выходит из яйца, имеет крошечные размеры (менее 1 мм) и живёт от нескольких секунд у разнокрылых стрекоз до нескольких минут у равнокрылых, после чего линяет и переходит в следующую личиночную стадию. На первых стадиях у личинок отсутствуют крыловые зачатки (чехлики), лапки нерасчленённые, число сегментов в усиках неполное, отсутствуют глазки, но маска уже развита. В дальнейшем у разных видов развитие личинок проходит через 7—11 линек (9—11 линек у Lestes, 7—8 у Sympetrum). Личиночная стадия длится от 3 месяцев до 4—5 лет[49]. Личинки стрекозы двупятнистой развиваются 35 месяцев, коромысла голубого — 38, а у некоторых дедок из подсемейства Gomphinae — до 47 месяцев[52]. Рекордную продолжительность развития имеют личинки дозорщика-седлоносца — до 6 лет[71]. Сроки развития зависят от наличия кормовых ресурсов и климатических условий[52]. Длительное развитие, как правило, характерно для реофильных видов и видов, развивающимся при низких температурах в северных широтах и высокогорьях[49].

_(Blue_Emperor)_-_(larva_-_nimf)%252C_Buren_(Gld.)%252C_the_Netherlands.jpg.webp)
После первых линек у личинок появляются крыловые чехлики, увеличивающиеся в размерах с течением времени. Сегменты антенн становятся более многочисленными. При каждой линьке увеличивается число ячеек сложных глаз. С ростом личинки также происходят изменения всех органов, в том числе маски, лапок, спинного придатка, хвостовых и ректальных жабр[61].
Личинки стрекоз ведут водный образ жизни. Обычно они живут в медленно текущих или стоячих пресных водоёмах (заросшие пруды, озёра, старицы рек, канавы, болота). Лишь некоторые (стрелки настоящие, ложнокрасотка Фатима, дедки) встречаются в реках с сильным течением. В солоноватых водах живут некоторые представители родов стрелки настоящие, синестрелки, красноглазки, тонкохвосты, плоскобрюхи, прямобрюхи, стрекозы-каменушки. Булавобрюх увенчанный — единственный вид стрекоз, обитающий в горах Средней Азии, преимагинальные фазы которого могут развиваться в горных ручьях с преимущественно ледово-снеговым питанием[72]. Большинство личинок стрекоз живёт на дне или на подводных растениях. Некоторые закапываются в грунт (Gomphinae, Cordulegastridae). Личинки стрелок и настоящих стрекоз способны переносить пересыхание и промерзание водоёма, продолжая своё развитие после нового наполнения его водой[49]. Некоторые личинки способны развиваться в микроводоёмах, таких как заполненные водой углубления в почве. Отдельные виды способны развиваться в сильно минерализованных или загрязнённых водоёмах[49].
Личинки являются малоподвижными и обычно сидят среди подводной растительности или зарываются в грунт, терпеливо поджидая добычу. Плавают не очень хорошо и редко, обычно будучи потревоженными или спасаясь от врагов. Личинки стрекоз характеризуются отрицательным фототаксисом и избегают яркого света[49].
Личинки стрекоз, как и взрослые особи, являются хищниками, питаясь главным образом личинками водных насекомых. Разные подотряды стрекоз специализированы на разных видах корма. Разнокрылые стрекозы (Anisoptera) питаются активно двигающимися личинками водных животных (веснянок, жуков, комаров, подёнок), мальками рыб, а также пиявками, головастиками, водными клопами, рачками-осликами. Равнокрылые стрекозы (Zygoptera) поедают в основном мелких ракообразных (дафний, остракод, циклопов)[61].
- Морфоэкологические типы личинок
По особенностям строения, обитания и поведения выделяют несколько морфоэкологических типов личинок:[49]
1. Широкожаберные личинки, обитающие в стоячих водоёмах. Характерны для многих равнокрылых стрекоз (кроме красоток). Активно плавают, при этом гребут ногами и изгибают тело. Ловят мелкую добычу (личинок насекомых и ракообразных).
2. Длинноногие узкожаберные личинки, обитающие в реках и ручьях. Указан для семейства красотки. Питаются личинками подёнок, прицепляются к водным растениям.
3. Длиннобрюхие и активно плавающие личинки, обитающие в стоячих водах. Это виды семейства коромысла, передвигающиеся реактивным способом, выталкивая воду из прямой кишки.
4. Волосатые личинки, зарывающиеся в грунт. Характерен для семейства дедки. Активные хищники, поедающие крупную добычу, включая мальков рыб и головастиков.
5. Короткобрюхие ползающие личинки, обитающие в стоячих водоёмах. Отмечен в семействах бабки и настоящие стрекозы.
Имаго

К моменту превращения личинок во взрослых стрекоз личинки начинают дышать атмосферным воздухом. Для этого они по любой опоре (стебли водных растений, камни, коряги, грунт и прочие надводные объекты) наполовину вылезают из воды и затем остаются в этом положении от нескольких часов до нескольких дней. Для окончательного превращения личинки полностью выбираются из воды, сидят крепко, держась ногами за опору, некоторое время отдыхают и после отдыха начинают постепенно освобождаться от шкурки. Оболочка личинки разрывается, и из неё выходит взрослая стрекоза. Для приобретения нормальной окраски и окончательного формирования покровов у разных видов и в разных условиях окружающей среды требуется от нескольких часов до суток[52].
Особенности физиологии
Полёт
.JPG.webp)
Основной формой передвижения стрекоз является полёт, помогающий им искать пищу и полового партнёра для размножения, расселяться и мигрировать, спасаться от хищников. Разнокрылые стрекозы характеризуются быстрым резким полётом, способностью отлетать на далёкие расстояния от водоёмов и спариваться вдали от водоёма. Равнокрылые — медленно порхающие стрекозы, не отлетающие далеко от водоёмов и спаривающиеся всегда около воды[50].
При полёте стрекозы машут передней и задней парами крыльев поочерёдно, добиваясь лучшей манёвренности, или одновременно — для большей скорости. Скорость полёта некоторых крупных разнокрылых стрекоз достигает 15 м/с[73] (36—55 км/ч)[74]. Стрекоза Austrophlebia costalis способна летать со скоростью до 97 км/ч[75][76].
Питание и охота

Имаго стрекоз — единственные беспозвоночные, которые благодаря своей массовости и лётным способностям практически безраздельно освоили нишу воздушных хищников. Особенностью стрекоз является отсутствие пищевой специализации: они питаются буквально всеми доступными им насекомыми. Представители подсемейства Pseudostigmatinae являются единственным среди стрекоз примером пищевой специализации, они питаются исключительно пауками-кругопрядами, которых хватают прямо с их ловчих сетей[77].
Взрослые стрекозы — активные хищники воздушной среды. Наличие индивидуальных охотничьих участков отмечено у представителей семейств коромысла (Aeshnidae) и бабки (Corduliidae), видов рода настоящие стрекозы (Libellula) и даже у некоторых мелких стрекоз подотряда равнокрылые стрекозы (Zygoptera)[49].
У стрекоз выделяются несколько групп по способам охоты[49]:
- Свободные охотники верхнего яруса — виды с развитыми крыльями, обитающие на открытых пространствах, часто вдали от водоёмов. Охотятся на высоте 2—10 м. Каждая стрекоза контролирует индивидуальный охотничий участок, иногда даже несколько. При наличии большого количества добычи у них может наблюдаться «роевая охота», когда стрекозы собираются в подобие охотничьих стай численностью до 20 особей. К этой группе относятся представители семейств булавобрюхи, коромысла и других.
- Свободные хищники среднего яруса — виды, которые охотятся главным образом над поверхностью воды водоёмов на высоте 0,5—2 м. Такие стрекозы практически постоянно находятся в полёте, изредка присаживаясь на прибрежные или торчащие из воды растения. К этой группе относится большинство представителей семейств дедки и бабки.
- Подстерегающие хищники среднего яруса — стрекозы, большую часть времени охоты проводящие на «наблюдательных пунктах» (верхушках трав и кустарников, торчащих из воды ветках и стеблях растений) и время от времени срывающиеся в погоню за пролетающими насекомыми. К этой группе относятся представители семейства настоящие стрекозы.
- Пасущиеся охотники нижнего яруса — стрекозы, охотящиеся в прибрежном травостое, медленно перепархивая с растения на растение в поисках сидящих на них мелких насекомых. Замеченную жертву стрекоза хватает в резком броске и затем поедает, усевшись на какое-либо растение. Такой способ охоты свойствен всем представителям подотряда равнокрылых стрекоз.
Весьма широко у стрекоз развит каннибализм — поедание более мелких стрекоз крупными представителями других видов, особенно широко это явление распространено у крупных стрекоз[78][79]. Известны случаи сексуального каннибализма, когда самки (например, стрелки Ischnura graellsii) поедают самцов во время копуляции[80].
Особенности биологии и экологии

Жизнь имаго стрекоз можно разделить на два периода. Ювенильный период начинается после линьки и выхода в воздух и заканчивается половым созреванием. На протяжении этого периода стрекозы активно питаются на определённом удалении от водоёмов и приобретают характерную для имаго окраску тела. По окончании ювенильного периода они вновь возвращаются к водоёмам. Во время репродуктивного периода стрекозы спариваются вблизи водоёмов, самки откладывают яйца[64].
Большие отличия существуют между видами по степени привязанности к водоёмам. Так, представители рода красоток держатся исключительно в непосредственной близости от воды, другие же виды способны разлетаться на далёкие расстояния от водоёмов, например, в пустыни или населённые пункты. При этом все виды характеризуются приуроченностью к свойственным им водоёмам, к которым они прилетают для размножения[50].
Характерной особенностью крупных стрекоз (Anax, Aeshna, Orthetrum, Crocothemis, Sympetrum, Libellula и др.) является совершение «патрулирующих» полётов. У них отмечается хорошо выраженное территориальное поведение, заключающееся в наличии индивидуальных кормовых участков[81]. Мелкие виды обычно охотятся из засады, что особенно выражено, например, у представителей родов Lestes, Sympecma, Coenagrion, Erythromma, Ischnura, Enallagma и других. Если охота не приносит результатов, они перелетают на новое место. У видов с подобными особенностями трофической активности индивидуальные кормовые участки не выявлены. Этим объясняются скопление особей на территории кормления, вблизи водоёмов, и единичность особей на значительном удалении от них[81]. Таким образом, разнокрылые стрекозы преимущественно ловят насекомых во время полёта в воздухе, а равнокрылые часто собирают их с листьев и стеблей травянистых растений и кустарников.

Активность и продолжительность жизни

В умеренных широтах стрекозы встречаются начиная со второй половины апреля и вплоть до октября, в тропиках же они встречаются круглогодично. У стрекоз прослеживается чёткая тенденция к удлинению с севера на юг длительности времени лёта видов. Например, у представителей рода стрекозы-каменушки в центральной части умеренной климатической зоны лёт длится четыре месяца, а на юге этой же зоны — шесть месяцев; в субтропическом климате он растягивается до восьми и более месяцев, а при благоприятных погодных условиях может продолжаться круглогодично за счёт перманентного выплода имаго и развития второго поколения за год. Помимо этого, причиной удлинения лёта является появление на юге видов с весьма длительным периодом жизни имаго, что происходит благодаря нарушениям синхронизации жизненных циклов и сильно растянутому выплоду стрекоз из личинок, а также изменениям периодизации жизненных циклов взрослых стрекоз. На севере циклы развития гораздо более синхронизированны за счёт сезонных изменений температур и связанного с ними комплекса фенологических явлений[81].
Стрекозы активны в дневное время в тёплую солнечную погоду. В утренние часы и при низких температурах стрекозы часто сидят, греясь под лучами солнца, на камнях, коре деревьев и прочих субстратах, которые способны к быстрому теплообмену[18]. При высоких температурах в дневное время, чтобы избежать перегрева, настоящие стрекозы и дедки, сидя, принимают характерное положение «обелиска» — ориентируют кончик брюшка вверх к солнцу, снижая воздействие тепла на тело[82][83].
На протяжении дня поведение стрекоз изменяется: так, у большинства видов утром и вечером преобладает охотничья активность[84], а в дневные часы — поведение, связанное с размножением. Некоторые виды, преимущественно представители семейства коромысел, характеризуются поздними вечерними пиками охотничьей активности, кормясь пойманными насекомыми из скоплений двукрылых до наступления темноты[84][18].
Продолжительность жизни имаго в среднем составляет от нескольких дней до нескольких недель. Отдельные экземпляры, особенно разнокрылых стрекоз, могут жить до нескольких месяцев[18], например, лютка-невеста и дозорщик-император — около 60 дней, а дедка обыкновенный — 26 дней. Виды, которые зимуют на стадии имаго, могут прожить до 6 месяцев (лютка рыжая). Однако наблюдения показывают, что взрослая жизнь стрекоз чаще всего заканчивается преждевременно, так как они становятся жертвами различных хищников[71].
Отношение к водоёмам
Стрекозы в целом проявляют исключительную пластичность в выборе водоёмов для развития своих личинок: для них подходящими являются все водоёмы, начиная от горных потоков и ручьёв, прудов, озёр, стариц рек, ручьёв и заканчивая солоноватыми и торфяными водоёмами, вплоть до созданных человеком прудов, оросительных каналов и канав. Примечательно, что в таком большом озере, как Байкал, стрекозы отсутствуют[85]. Представителям подсемейства Pseudostigmatinae достаточно различных малых природных водных резервуаров среди подлеска — они используют затопленные норы и дупла, участки со скапливающейся и застаивающейся водой, заполненные водой розетки листьев эпифитов[86].
В то же время личинки отдельных видов и родов, а порой и семейств, приспособлены к обитанию только в водоёмах определённого типа. Так, имеются целые семейства, способные жить только в проточной воде; другие же виды приспособлены к обитанию в быстрых потоках и совершенно не переносят стоячей и болотистой воды. Существуют стрекозы, личинки которых обитают в сильно минерализованных водоёмах — солоноватых или щелочных водах — как в нормальной для них среде обитания. Однако по существу они лишь обладают определённой толерантностью к минерализации воды, а нормально живут в относительно чистой воде. Для большинства же видов минерализованная вода малопригодна для обитания[52]. Прямобрюх белохвостый на севере ареала часто населяет горячие источники с температурой не менее 35—40 °C, где личинки развиваются в мелководных разливах остывающей термальной воды[87].
Бо́льшая часть видов стрекоз не придерживается строго определённого типа водоёмов. Виды, чьи личинки обитают в проточных водах, более требовательны в выборе водоёмов, так как нуждаются в большом количестве кислорода и не могут жить в стоячих водах[85].
Некоторые тропические стрекозы, например, Mecistogaster modesta и другие представители семейства Pseudostigmatidae, развиваются в небольших скоплениях воды (микроводоёмах), образовавшихся в листьях бромелиевых и разных эпифитных растений. В тропических лесах северо-западной Коста-Рики в Центральной Америке они достигают высокой плотности: до 6 тысяч личинок на гектар в участках вторичного леса[88].
Миграции
Как и многие другие насекомые, стрекозы могут совершать миграции, во время которых могут преодолевать океаны и высокие горные хребты, летая ночью и на больших высотах[89][90][91]. Миграции с севера на юг характерны для Anax junius в Северной Америке; некоторые мигранты перелетали на расстояние до 2800 км с северной на южную части континента[92]. Стрекозы всемирно распространённого рода бродяжки используют пассивное распространение, чтобы пересечь Индийский океан[90], при этом поддерживая местные островные популяции, например, в Тихом океане на острове Пасхи[57]. Адаптивная функция миграции заключается в перемещении отдельных особей и популяций из ухудшающейся со временем среды обитания в более благоприятную на текущий момент[93].
Численность мигрирующих стрекоз порой достигает гигантских количеств. Например, в 1991 году в Аргентине была отмечена гигантская стая, предположительно, Aeshna bonariensis, численностью примерно в 4—6 млрд особей и общей массой около 4 тысяч тонн[94]. Согласно письменным свидетельствам, в XIX веке через Антверпен пролетала большая стая Libellula quadrimaculata. В Северном Казахстане подобная стая стрекоз была оценена в 100 млн особей[95]. Однако подобные массовые миграции отмечаются редко, как правило, перелёты осуществляются разрозненными особями, а не стаями[89].
Для разных видов стрекоз характерны различные типы миграций. Выделяют два типа миграций — нерегулярные и регулярные. В первом случае стрекозы совершают миграции лишь в отдельные годы, могут образовывать гигантские стаи. При втором типе миграции являются неотъемлемой частью жизненного цикла видов и совершаются ими ежегодно[89].
К нерегулярному типу миграций можно отнести массовые перелёты ряда видов — например, массовое размножение коромысел зеленобоких на юге Кыргызстана в 2009 году либо коромысел помесных и симпетрумов южных в 2006 году в дельте Дуная[96]. Подобный тип миграций, вероятно, вызван необходимостью массового переселения стрекоз из мест обитания при чрезмерном росте плотности популяций[95].
Второй тип миграции характерен для группы стрекоз Северного полушария и представлен широтными миграциями разных поколений из одной природно-климатической зоны в другую[89][97]. Выделяют два варианта регулярных миграций. Первый — сезонные трансширотные миграции — из районов выплода стрекоз в новые места обитания. Весной стрекозы прилетают из южных частей ареалов (вероятно, тропических и субтропических регионов) в умеренные широты, где дают начало новому поколению. Осенью особи летнего поколения мигрируют обратно на юг, где размножаются[82][97]. Протяжённость таких миграций составляет сотни и тысячи километров. Данный тип миграций характерен для немногих видов — например, бродяжки рыжей, дозорщика-седлоносца, сжатобрюха Фонколома, Anax junius и других[89]. Второй — сезонные межстациальные миграции, в ходе которых стрекозы перемещаются из районов выплода (репродуктивные стации) в кормовые и обратно[82]. Они обусловлены сезонным пересыханием водоёмов или иными временными ухудшениями условий в местах обитания. Протяжённость этих миграций составляет от сотен метров до десятков или сотен километров. Они являются типичными для Sympetrum, Sympecma, Selysiothemis nigra и некоторых других стрекоз[89][97].
Роль в экосистемах
Будучи амфибионтными насекомыми и обладая высокой численностью, стрекозы являются важным компонентом как водных, так и наземных экосистем. Личинки стрекоз составляют до 20 % пищевого рациона некоторых видов рыб, а у некоторых ныряющих видов водоплавающих птиц — до половины рациона[49].
Враги и паразиты

Среди паразитов стрекоз отмечены споровики-грегарины, черви-сосальщики трематоды, круглые черви филярии, водяные клещи (Hydrachnidae, Prostigmata), некоторые наездники[52].
Основные враги личинок стрекоз — рыбы и водяные птицы, которые ими питаются. Хищные и всеядные рыбы поедают личинок в большом количестве, в то время как мелкие рыбки могут поедать яйца стрекоз. Личинками питаются птицы, обитающие на отмелях, заросших камышом и водной растительностью, по берегам различных водоёмов (кулики, крачки, цапли и другие). На личинок стрекоз нападают некоторые водные клопы (Ranatra, Notonecta), жуки-плавунцы (Dytiscus) и их личинки. Мелких личинок могут улавливать пресноводные гидры. Взрослые стрекозы попадаются в сети пауков-кругопрядов, плетущих их около водоёмов, пауков-бродяг (Dolomedes), пауков-тетрагнатид (Tetragnatha). Взрослых стрекоз ловят крупные хищные мухи-ктыри[52]. Стрекозы отмечены в питании крачек, чаек, уток и других околоводных птиц, некоторых воробьинообразных птиц, чёрных стрижей, большого пёстрого дятла[98]. Среди птиц выделяется своей специализацией золотистая щурка, а также белая трясогузка, которая даже устраивает на побережьях «кухонные, или кормовые, столики», куда переносит добычу и где скапливаются обломки крыльев стрекоз[98]. Враги стрекоз не отмечены среди пресмыкающихся и млекопитающих[52].
Также стрекозы могут становиться случайной добычей насекомоядных растений, таких как росянки (Drosera), которые растут на торфяных и сфагновых болотах[99].
Взаимодействие с человеком
Хозяйственное значение
Как и другие хищники, стрекозы играют двоякую роль. С одной стороны, они активные истребители личинок комаров, а эти личинки служат пищей для рыб и некоторых околоводных птиц. Однако некоторые виды поедают также мальков хозяйственно важных видов рыб. Личинки таких крупных, включённых в Красные книги стрекоз, как коромысло большое (Aeshna grandis) и дозорщик-император (Anax imperator), наносят некоторый вред рыбному хозяйству, поедая мальков карпа и линя[61].
Личинки ряда видов являются переносчиками простогонимоза птиц, выступая в роли дополнительных хозяев паразита. Основными возбудителями заболевания являются трематоды Prosthogonimus pellucidus, P. cuneatus и P. ovatus из семейства Prosthogonimidae. Из домашних птиц часто поражаются куры, индейки, утки и гуси; часто эти трематоды обнаруживаются у диких птиц (куропаток, глухарей, фазанов, водоплавающих и болотных птиц). Вместе с экскрементами больных птиц яйца попадают в воду, где заглатываются пресноводными моллюсками. В их организме из яиц выходят мирацидии, превращающиеся в спороцисты, редии и церкарии. Церкарии покидают моллюсков и затем, попадая в организм личинок стрекоз, превращаются в метацеркариев, которые являются жизнеспособными и после превращения личинок во взрослых насекомых. Птицы заражаются при поедании заражённых личинок стрекоз. Свободные метацеркарии обычно локализуются в полости брюшка (в его задней части), а инцистированные — в мышцах груди и брюшка, на стенках пищевода или в жировом теле взрослых стрекоз[100].
Изучение стрекоз
Специалисты по стрекозам, или одонатологи, исследуют их как в лабораторных, так и в полевых условиях. В области одонатологии на рубеже XX и XXI веков работало более 700 исследователей более чем из 70 стран мира[101]. В 1971 году для координации работы одонатологов на Первом европейском симпозиуме было учреждено Международное общество одонатологов (The Societas Internationalis Odonatologica, S.I.O.) со штаб-квартирой при Утрехтском университете в Нидерландах, которое с 1972 года начало издавать ежеквартальный журнал «Odonatologica»[102]. В 1957 году в Японии было основано одно из старейших обществ одонатологов (日本トンボ学会, The Japanese Society for Odonatology)[103]. В Германии с 1981 года издаётся журнал «Libellula», а в 1982 году в Бонне образовано Общество немецкоязычных одонатологов (Gesellschaft deutschsprachiger Odonatologen e.V., GdO)[104]. Крупная одонатологическая научная школа была создана в России. Благодаря трудам видных её представителей, таких как А. Н. Бартенев (1882—1946), Б. Ф. Белышев (1910—1993), А. Ю. Харитонов, были вскрыты многие принципиальные особенности пространственного и зоогеографического распределения стрекоз[101]. В 1986 году в Новосибирске был проведён Первый Всесоюзный симпозиум одонатологов. В 1998 году создано Российское отделение S.I.O. По данным реферативного раздела журнала «Odonatologica», с 1971 по 2005 год в мировой научной литературе опубликовано свыше 14 000 публикаций по стрекозам[105][106]. В 1988 году одонатологи США, Канады и Латинской Америки объединились в Dragonfly Society of the Americas (DSA), которое издаёт два журнала: «Argia» (News Journal of the DSA) и «Bulletin of American Odonatology»[107].
С 2010 года в проводятся Европейские одонатологические конгрессы: 1-й (1st European Congress on Odonatology, 2010, Вила-ду-Конди, Португалия), 2-й (2012, Белград, Сербия), 4-й (2016, Тюринге, Швеция)[108]. Существует Всемирная ассоциация исследователей стрекоз (The Worldwide Dragonfly Association, WDA), созданная как для научных целей, так и для освещения вопросов сохранения редких видов[109]. Ассоциация публикует три специализированных журнала: «International Journal of Odonatology» (2004), «Agrion» (1989) и «Odonatological Abstracts»[110]. WDA проводит Международные одонатологические конгрессы (International Congress on Odonatology): 2012 (Odawara, Япония), 2013 (Freising, Германия), 2015 (La Plata, Аргентина), 2017 (Clare College, Кембридж, Великобритания)[111]. Кроме того, Всемирная ассоциация поддерживает систему грантов для поддержки одонатологических исследовательских проектов[112].
Охрана
.jpg.webp)
Стрекозы являются важной индикаторной группой, остро и быстро реагирующей как на любые изменения качества воды в водоёмах, где обитают личинки, так и на изменения структуры самого биотопа[114]. Целый ряд факторов может негативно влиять на численность популяций стрекоз, вызывать их угнетение и исчезновение. В настоящее время главной угрозой сокращения численности многих видов является хозяйственная деятельность человека. Вызванное ей глобальное уничтожение, сокращение и загрязнение естественных мест обитания привело к тому, что целый ряд видов стрекоз занесён в национальные Красные книги[18]. В Красную книгу России занесена стрекоза дозорщик-император (категория 2 — сокращающийся в численности вид)[115], 9 видов занесены в Красную книгу Украины[116], 8 видов — в Красную книгу Белоруссии[117], 13 видов — в Красную книгу Литвы[118], 3 вида — в Красную книгу Молдавии[119], 7 видов — в Красную книгу Казахстана[120], 1 вид — в Красную книгу Узбекистана[11].
В Международную Красную книгу (Красный список угрожаемых видов МСОП, The IUCN Red List of Threatened Species) по состоянию на август 2018 года занесено 308 редких, исчезающих и уже вымерших видов и 26 подвидов стрекоз (с охранными статусами Vulnerable, Endangered, Critically Endangered, Extinct species), из них 1 вид — вымерший, 59 видов и 2 подвида — находящиеся на грани исчезновения, 121 вид и 19 подвидов — вымирающие, 127 видов и 5 подвидов — уязвимые. Всего на сайте МСОП перечислено 3643 вида (более половины всех известных видов) и 30 подвидов стрекоз с учётом тех, информации о которых недостаточно (Data Deficient, 907 видов), которые близки к уязвимому положению (Near Threatened, 140 видов) или чьё положение под наименьшей угрозой (Least Concern, 2288 видов и 4 подвида)[121][122]. Вид Megalagrion jugorum с Гавайских островов значится как уже полностью вымерший[123].
В Европе 15 % из 138 видов стрекоз близки к уязвимому положению, ещё около четверти видов (33) снизили свою численность и размер ареал. В Японии 57 из 200 видов Odonata сокращаются в численности, 24 вида уязвимы и 19 находятся под угрозой исчезновения. В Южной Африке из 155 видов стрекоз 13 сокращаются в численности, а 4 вида считаются вымершими в современное время. В целом по данным МСОП около 10 % видов стрекоз мировой фауны находятся на грани исчезновения[124].
Стрекозы в культуре

Одно из самых первых упоминаний стрекоз относится к XVIII—XVII векам до н. э. В клинописном «Эпосе о Гильгамеше», написанном на аккадском языке на основании шумерских сказаний, есть упоминание этих насекомых как пример невозможности бессмертия, чтобы проиллюстрировать краткость человеческой жизни[125].
В древнейшей японской книге, «Кодзики» (VIII век), образ стрекозы синонимичен образу Японских островов. Одно из древних названий острова Хонсю — Акицусима — дословно переводится как «Остров-стрекоза». Наиболее ранние изображения стрекоз встречаются на ритуальных колоколах дотаку, изготовленных в период Яёй. В эпоху Ямато при императорском дворе стрекоза была символом власти. В период Сражающихся царств (1467—1560) стрекозы были символом воинской отваги. Их изображали на одежде и самурайских мечах, а самих стрекоз называли «катимуси» («победители»). Стрекозам посвящено много народных японских сказок. Они также являются одним из распространённых символов в хайку[126]. Согласно «Японской энциклопедии Коданся», стрекоза считалась в мифологии духом риса и предвестником богатых урожаев[127]. В различных городах Японии с 1988 года функционируют парки стрекоз[128].
В VIII веке о стрекозах писал один из крупнейших китайских поэтов Ду Фу (712—770): «Догоняют стрекозы/ Друг друга,/ Изумрудные крылья/ Колебля,/ И блаженствуют/ Лотосы Юга,/ Обнимаясь/ На сдвоенном стебле» (767)[129].

В Европе отношение к стрекозам было неоднозначным. На английском языке стрекоза значится как «dragonfly», что дословно переводится как «драконья муха». В верованиях народов Европы стрекозы часто связаны с нечистой силой и могут выступают как «ездовые животные чёрта»[130]. В эпоху Средневековья стрекоз нередко ассоциировали с ведьмами. В Уэльсе на валлийском языке стрекоза (gwas-y-neidr) в дословном переводе означает «слуга гадюки»[82]. В Швеции существует миф о том, что дьявол использует стрекоз для взвешивания людских душ[131].
На юге США есть фольклорное понятие «змеиный врач» («snake doctor»), согласно которому стрекозы следуют за ранеными змеями и лечат их[132]. Для некоторых аборигенных индейских племён Северной Америки стрекозы представляют собой стремительность и активность; для навахо они символизируют чистую воду. Стрекозы являются общим мотивом в керамике народности зуни; в стилизованной форме креста они появляются в наскальном искусстве индейцев хопи и на ожерельях пуэбло[131].
Образ стрекоз прослеживается на протяжении всей мировой художественной культуры в самых различных произведениях искусства. Предвестниками использования образа стрекозы в декоративно-прикладном искусстве стали живопись и графика Японии, а затем инро — коробочки для хранения мелких предметов[133]. Эти и многие другие предметы японского искусства XVI—XIX века часто гравировались и украшались изображениями растений и насекомых, в том числе стрекоз[134]. В Викторианскую эпоху насекомые вновь стали очень популярным мотивом, и стрекозы не стали исключением. Со временем использование образа стрекоз в искусстве возрастало. В конце XIX века эстетика стрекоз становится одним из ведущих мотивов стиля модерн. В качестве примера можно привести исполнение номера «Стрекоза» Анны Павловой на музыку Фрица Крейслера[135].
Бродят в траве золотые букашки.
Вся голубая, как бирюза,
Села, качаясь, на венчик ромашки,
Словно цветной самолет, стрекоза.
Стрекозы быстрыми кругами
Тревожат черный блеск пруда,
И вздрагивает, тростниками
Чуть окаймленная, вода.
На прозрачном ручейке
Стрекоза-красотка пляшет,
Прихотлива и легка,
Бойко крылышками машет.
Многие поэты — Б. Гёте, А. К. Толстой, О. Мандельштам, Г. Гейне и другие — вдохновляясь красотой и изяществом этих насекомых, писали о них стихи[136][137]. Крупнейший представитель жанра хайку японский поэт Мацуо Басё (1644—1694) написал несколько трёхстиший, посвящённых четырёхкрылым летунам, например: «Всё кружится стрекоза…/Никак зацепиться не может/За стебли гибкой травы» и «О стрекоза!/С каким же трудом на былинке/ты примостилась!»[138]

В русскую культуру стрекоза вошла, прежде всего, как героиня басни Лафонтена «La Cigale et la Fourmi», хотя во французском оригинале и в наиболее распространённом варианте греческой первоосновы она была цикадой. Самое известное переложение «Стрекоза и муравей» сделал в начале XIX века Крылов, хотя и до него существовали близкие по содержанию произведения таких авторов, как Сумароков, Хемницер или Нелединский-Мелецкий[139]. Исключением тут служит В. А. Озеров: герой его басни, написанной в 1797 году, — куда более похожий на цикаду кузнечик[139]. В ранних, семнадцатого века, изложениях исходной басни Эзопа, например у Гозвинского, цикаду также называют сверчком или коником (кузнечиком)[139]. Крыловская стрекоза во многом сохраняет признаки цикад, она «попрыгунья», умеет петь, живёт в траве[139]. Другой басенный сюжет, восходящий к Эзопу, изложил в 1773 году М. Н. Муравьёв. В оригинале говорится о бедном крестьянине, питавшемся саранчой. Когда он поймал цикаду, та взмолилась не есть её, поскольку она не губит урожай, а лишь услаждает людской слух пением. В переложении (предшествовавшем самым известным переводам Лафонтена и считающемся первой из историй о «Стрекозе и Муравье») цикада «превратилась» в стрекозу, а саранча — в кузнечика[139]. Такая путаница характерна только для языка поэзии, в науке соответствующего периода стрекоз с кузнечиками никто не смешивал, и первое название закрепляется именно за теми насекомыми, которых мы знаем под этим именем[139], хотя в различных диалектах русского языка, откуда это слово попало в литературный язык, так могли называть и стрекоз, и кузнечиков, причём исходным, возможно, было второе значение, в силу очевидной этимологической связи слова стрекоза с глаголом «стрекотать»[139].
Полька-мазурка «Стрекоза» (Die Libelle) была написана австрийским композитором Йозефом Штраусом в 1866 году после посещения озера Траунзе, где в тот момент летали массы этих околоводных насекомых, которые и вдохновили музыканта[140][141]. Стрекозе посвящены песни «Белая стрекоза любви» (Quest Pistols)[142] и «Стрекоза» («Нэнси»)[143].
В IV веке в Древнем Китае во времена учёного Гэ Хуна (283—343) впервые упоминается игрушка «бамбуковая стрекоза» (чжу цин тин). Она или использовалась самостоятельно в виде пропеллера на оси (который раскручивался намотанной на него нитью, шнуром или ладонями рук), или прикреплялась к «воздушному змею» для его запуска в воздух[144]. Британский производитель мотоциклов фирма Douglas, базирующаяся в Бристоле, назвала свою новаторскую конструкцию послевоенной модели мотоцикла с двухцилиндровым двигателем «Стрекозой» (Douglas Dragonfly)[145].

Стрекозы являются популярными объектами и в современной культуре. Их фотографии и изображения в той или иной интерпретации часто появляются как на обложках научных книг, посвящённых природе, так и художественных произведений; на рекламных плакатах, на обложках музыкальных CD-дисков, на керамических и ювелирных изделиях[127]. Они также использовались в качестве декоративного мотива на тканях и предметах домашнего обихода[146].
Примечания
- Фасмер М. Этимологический словарь русского языка. — Прогресс. — М., 1964—1973. — Т. 3. — С. 774.
- Черных П. Я. Историко-этимологический словарь современного русского языка. — М.: Русский язык, 1993. — Т. 2. — С. 207—208.
- Mickel Clarence E. The significance of the dragonfly name «Odonata» (англ.) // Annals of the Entomological Society of America : Журнал. — 1934. — Vol. 27, no. 3. — P. 411—414. — ISSN 0013-8746. — doi:10.1093/aesa/27.3.411.
- Jill Silsby. Dragonflies of the World. — Plymouth: The National History Museum, 2001. — P. 180. — ISBN 0-565-09165-4.
- Zhang Z.-Q. «Phylum Athropoda». — In: Zhang Z.-Q. (Ed.) «Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013)» (англ.) // Zootaxa / Zhang Z.-Q. (Chief Editor & Founder). — Auckland: Magnolia Press, 2013. — Vol. 3703, no. 1. — P. 17—26. — ISBN 978-1-77557-248-0 (paperback) ISBN 978-1-77557-249-7 (online edition). — ISSN 1175-5326. Архивировано 19 сентября 2013 года.
- Zhang Z.-Q. et al. «Phylum Arthropoda von Siebold, 1848». — In: Zhang Z.-Q. (Ed.) «Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness» (англ.) // Zootaxa / Zhang Z.-Q. (Chief Editor & Founder). — Auckland: Magnolia Press, 2011. — Vol. 3148. — P. 99—103. — ISBN 978-1-86977-849-1 (paperback) ISBN 978-1-86977-850-7 (online edition). Архивировано 17 апреля 2016 года.
- Resh V., Carde R. Encyclopedia of Insects, Second Edition. — Elsevier Academic Press, 2009. — P. 721—728. — 1168 p. — ISBN 978-0123741448.
- Белышев Б. Ф., Харитонов А. Ю., Борисов С. Н. и др. Фауна и экология стрекоз. — Новосибирск: Наука. Сиб. отд-ние, 1989. — С. 33, 44—48. — 207 с.
- James William Tutt. The Entomologist's Record and Journal of Variation (нем.) : magazin. — Charles Phipps, 1997. — S. 213.
- Kalkman V. J., Lopau W. Identification of Pyrrhosoma elisabethae with notes on its distribution and habitat (Odonata: Coenagrionidae) (англ.) // International Journal of Odonatology. — 2006. — No. 9. — P. 175—184.
- Красная книга Республики Узбекистан. Том 2. Животные / гл. ред. Ж. А. Азимов. — Ташкент: Chinor ENK, 2009. — 218 с.
- Foottit R. G., Adler P. H. Insect Biodiversity: science and society. — Blackwell Publishing Ltd, 2009. — P. 31. — 642 p. — ISBN 978-1-4051-5142-9.
- Dumont H. J. Relict distribution patterns of aquatic animals: another tool in evaluating late pleistocene climate changes in the Sahara and Sahel // Palaeoecology of Africa and the Surrounding Islands. — 1982. — № 14. — P. 1—24.
- Suhlin F., Jödick R., Schneide W. Odonata of African arid regions — are there desert species // Cimbebasia. — 2003. — № 18. — P. 207—222. Архивировано 13 августа 2018 года.
- Стрекозы (Odonata) России (недоступная ссылка). Стрекозы РФ. Дата обращения: 28 июня 2017. Архивировано 23 апреля 2017 года.
- Чаплина И. А. Одонатологические исследования в Казахстане // Зоологические исследования в Казахстане. — Алма-Ата, 2002. — С. 266—267.
- Чаплина И. А. К фауне стрекоз Казахстана // Биологическая наука и образование в педагогических вузах. — Новосибирск, 2003. — Вып. 3. — С. 1923.
- Горб С. Н., Павлюк Р. С., Спурис З. Д. Стрекозы (Odonata) Украины: фаунистический обзор (укр.) = Бабки (Odonata) України: фауністичний огляд // Вестник зоологии. — К., 2000. — Т. Отдельный выпуск 15. — С. 1—155. (укр.)
- Савчук В. В., Каролинский Е. А. Новые находки редких видов стрекоз (Insecta: Odonata) на Украине // Вестник зоологии. — 2013. — Т. 47, № 6. — С. 506.
- Buczynski P., Dijkstra D-B., Mauersberger R., Moroz V. D. Review of the Odonata of Belarus (англ.) // Odonatologica. — 2006. — Vol. 35, no. 1. — P. 1—13.
- На Ольманских болотах нашли новый для Беларуси вид стрекоз (рус.) ?. Telegraf.by (4 августа 2021). Дата обращения: 4 августа 2021.
- Miguel T. B., Calvão L/ B., Vital M. V. C., Juen L. A scientometric study of the order Odonata with special attention to Brazil (англ.) // International Journal of Odonatology. — Worldwide Dragonfly Association, Taylor & Francis, 2017. — Vol. 20, no. 1. — P. 27—42. — ISSN 2159-6719. Архивировано 2 августа 2017 года.
- Sánchez-Herrera M., Ware J. L. Biogeography of Dragonflies and Damselflies: Highly Mobile Predators // Global Advances in Biogeography / Dr. Lawrence Stevens (Ed.). — InTech, 2012. — P. 291—306. — 360 p. — ISBN ISBN 978-953-51-0454-4. Архивированная копия (недоступная ссылка). Дата обращения: 7 июля 2017. Архивировано 13 мая 2018 года.
- Suhling F., Sahlén G., Gorb S., Kalkman V. J., Dijkstra K-D. B., van Tol J. Order Odonata // Ecology and general biology. Thorp and Covich's Freshwater Invertebrates / Thorp J., Rogers C. D.. — 4. — Academic Press, 2015. — P. 893—932. — ISBN 978-0-12-385026-3.
- Carle F. L. A new Epiophlebia (Odonata: Epiophlebioidea) from China with a review of epiophlebian taxonomy, life history, and biogeography // Arthropod Systematics & Phylogeny. — 2012. — Vol. 70, № 2. — P. 75—83.
- Историческое развитие класса насекомых / Ред. Родендорф Б. Б., Расницын А. П. — М.: Наука, 1980. — Т. 175. — С. 127—133. — 256 с. — (Труды ПИН).
- Martynov A. V. New Permian Palaeoptera with the discussion of some problems of their evolution // Труды Палеозоологического института АН СССР. — 1932. — Т. 1. — С. 1—44.
- Мартынов А. В. Очерки геологической истории и филогении отрядов насекомых (Pterygota). Ч. 1. Palaeoptera и Neoptera-Polyneoptera // Труды ПИН АН СССР. — М.—Л., 1938. — Т. 7, вып. 1. — С. 1—150.
- Grimaldi D. A., Engel M. S. Evolution of the Insects. — Cambridge University Press, 2005. — P. 173—187. — 763 p. — ISBN 0-521-82149-5. Архивированная копия (недоступная ссылка). Дата обращения: 4 октября 2017. Архивировано 13 мая 2018 года.
- Maddison, David R. Pterygota. Winged insects (англ.). Tree of Life Web Project (1 января 2002). Дата обращения: 6 июля 2017. Архивировано 10 июня 2017 года.
- Trueman, John W.H. Pterygote Higher Relationships (англ.). Tree of Life Web Project. Дата обращения: 6 июля 2017. Архивировано 15 июля 2017 года.
- Bechly G., Nel A., Martínez−Delclòs X., Jarzembowski E.A., Coram R., Martill D., Fleck G., Escuillié F., Wisshak M. M., Maisch M. Revision and phylogenetic study of Mesozoic Aeshnoptera, with description of numerous new taxa (Insecta: Odonata: Anisoptera) // Neue Paläontologische Abhandlungen. — CPress, 2001. — Vol. 4. — P. 1—230.
- Rehn A. C. Phylogenetic analysis of higher-level relationships of Odonata (англ.) // Systematic Entomology. — Wiley-Blackwell, 2003. — Vol. 28, no. 2. — P. 181—240. — doi:10.1046/j.1365-3113.2003.00210.x.
- Dijkstra K. D. B., Bechly G., Bybee S. M., Dow R. A., Dumont H. J., et al. The classification and diversity of dragonflies and damselflies (Odonata) (англ.) // Zootaxa. — 2013. — Vol. 3703, no. 1. — P. 36—45. — doi:10.11646/zootaxa.3703.1.9.
- Dijkstra K.-D. B., Kalkman V. J., Dow R. A., Stokvis F. R., Van Tol J. Redefining the damselfly families: a comprehensive molecular phylogeny of Zygoptera (Odonata) (англ.) // Systematic Entomology. — Wiley-Blackwell, 2014. — Vol. 39, no. 1. — P. 68—96. — doi:10.1111/syen.12035. Архивировано 11 июля 2017 года.
- Жерихин В. В. Избранные труды по палеоэкологии и филоценогенетике. — М.: Litres, 2018. — С. 132—133. — 547 с.
- Bechly G. Phylogenetic Systematics of Odonata. in Schorr M. & Lindeboom M. (Eds.) // Dragonfly Research. — Zerf-Tübingen (CD-ROM), 2003. — № 1. ISSN 1438-034X
- Petrulevicius J. F., Gutiérrez P. R. New basal Odonatoptera (Insecta) from the lower Carboniferous (Serpukhovian) of Argentina // Arquivos Entomolóxicos. — 2016. — Vol. 16. — P. 341—358.
- Wilson K. D. P. Hong Kong Dragonflies. — Hong Kong: Urban Council of Hong Kong, 1995. — P. 155—157.
- Lai Y. L. An introduction to the Odonata of Hong Kong // New Asia College Academic Annual. — 1971. — Vol. 13. — P. 1—48.
- Groeneveld Linn F.; Viola Clausnitzerb and Heike Hadrysa. (2007), Convergent Evolution of Gigantism in Damselflies of Africa and South America? Evidence from Nuclear and Mitochondrial Sequence Data, Molecular Phylogenetics and Evolution Т. 42 (2): 339—346, PMID 16945555, doi:10.1016/j.ympev.2006.05.040, <http://linkinghub.elsevier.com/retrieve/pii/S1055790306002296>. Проверено 21 сентября 2013. Архивная копия от 2 августа 2017 на Wayback Machine
- Wilson K. Dragonfly Giants // Agrion. — The Worldwide Dragonfly Association, 2009. — Vol. 13, № 1. — P. 29—31. — ISSN 1476-2552. Архивировано 22 июня 2016 года.
- Tillyard R. J. The Biology of Dragonflies (Odonata or Paraneuroptera). — Cambridge: University Press, 1917.
- Silsby J. Dragonflies of the World. — Csiro Publishing, 2001. — P. 37. — 216 p. — ISBN 0643065121. Архивированная копия (недоступная ссылка). Дата обращения: 10 июля 2017. Архивировано 13 мая 2018 года.
- Ziegler A. Hawaiian Natural History, Ecology and Evolution. — University of Hawaii Press, 2002. — 477 p.
- Cooper A. Dragonflies: Q&A Guide. — Stackpole Books, 2014. — 112 p.
- Albert G. Orr, Robin W.J. Ngiam, Tzi Ming Leong. The larva of Tetracanthagyna plagiata, with notes on its biology and comparisons with congeneric species (Odonata: Aeshnidae) (англ.) // International Journal of Odonatology. — The Worldwide Dragonfly Association, 2010. — Vol. 13, no. 2. — P. 153—166. — doi:10.1080/13887890.2010.9748371.
- Garrison R.W., N. von Ellenrieder, Louton J. A. Damselfly Genera of the New World. An Illustrated and Annotated Key to the Zygoptera. — Baltimore: The Johns Hopkins University Press, 2010. — 490 p.
- Татаринов А.Г., Кулакова О.И. Фауна европейского северо-востока России. — СПб.: Наука, 2009. — Т. X Стрекозы. — С. 19—33. — 213 с. — ISBN 978-5-02-026252-9.
- Скворцов В. Э. Стрекозы Восточной Европы и Кавказа: Атлас-определитель. — М.: Товарищество научных изданий КМК, 2010. — С. 27—70. — 624 с. — 1000 экз. — ISBN 978-5-87317-657-1.
- Захваткин Ю. А. Курс общей энтомологии. — М.: Колос, 2001. — С. 37, 311—312. — 376 с. — ISBN 5-10-003598-6.
- Белышев Б. Ф. . Определитель стрекоз Сибири по имагинальным и личиночным фазам. — М.—Л.: Издательство Академии наук СССР, 1963. — С. 5—23. — 336 с. — 1100 экз.
- Акимушкин И. И. Насекомые. Пауки. Домашние животные. — 4-е изд., испр. и доп. — М.: Мысль, 1998. — С. 30. — 445 с. — (Мир животных). — ISBN 5-244-00899-4.
- Шванвич Б. Н. Курс общей энтомологии: Введение в изучение строения и функций тела насекомых. — М.—Л.: Советская наука, 1949. — С. 803. — 900 с.
- Serrano-Meneses M. A., Cordoba-Aguilar A., Azpilicueta-Amorin M., Gonzalez-Soriano E., Szekely T. Sexual selection, sexual size dimorphism and Rensch’s rule in Odonata (англ.) // Journal of Evolutionary Biology. — 2008. — Vol. 21, no. 5. — P. 1259—1273. — doi:10.1111/j.1420-9101.2008.01567.x.
- Serrano-Meneses M. A., Córdoba-Aguilar A., Székely T. Sexual size dimorphism: patterns and processes // Dragonflies and Damselflies. Model organisms for ecological and evolutionary research / Alex Córdoba-Aguilar (ed.). — Oxford, UK: Oxford University Press, 2008. — P. 231—247. — 290 p. — ISBN 978-0-19956758-4.
- Samways M. J., Osborn, R. Divergence in a transoceanic circumtropical dragonfly on a remote island (англ.) // Journal of Biogeography : Журнал. — 1998. — Vol. 25, no. 5. — P. 935—946. — ISSN 1365-2699. — doi:10.1046/j.1365-2699.1998.00245.x. (недоступная ссылка)
- Cordero A., Carbone S., Utzeri C. Mating opportunities and mating costs are reduces in androchrome female damselflies Ischnura elegans (Odonata) (англ.) // Animal Behaviour. — Elsevier, 1998. — Vol. 55. — P. 185—197.
- Белышев Б. Ф. . Стрекозы Сибири, т. I, часть 1. — Новосибирск: Наука, Сибирское отделение, 1973. — С. 36—39. — 336 с. — 1100 экз.
- Кутикова Л. А., Старобогатов Я. И. (ред.). Определитель пресноводных беспозвоночных европейской части СССР. — Л.: Гидрометеоиздат, 1977. — С. 266—268. — 513 с.
- Попова А. Н. Личинки стрекоз фауны СССР (Odonata). — Определители по фауне СССР, издаваемые Зоологическим институтом Академии наук СССР. Вып.50. — М.—Л.: Издательство Академии наук СССР, 1953. — С. 9—52. — 236 с. — 1500 экз.
- Райков Б. Е., Римский-Корсаков М. Н. Зоологические экскурсии. — 6-е. — Л.: Учпедгиз, 1956. — 694 с.
- Corbet Philip S., Suhling Frank, Soendgerath Dagmar. Voltinism of Odonata: a review (англ.) // International Journal of Odonatology. — 2006. — Vol. 9, no. 1. — P. 1—44. — doi:10.1080/13887890.2006.9748261.
- Wendler A., Nuss J.-H. Libellen. — Hamburg: DJN, 1991. — P. 129.
- Lorenzo-Carballa M. O, Cordero-Rivera A. Thelytokous parthenogenesis in the damselfly Ischnura hastata (Odonata, Coenagrionidae): genetic mechanisms and lack of bacterial infection (англ.) // Heredity : Журнал. — 2009. — Vol. 103, no. 5. — P. 377—384. — doi:10.1038/hdy.2009.65. — PMID 19513091.
- Salindra H. G., Dayananda K., Kitching R. L. Ovo-viviparity in the Odonata? The case of Heliocypha perforata (Zygoptera: Chlorocyphidae) (англ.) // International Journal of Odonatology. — Worldwide Dragonfly Association, Taylor & Francis, 2014. — Vol. 17, no. 4. — P. 181—185. — ISSN 2159-6719. — doi:10.1080/13887890.2014.959076.
- Koch K. Lifetime egg production of captive libellulids (Odonata) (англ.) // International Journal of Odonatology. — Worldwide Dragonfly Association, Taylor & Francis, 2015. — Vol. 18, no. 3. — P. 193—204. — ISSN 2159-6719.
- Bennett S., Mill P. J. Lifetime egg production and egg mortality in the damselfly Pyrrhosoma nymphula (Sulzer) (Zygoptera, Coenagrionidae) (англ.) // Hydrobiologia. — 1995. — Vol. 310. — P. 71—78.
- Stoks R., Cordoba-Aguilar A. Evolutionary Ecology of Odonata: A Complex Life Cycle Perspective (англ.) // Annual Review of Entomology. — 2012. — Vol. 57. — P. 249—265. — ISSN 0066-4170. — doi:10.1146/annurev-ento-120710-100557. Архивировано 9 августа 2017 года.
- Corbet Ph. S. Biology of Odonata (англ.) // Annual Review of Entomology. — 1985. — Vol. 25. — P. 189—217. — ISSN 0066-4170. — doi:10.1146/annurev-ento-010180-001201.
- Бешовски В. Л. Фауна Болгарии. Т. 23. Insecta, Odonata / Ред. В. Найденов. — София: Издательство Болгарской Академии наук, 1994. — P. 26. — 372 p. — ISBN 954-430-228-X.
- Борисов С. Н. Высотное распространение стрекоз (Odonata) на Памиро-Алае // Сибирский экологический журнал. — 2002. — № 6. — С. 685—696.
- Бродский А. К. Механика полёта насекомых и эволюция их крылового аппарата. — ЛГУ им. А. А. Жданова, 1988. — С. 206.
- Frequently Asked Questions about Dragonflies. British Dragonfly Society. Дата обращения: 28 февраля 2015. Архивировано 11 марта 2015 года.
- Tillyard, Robert John. The Biology of Dragonflies (неопр.). — 1917. — С. 322—323. Архивированная копия (недоступная ссылка). Дата обращения: 10 июля 2017. Архивировано 3 июля 2017 года.
- Dean T. J. Chapter 1 — Fastest Flyer. Book of Insect Records. University of Florida (1 мая 2003). Дата обращения: 10 июля 2017. Архивировано 20 июля 2011 года.
- Ingley S. J., Bybee S. M., Tennessen K. J., Whiting M. F., Branham M. A. Life on the fly: phylogenetics and evolution of the helicopter damselflies (Odonata, Pseudostigmatidae) // Zoologica Scripta. — 2012. — Vol. 41. — P. 637—650.
- Казенас В. Л., Маликова Е. И., Борисов С. Н. Стрекозы. Животные Казахстана в фотографиях. — Алматы, 2014. — С. 3. — 176 с.
- Sluvko A. A., Chuikov Y. S. Diurnal activity, distribution and power spectrum of some species of dragonflies in Astrakhan region // Ecological and biological problems of the Volga region and the Northern Caspian Sea: Proceedings of the All-Russian scientific-practical conference. 2001 October 1st. — Astrakhan: Publisher of the Astrakhan State Pedagogical University, 2001. — P. 96—98.
- Cordero, Adolfo. Sexual Cannibalism in the Damselfly Species Ischnura graellsii (Odonata: Coenagrionidae) (англ.) // Entomologia Generalis : Журнал. — Vol. 17, no. 1. — P. 17—20. — doi:10.1127/entom.gen/17/1992/17. Архивировано 13 июня 2013 года.
- Кетенчиев Х. А., Тихонова А. В. Трофические связи стрекоз (Odonata) в биоценозах // Актуальные проблемы гуманитарных и естественных наук. — 2010. — № 12. — С. 29—31.
- Corbet Ph.S. Dragonflies: Behavior and ecology of Odonata. — Colchester: Harley Books, 1999. — P. 829.
- Michael L.-М. Thermoregulation and Adaptation to Temperature in Dragonflies (Odonata: Anisoptera) // Ecological Monographs. — 1976. — Vol. 46. — P. 1—32. — doi:10.2307/1942392.
- Utzeri C., Gianandrea G. Perching in the shade: a thermoregulatory behaviour in some dispersed aeshnid dragonflies? // Opuscula Zoologica Fluminensia. — 1989. — № 35. — P. 7—8.
- Белышев Б. Ф. Определитель стрекоз Сибири по имагинальным и личиночным фазам. — М.—Л.: Издательство Академии наук СССР, 1983. — 115 с.
- Fincke O. M. Insect Populations: in Theory and Practice. — Kluwer Academic Press, 1998. — P. 391—416.
- Белышев Б. Ф. Условия существования личинок реликтовой стрекозы Orthetrum albistylum Selys в горячем ключе Северо-Восточного Прибайкалья // Зоологический журнал. — 1960. — Т. 39, № 9. — С. 1432—1433.
- Srivastava Diane S., Melnychuk Michael C., Ngai Jacqueline T. Landscape variation in the larval density of a bromeliad-dwelling zygopteran, Mecistogaster modesta (Odonata: Pseudostigmatidae) (англ.) // International Journal of Odonatology : Журнал. — 2005. — Vol. 8, no. 1. — P. 67—79. — doi:10.1080/13887890.2005.9748244.
- Харитонов А. Ю., Попова О. Н. Миграции стрекоз (Odonata) на юге Западно-Сибирской равнины // Зоологический журнал. — 2010. — Т. 89, № 11. — С. 1—9.
- Anderson R. C. Do dragonflies migrate across the western Indian Ocean? (англ.) // Journal of Tropical Ecology. — 2009. — Vol. 25. — P. 347—358. Архивировано 23 ноября 2017 года.
- Feng H-Q., Wu K-M., Ni Y-X., Cheng D-F., Guo Y–Y. Nocturnal migration of dragonflies over the Bohai Sea in northern China // Ecological Entomology. — 2006. — Vol. 31. — P. 511—520. — ISSN 1365—2311.
- May M. L., Matthews J. H. Migration in Odonata: a case study of Anax junius // Dragonflies and Damselflies. Model Organisms for Ecological and Evolutionary Research / Cordoba–Aguilar A. (ed.). — Oxford: Oxford University Press, 2008. — P. 63—77.
- Dingle H. Migration: The biology of life on the move. — Oxford: Oxford University Press, 1996.
- Holland R.A., Wikelski M., Wilcove D.S. How and why do insects migrate (англ.) // Science. — 2006. — Vol. 313. — P. 794—796. — ISSN 1095-9203.
- Попова О. Н., Харитонов А. Ю. Динамика численности и миграции стрекозы Libellula quadrimaculata L., 1758 (Odonata, Libellulidae) // Евразиатский энтомологический журнал. — 2010. — Т. 9. — Vol. 313, № 2. — С. 231—238.
- Dyatlova E.S., Kalkman V.J. The Odonata of southwestern Ukraine, with emphasis on the species of the EU Habitats Directive // Libellula. — 2008. — Vol. 313 (27), № 3/4. — P. 275—290.
- Борисов С. Н. Миграции стрекоз (Odonata) в Средней Азии: обзор. Часть 1. Широтные миграции // Евразиатский энтомологический журнал. — 2015. — Т. 4, № 3. — С. 241—255.
- Прокофьева И. В. Роль стрекоз Odonata в питании птиц // Русский орнитологический журнал. — 2004. — Т. 13, вып. 257. — С. 299—303.
- Чарлз Дарвин. Насекомоядные растения // Собрание сочинений Ч. Дарвина / Н. Г. Холодный. — М.—Л.: Издательство АН СССР, 1948. — Т. 7.
- Шевцов А.А. Ветеринарная паразитология. — М.: Колос, 1979. — С. 80—81. — 400 с.
- Кетенчиев, Х. А. Стрекозы Средиземноморья: Состав, распределение и генезис фауны (дисс. на соискание … канд. биол. наук). Нальчик, 2002. С. 1—340.
- Чаплина И. А. Фауна и экология стрекоз Казахстана (дисс. на соискание … канд. биол. наук). Новосибирск, 2004. С. 1—257.
- The Japanese Society for Odonatology (англ.). odonata.ne.jp. Архивировано 16 июня 2017 года.
- Libellula (англ.) (недоступная ссылка). libellula.org. Дата обращения: 12 августа 2018. Архивировано 7 января 2008 года.
- Залиханов К. Х. Фауна и экология стрекоз Центрального Кавказа (дисс. на соискание … канд. биол. наук). Нальчик, 2005. С. 1—201.
- Первый Всесоюзный симпозиум одонатологов. Новосибирск (англ.) (недоступная ссылка). odonata.su. Архивировано 13 октября 2012 года.
- Dragonfly Society of the Americas (DSA) (англ.). odonatacentral.org. Дата обращения: 12 августа 2018. Архивировано 12 августа 2018 года.
- Other Odonatological meetings (англ.). worlddragonfly.org. Дата обращения: 12 августа 2018. Архивировано 8 сентября 2017 года.
- The Society (The Worldwide Dragonfly Association) (англ.) (недоступная ссылка). worlddragonfly.org. Дата обращения: 12 августа 2018. Архивировано 8 сентября 2017 года.
- International Journal of Odonatology (The Worldwide Dragonfly Association) (англ.) (недоступная ссылка). worlddragonfly.org. Дата обращения: 12 августа 2018. Архивировано 12 октября 2017 года.
- Past meetings (The Worldwide Dragonfly Association) (англ.). worlddragonfly.org. Дата обращения: 12 августа 2018. Архивировано 12 октября 2017 года.
- WDA conservation and research grants (The Worldwide Dragonfly Association) (англ.). worlddragonfly.org. Дата обращения: 12 августа 2018. (недоступная ссылка)
- Polhemus D. A. 2006. Megalagrion leptodemas Архивная копия от 1 августа 2018 на Wayback Machine. The IUCN Red List of Threatened Species 2006.
- Schindler M. et al. Dragonfly associations (Insecta: Odonata) in relation to habitat variables: a multivariate approach // Hydrobiologia. — 2003. — Vol. 497. — P. 169—180.
- Красная книга Российской Федерации. — М.: Астрель, 2001. — Т. 1: Животные. — ISBN 5170048785. Архивированная копия (недоступная ссылка). Дата обращения: 28 июня 2017. Архивировано 11 сентября 2017 года.
- Червона книга України. Тваринний світ / І. А. Акімов. — К.: «Глобалконсалтинг», 2009. — 624 с. — ISBN 978-966-97059-0-7.
- Красная книга Республики Беларусь: редкие и находящиеся под угрозой исчезновения виды дикорастущих растений растения / гл. редкол.: И. М. Качановский (предс.). М. Е. Никифоров, В. И. Парфенов [и др.]. — Мн.: «Беларуская Энцыклапедыя» імя Петруся Броўкі, 2015. — 448 с. — ISBN 978-985-11-0843-1.
- Lietuvos raudonoji knyga / vyr. red. V. Rašomavičius. — Kaunas: Lututė, 2007. — 800 p. — ISBN 978-9953-692-71-3
- Duca G., et al. Cartea Roșie a Republicii Moldova. — 3. — Ch.: Știința, 2015. — 492 p. — ISBN 9975-67-311-2.
- Красная книга Казахстана. — Т. I. — Животные. — Часть 2. Беспозвоночные. — 4-е. — Алматы: Онер, 2006.
- Odonata: информация на сайте Красной книги МСОП (англ.) (Дата обращения: 14 августа 2018)
- Ulf Gärdenfors, A. J. Stattersfield. 1996 IUCN Red List of Threatened Animals. — 1996. — P. 47—48. — 455 p. — ISBN 2-8317-0335-2. Архивированная копия (недоступная ссылка). Дата обращения: 24 июля 2017. Архивировано 1 августа 2017 года.
- Odonata Specialist Group. 1996. Megalagrion jugorum Архивная копия от 3 октября 2017 на Wayback Machine. The IUCN Red List of Threatened Species 1996.
- Francisco Sánchez-Bayo, Kris A.G. Wyckhuys. Worldwide decline of the entomofauna: A review of its drivers (англ.) // Biological Conservation : Журнал. — Elsevier & Society for Conservation Biology, 2019. — Vol. 232. — P. 8—27. — ISSN 0006-3207. — doi:10.1016/j.biocon.2019.01.020. Архивировано 24 марта 2019 года.
- Gullan P. J., Cranston P. S. The Insects: An Outline of Entomology. — John Wiley & Sons. — P. 9—13. — ISBN 978-1-4051-4457-5.
- Mitchell F. L., Lasswell J. A Dazzle Of Dragonflies. — Texas A&M University Press, 2005. — P. 31—39. — 224 p.
- Moonan Wendy. Dragonflies Shimmering as Jewelry (англ.) // New York Times : Газета. — New York, 13 августа 1999. — P. E2:38. Архивировано 12 августа 2018 года.
- 88 places of Shikoku pilgrim flower. — Shinchosha, 1997. — ISBN 4106020572.
- Ли Бо, Ду Фу. Избранная лирика / Переводы Анны Ахматовой, Александра Гитовича, Леонида Бежина, Эдуарда Балашова. Составление, предисловие и примечания Леонида Бежина. — М.: «Детская литература», 1987. — 224 с.
- Славянское языкознание: X Международный съезд славистов. София, 1988 год. — М.: Наука, 1988. — С. 54.
- Mitchell Forrest L., Lasswell, James L. A Dazzle of Dragonflies. — College Station, TX: Texas A&M University Press. — P. 20—27. — ISBN 1-58544-459-6.
- Hand Wayland D. From Idea to Word: Folk Beliefs and Customs Underlying Folk Speech (англ.) // American Speech : Журнал. — Duke University Press, 1973. — Vol. 48, no. 1/2. — P. 67—76. — doi:10.2307/3087894. — .
- Тимонова Е. Стрекозы: самые древние летающие насекомые // GEO. — Москва: Axel Springer Russia, 2017. — № 8. Архивировано 14 августа 2018 года.
- Сабуро И. История японской культуры. — М.: Прогресс, 1972. — 260 с.
- Красовская В. М. Русский балетный театр начала XX века. 1. Хореографы. — Л.: Искусство, 1971. — 526 с.
- Осип Мандельштам. Сочинения в 2-х тт / Составители С. Аверинцев и П. Нерлер. — Москва: Художественная литература, 1990. — С. 285. — 640 с. — ISBN 5-280-00559-2.
- Генрих Гейне. Полное собрание сочинений Генриха Гейне / Под редакцией и с биографическим очерком Петра Вейнберга. — 2-е изд. — СПб.: Издание А. Ф. Маркса, 1904. — Т. 6. — С. 141—143.
- Басё (松尾芭蕉). Японская поэзия. japanpoetry.ru. Дата обращения: 12 августа 2018. Архивировано 12 августа 2018 года.
- Успенский Ф. Б. Habent sua fata libellulae («Дайте Тютчеву стрекозу…») // Три догадки о стихах Осипа Мандельштама. — Языки русской культуры, 2008. — С. 9—73. — 112 с. — (Studia philologica. Series minor). — 1000 экз. — ISBN 978-5-9551-0247-4.
- Strauss. La Libélula (англ.). Youtube. Дата обращения: 13 августа 2018. Архивировано 25 июля 2021 года.
- Josef Strauss Die Libelle / Polka mazurka op. 204 (1866). Commentary by Wiener Johann Strauss Orchester(WJSO) (англ.). Youtube. Дата обращения: 13 августа 2018. Архивировано 12 августа 2018 года.
- Quest Pistols Show — Белая стрекоза любви (видеоклип) (англ.). Youtube. Дата обращения: 13 августа 2018. Архивировано 10 сентября 2018 года.
- NENSI — Стрекоза (англ.). Youtube. Дата обращения: 13 августа 2018. Архивировано 25 июля 2021 года.
- Еремеев В.Е. Инженерная мысль [Китая] // Духовная культура Китая: энциклопедия: в 5 томах / гл. ред. М.Л. Титаренко; Институт Дальнего Востока. — М.: Восточная литература, 2009. — Т. 5. Наука, техническая и военная мысль, здравоохранение и образование. — С. 276—304. — 1055 с. — ISBN 978-5-036381-6.
- Brown, Roland 1955 Douglas Dragonfly. Motorcycle Classics (November–December 2007). Дата обращения: 10 августа 2009. Архивировано 14 мая 2010 года.
- Large Elizabeth. The latest buzz; In the world of design, dragonflies are flying high (англ.) // The Sun : Газета. — Baltimore, MD, 27 июня 1999. Архивировано 23 февраля 2015 года.
Литература
- Белышев Б. Ф. . Стрекозы Сибири, т. I, часть 1. — Новосибирск, 1973. — 336 с. — 1100 экз.
- Белышев Б. Ф. . Стрекозы Сибири, т. I, часть 2. — Новосибирск, 1973. — С. 337—620.
- Белышев Б. Ф. . Стрекозы Сибири, т. 2, часть 3. — Новосибирск, 1974. — 351 с.
- Белышев Б. Ф., Харитонов А. Ю. . Определитель стрекоз по крыльям. — Новосибирск: Наука, 1977. — 397 с.
- Белышев Б. Ф. Харитонов А. Ю. . География стрекоз (Odonata) Бореального фаунистического царства. — Новосибирск: Наука, 1981. — 388 с.
- Белышев Б. Ф., Харитонов, А. Ю. Харитонова И. Н., Борисов С. Н. . Фауна и экология стрекоз. — Новосибирск: Наука, 1989. — 207 с.
- Кетенчиев Х. А., Харитонов А. Ю. . Определитель стрекоз Кавказа. — Нальчик, 1998. — 118 с.
- Кетенчиев Х. А., Харитонов А. Ю. . Стрекозы Средиземноморья. — Нальчик, 1999. — 115 с.
- Мордкович В. М. Фауна и экология стрекоз. — Новосибирск: Наука, 1989. — 207 с. — ISBN 5-02-028931-0.
- Скворцов В. Э. . Стрекозы Восточной Европы и Кавказа: Атлас-определитель. — М.: Товарищество научных изданий КМК, 2010. — 624 с. — 1000 экз. — ISBN 978-5-87317-657-1.
- Татаринов А. Г., Кулакова О. И. Стрекозы (Фауна европейского Северо-Востока России. т. Х). — СПб.: Наука, 2009. — 213 с. — ISBN 978-5-02-026352-9.
- Corbet Ph. S. Dragonflies: Behavior and ecology of Odonata. — Colchester, Ithaca, New York: Harley Books, 1999. — 829 p. (англ.)
- Heckman Charles W. Encyclopedia of South American Aquatic Insects: Odonata - Anisoptera. Illustrated Keys to Known Families, Genera, and Species in South America. — Dordrecht, The Netherlands: Springer, 2006. — 725 p. — ISBN 978-1-4020-4801-2.
- Heckman Charles W. Encyclopedia of South American Aquatic Insects: Odonata - Zygoptera. Illustrated Keys to Known Families, Genera, and Species in South America. — Olympia, Washington, USA: Springer Science + Business Media B.V., 2008. — 688 p. — ISBN 978-1-4020-8175-0.
- Needham J. G., M. J. Westfall, Jr. and M. L. May. Dragonflies of North America. — Gainesville, Florida: Scientific Publishers, 2000. — 939 p. — ISBN 978-0945417941.
Ссылки
- Определители стрекоз по наядам и имаго. Стрекозы Южного Урала. (недоступная ссылка)
- Dragonflies and damselflies (Odonata) (англ.). Featured Creatures. University of Florida. Дата обращения: 29 июня 2017. Архивировано 8 июля 2017 года.
- Dragonflies and Damselflies (англ.). Odonata Information Network. Дата обращения: 29 июня 2017. Архивировано 14 октября 2017 года.
- OdonataCentral (англ.). Dragonfly Soceity of the America. University of Texas at Austin. Дата обращения: 29 июня 2017. Архивировано 29 июня 2017 года.
- Dragonflies (англ.). Slater Museum of Natural History. University of Puget Sound, Tacoma, WA. pugetsound.edu. Дата обращения: 29 июня 2017. Архивировано 6 февраля 2017 года.
- Resources for the study of Odonata in Canada (англ.). Newsletter of the Biological Survey of Canada. Biological Survey of Canada. Дата обращения: 7 июля 2017.
| Таксономия | |
|---|---|
| В библиографических каталогах |

Систематика современных стрекоз (Odonata) | ||
|---|---|---|
| ||
| Равнокрылые стрекозы |
|  |
| Разнокрылые стрекозы |
| |
| Anisozygoptera | ||