Разнокрылые стрекозы
Разнокрылые стрекозы[1][2] (лат. Anisoptera) — инфраотряд крылатых насекомых из отряда стрекоз (Odonata). Обладают крупными фасеточными глазами, двумя парами сильных прозрачных крыльев и удлинённым телом. Задние крылья у разнокрылых стрекоз отличаются по форме от передних — они шире у основания. От представителей другой крупной группы отряда — равнокрылых стрекоз — отличаются перпендикулярным к продольной оси тела положением крыльев в спокойном состоянии. Разнокрылые стрекозы — самые быстрые летающие насекомые, охотящиеся на москитов, мух, пчёл, муравьёв и бабочек. Например, стрекоза Austrophlebia costalis может летать со скоростью до 97 км в час[3][4][5]. Максимальная скорость крупных стрекоз достигает 10–15 м/сек (36–55 км/час)[6].
| Разнокрылые стрекозы | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Somatochlora flavomaculata | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Отряд: Подотряд: Разнокрылые стрекозы |
||||||||||
| Международное научное название | ||||||||||
| Anisoptera Selys, 1854 | ||||||||||
| Надсемейства | ||||||||||
|
||||||||||
| ||||||||||
Обычно встречаются вблизи водоёмов и болот, так как их личинки («нимфы») живут в воде.
Описание имаго
К подотряду относятся крупные и очень крупные, хорошо летающие, обычно ярко окрашенные насекомые[7].


Голова крупная, шире всех остальных отделов тела. Она обособлена от переднегруди и сочленена с ней чрезвычайно подвижно, благодаря чему способна поворачиваться вокруг продольной оси тела на 180°. Бо́льшую часть головы занимают очень крупные полушаровидные сложные фасеточные глаза. У разнокрылых стрекоз глаза более или менее смещены ко лбу. У Gomphidae они разделены промежутком, меньшим, чем диаметр глаз. У булавобрюхов глаза соприкасаются в одной точке, а у настоящих стрекоз и коромысел — на протяжении некоторого отрезка[8]. Ротовой аппарат мощный, хорошо развитый, грызущего типа. Он образован непарными верхней и нижней губами, парными острозубчатыми верхними (мандибулы) и нижними (максиллы) челюстями. Верхняя губа прикрывает сверху челюсти, она в виде короткой пластинки, вытянутой в поперечном направлении, с полукруглым внешним краем. Нижняя губа из трёх лопастей (средней и двух боковых) покрывает собой челюсти снизу. Она крупнее верхней, наиболее развита у разнокрылых стрекоз, которые жуют добычу на лету. Парные челюсти размельчают пищу, двигаясь при этом в горизонтальной плоскости. Челюсти стрекоз чрезвычайно прочные и характеризуются острыми твёрдыми зубцами. На максиллах также имеются щупики, выполняющие осязательную роль[8].
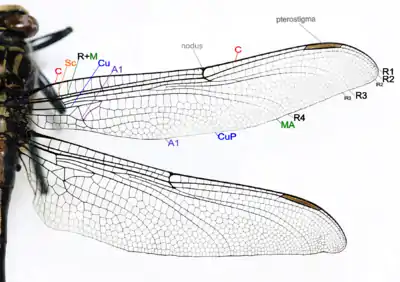
Грудь хорошо развитая; как и у всех насекомых, состоит из переднегруди, среднегруди и заднегруди. Каждая из частей несёт на себе одну пару конечностей, а среднегрудь и заднегрудь ещё и по паре крыльев каждая. Целый ряд особенностей строения груди стрекоз является своеобразным. Переднегрудь отделена от среднегруди, сильно редуцирована (иногда практически целиком помещается в углублении задней стороны головы). Задне- и среднегрудь хорошо развитые и плотно сросшиеся между собой, образуют синторакальный комплекс, или синторакс, который воспринимается как собственно грудь и часто так и называется. Склериты каждого сегмента отделены друг от друга швами. Боковой шов между средне- и заднегрудью развит только у представителей семейства Calopterygidae[8]. На каждом крыле имеется шесть основных продольных жилок[9]:
- Костальная жилка (costa, С) — образует собой весь передний (костальный) край крыла.
- Субкостальная жилка (subcosta, Sc) — располагается параллельно костальной жилке на расстоянии одного ряда ячеек, но короче неё.
- Радиальная жилка (radius, R) — представляет собой целую систему ветвей, называемых секторами. От основания крыла радиальная жилка слита со следующей, медиальной жилкой (medius, M); этот отрезок обозначается как R+M.
- Кубитальная жилка (cubitus, Си) — не ветвясь, идёт от основания крыла до средней части его заднего края, параллельно медиальной жилке, лишь в конце загибаясь назад.
- Анальная жилка (А) — проходит более или менее параллельно кубитальной жилке и заканчивается на заднем крае крыла.
У представителей отряда крылья заметно различны, задние крылья отличаются по форме от передних — они шире у основания[9]. К телу передние крылья прикрепляются всем основанием, а задние крылья в своей базальной части несколько шире передних и прикрепляются только верхней, стебельчато суженной частью расширенного основания[8]. В основании крыла, между медиальной и кубитальной жилками, у равнокрылых стрекоз находится поле, которое называется треугольником. Непрозрачное поле в основании крыла, свойственное большинству разнокрылых стрекоз, получило название перепоночки[9]. Важной особенностью строения крыла стрекоз является наличие птеростигмы и узелка. Птеростигма — утолщение у переднего края крыла, расположенное ближе к его вершине. Она состоит из утолщённых (иногда дополнительно пигментированных) жилок и ячеек и внешне выглядит как плотный тёмноокрашенный участок края крыла. Птеростигма увеличивает амплитуду размаха крыла за счёт утяжеления его вершины и, вероятно, способна гасить вредные колебания (флаттер) крыла[10].
Конечности состоят из 5 основных частей — тазика, вертлуга, бедра, голени и лапки, которые соединены подвижными суставами. Неподвижно соединены лишь дистальный конец вертлуга и проксимальный конец бедра, сросшиеся друг с другом. Форма ног практически постоянна между представителями; их длина увеличивается от передней пары к задней. Расстояния между парами ног несколько неравные — средняя пара ближе к задней, чем к передней[8].
_(4).JPG.webp)
Брюшко очень длинное, палочкообразное. Чаще оно в сечении округлое, тонкое, реже — широкое и плоское. На его долю приходится до 90 % от всей длины тела. Брюшко состоит из десяти полных сегментов и одиннадцатого рудиментарного сегмента (тельсон). Каждый сегмент образован двумя хитиновыми щитками. Верхний щиток (тергит) дуговидный в поперечном сечении, заходя на брюшную сторону, частично перекрывает нижний (стернит). Последний, в отличие от тергита, имеет слабо выгнутую форму и покрывает брюшко только снизу. По бокам между тергитами и стернитами располагаются растяжимые плевральные перепонки, позволяющие стрекозе изгибать брюшко, а также сжимать и разжимать его при дыхании и откладывании яиц. С каждой стороны на каждом сегменте (кроме IX и X) на плевральной перепонке находится по одному дыхальцу (стигме). Характерная черта строения брюшка самцов большинства видов коромысел и булавобрюхов — наличие продольных выростов на каждой стороне нижней поверхности второго тергита — так называемых ушек (auriculae). Рудиментарные ушки имеются и у самок многих представителей семейства Gomphidae[8]. На конце брюшка располагаются нечленистые анальные придатки, число которых у самок постоянно и равно 2, а у самцов стрекоз разных таксономических групп — либо 4 (по паре верхних и нижних), либо 3 (пара верхних и непарный нижний). Генитальные органы у самок располагаются на конце брюшка. У самцов копулятивный орган — так называемый вторичный копулятивный аппарат[7] — располагается на брюшной стороне второго сегмента брюшка, при этом первичное спермовыводящее отверстие находится на стерните десятого сегмента[8].
Окраска тела и крыльев представителей подотряда весьма разнообразна. У стрекоз, как и других групп насекомых, различают структурную и пигментную окраску[11]. У некоторых видов отмечается сочетание этих двух типов окраски. Структурная окраска связана с некоторыми специфическими особенностями поверхностной структуры кутикулы (микроскопическими рёбрышками, пластинками и т. п.), которые создают эффекты интерференции, дифракции и дисперсии света, результатом чего являются металлически блестящие оттенки в окраске[12].
Морфология преимагиальных стадий
Личинок стрекоз часто называют нимфами либо наядами. Тело у личинок имеет различную форму. У представителей группы оно очень короткое и толстое, совершенно не похоже на тело взрослой стрекозы. Голова у личинок стрекоз крупная и широкая и, за исключением ротового аппарата, устроена так же, как у имаго. Характерной чертой строения головы личинок стрекоз является ротовой аппарат, состоящий из непарной верхней губы, парных верхних челюстей, парных нижних челюстей и непарной нижней губы. Нижняя губа имеет очень своеобразное строение и преобразована в специальное хватательное приспособление для ловли добычи — маску, являющуюся уникальным органом личинок стрекоз[8][13]. При захвате добычи она стремительно раскрывается и выбрасывается вперёд, при этом зубцы на её переднем конце глубоко вонзаются в жертву. Когда маска складывается, добыча подтягивается ко рту и спокойно пережёвывается. В состоянии покоя маска прикрывает голову снизу и (или) с боков[14]. Маска состоит из подподбородка (submentum), подбородка (mentum), дистальной лопасти, слитой с подбородком, и губных щупиков, преобразованных в боковые лопасти. Подбородок своей вершиной подвижно сочленяется с пластинкой подподбородка, который в свою очередь подвижно соединяется с головой личинки, вследствие чего маска может выдвигаться вперёд и затем складываться[14].
.jpg.webp)
Существует два типа масок: плоская и выпуклая (шлемовидная)[13]. Плоская маска в состоянии покоя прикрывает рот личинки только снизу (её средняя пластинка более или менее плоская, боковые лопасти лежат вместе с ней в одной плоскости), а шлемовидная маска прикрывает одновременно сверху, спереди и снизу, как забрало или черпак. Этот тип характерен, например, для представителей родов коромысла и дозорщики, которые хватают добычу большими подвижными зубцами на её боковых лопастях. Второй тип масок характерен для личинок, обитающих в иле, например, у таких родов, как плоскобрюхи и булавобрюхи. Такие личинки захватывают добычу вместе с илом, а затем мелкие частицы просеиваются через своеобразное «сито», образованное длинными средними и боковыми щетинками на маске[13][15].
Бо́льшую часть жизни личинки стрекоз не могут дышать атмосферным воздухом и для дыхания используют жабры, расположенные на конце тела, которые могут быть внутренними (ректальными) или наружными (каудальными). На брюшке личинок разнокрылых стрекоз трахейные жабры расположены в специальной камере в прямой кишке (ректальные жабры), которая периодически заполняется водой[8].
Развитие и размножение
.jpg.webp)
Стрекозы развиваются с неполным превращением: они проходят стадии яйца, личинки (наяды) и имаго. Многие стрекозы способны производить более одной генерации в год (вольтинизм)[16]. Спаривание стрекоз происходит в воздухе. Самцы разнокрылых стрекоз захватывают самку своими анальными придатками за голову. Совокупительные органы самцов располагаются на II сегменте брюшка (исключительный случай среди насекомых), а семявыводящие протоки на IX сегменте. Предварительно перекачав сперму из девятого во второй абдоминальный сегмент, самцы начинают гоняться за самками. Продолжительность спаривания различается у разных видов и может длиться от нескольких секунд до трёх часов.
После спаривания оплодотворённые самки откладывают яйца прямо на воду, на водоросли, прибрежный ил, мох, гниющие растительные остатки (разнокрылые семейства Aeshnidae) либо в специальные углубления в тканях растений[9][17].
Личинки стрекоз ведут водный образ жизни. Обычно они живут в медленно текущих или стоячих пресных водоёмах (заросшие пруды, озёра, старицы рек, канавы, болота). Булавобрюх увенчанный — единственный вид стрекоз, обитающий в горах Средней Азии, преимагинальные фазы которого могут развиваться в горных ручьях с преимущественно ледово-снеговым питанием[18]. Большинство личинок стрекоз живёт на дне или на подводных растениях. Некоторые закапываются в грунт (Gomphinae, Cordulegastridae).
Личинки являются малоподвижными и обычно сидят среди подводной растительности или зарываются в грунт, терпеливо поджидая добычу. Плавают не очень хорошо и редко, обычно будучи потревоженными или спасаясь от врагов. Личинки стрекоз, как и взрослые особи, являются хищниками, питаясь главным образом личинками водных насекомых. Разнокрылые стрекозы питаются активно двигающимися личинками водных животных (веснянок, жуков, комаров, подёнок), мальками рыб, а также пиявками, головастиками, водными клопами, рачками-осликами[15].
По особенностям строения, обитания и поведения выделяют несколько морфоэкологических типов личинок
- Длиннобрюхие и активно плавающие личинки, обитающие в стоячих водах. Это виды семейства коромысла, передвигающиеся реактивным способом, выталкивая воду из прямой кишки.
- Волосатые личинки, зарывающиеся в грунт. Характерен для семейства дедки. Активные хищники, поедающие крупную добычу, включая мальков рыб и головастиков.
- Короткобрюхие ползающие личинки, обитающие в стоячих водоёмах. Отмечен в семействах бабки и настоящие стрекозы.
Особенности биологии
.JPG.webp)
Основной формой передвижения стрекоз является полёт, помогающий им искать пищу и полового партнёра для размножения, расселяться и мигрировать, спасаться от хищников. Разнокрылые стрекозы характеризуются быстрым резким полётом, способностью отлетать на далёкие расстояния от водоёмов и спариваться вдали от водоёма[8]. Скорость полёта некоторых крупных разнокрылых стрекоз достигает 15 м/с[19] (36—55 км/ч)[6]. Стрекоза Austrophlebia costalis способна летать со скоростью до 97 км/ч[20][21].
Взрослые стрекозы — активные хищники воздушной среды. Наличие индивидуальных охотничьих участков отмечено у представителей семейств коромысла (Aeshnidae) и бабки (Corduliidae), видов рода настоящие стрекозы (Libellula).
У стрекоз выделяются несколько групп по способам охоты:
- Свободные охотники верхнего яруса — виды с развитыми крыльями, обитающие на открытых пространствах, часто вдали от водоёмов. Охотятся на высоте 2—10 м. Каждая стрекоза контролирует индивидуальный охотничий участок, иногда даже несколько. При наличии большого количества добычи у них может наблюдаться «роевая охота», когда стрекозы собираются в подобие охотничьих стай численностью до 20 особей. К этой группе относятся представители семейств булавобрюхи, коромысла и других.
- Свободные хищники среднего яруса — виды, которые охотятся главным образом над поверхностью воды водоёмов на высоте 0,5—2 м. Такие стрекозы практически постоянно находятся в полёте, изредка присаживаясь на прибрежные или торчащие из воды растения. К этой группе относится большинство представителей семейств дедки и бабки.
- Подстерегающие хищники среднего яруса — стрекозы, большую часть времени охоты проводящие на «наблюдательных пунктах» (верхушках трав и кустарников, торчащих из воды ветках и стеблях растений) и время от времени срывающиеся в погоню за пролетающими насекомыми. К этой группе относятся представители семейства настоящие стрекозы.
Весьма широко у стрекоз развит каннибализм — поедание более мелких стрекоз крупными представителями других видов, особенно широко это явление распространено у крупных стрекоз[22][23].
Характерной особенностью крупных стрекоз (Anax, Aeshna, Orthetrum, Crocothemis, Sympetrum, Libellula и др.) является совершение «патрулирующих» полётов. У них отмечается хорошо выраженное территориальное поведение, заключающееся в наличии индивидуальных кормовых участков[24].
Разнокрылые стрекозы активны в дневное время в тёплую солнечную погоду. В утренние часы и при низких температурах стрекозы часто сидят, греясь под лучами солнца, на камнях, коре деревьев и прочих субстратах, которые способны к быстрому теплообмену. При высоких температурах в дневное время, чтобы избежать перегрева, настоящие стрекозы и дедки, сидя, принимают характерное положение «обелиска» — ориентируют кончик брюшка вверх к солнцу, снижая воздействие тепла на тело[25][26].
Разнокрылые стрекозы в целом проявляют исключительную пластичность в выборе водоёмов для развития своих личинок: для них подходящими являются все водоёмы, начиная от горных потоков и ручьёв, прудов, озёр, стариц рек, ручьёв и заканчивая солоноватыми и торфяными водоёмами, вплоть до созданных человеком прудов, оросительных каналов и канав. Примечательно, что в таком большом озере, как Байкал, стрекозы отсутствуют[27].
Как и многие другие насекомые, разнокрылые стрекозы могут совершать миграции, во время которых могут преодолевать океаны и высокие горные хребты, летая ночью и на больших высотах[28][29][30]. Миграции с севера на юг характерны для Anax junius в Северной Америке; некоторые мигранты перелетали на расстояние до 2800 км с северной на южную части континента[31]. Стрекозы всемирно распространённого рода бродяжки используют пассивное распространение, чтобы пересечь Индийский океан[29], при этом поддерживая местные островные популяции, например, в Тихом океане на острове Пасхи[32]. Адаптивная функция миграции заключается в перемещении отдельных особей и популяций из ухудшающейся со временем среды обитания в более благоприятную на текущий момент[33].
Классификация

В подотряде выделяют следующие современные таксоны уровня семейства[34]:
- Надсемейство Aeshnoidea
- Семейство Aeshnidae — Коромысла, 456 видов
- Семейство Austropetaliidae — 11 видов
- Надсемейство Petaluroidea
- Семейство Petaluridae — 10 видов
- Надсемейство Gomphoidea
- Надсемейство Cordulegastroidea
- Семейство Chlorogomphidae — 47 видов
- Семейство Cordulegastridae — Булавобрюхи, 46 видов
- Семейство Neopetaliidae — 1 вид
- Надсемейство Libelluloidea
- Семейство Synthemistidae — 46 видов
- Семейство Macromiidae — 125 видов
- Семейство Corduliidae — Бабки, 154 вида
- Семейство Libellulidae — Настоящие стрекозы, 1037 видов
- Роды incertae sedis — 19 родов с 98 видами
Разнообразие и распространение по семействам
Около 3000 видов (на 2010 год) и 348 родов встречаются во всех зоогеографических областях[35].
| Семейство | Индо-малайская зона | Неотропика | Австралазия | Афротропика | Палеарктика | Неарктика | Океания | В мире |
|---|---|---|---|---|---|---|---|---|
| Aeshnidae | 149 | 129 | 78 | 44 | 58 | 40 | 13 | 456 |
| Austropetaliidae | 7 | 4 | 11 | |||||
| Petaluridae | 1 | 6 | 1 | 2 | 10 | |||
| Gomphidae | 364 | 277 | 42 | 152 | 127 | 101 | 980 | |
| Chlorogomphidae | 46 | 5 | 47 | |||||
| Cordulegastridae | 23 | 1 | 18 | 46 | ||||
| Neopetaliidae | 1 | 1 | ||||||
| Corduliidae | 23 | 20 | 33 | 6 | 18 | 51 | 12 | 154 |
| Libellulidae | 192 | 354 | 184 | 251 | 120 | 105 | 31 | 1037 |
| Macromiidae | 50 | 2 | 17 | 37 | 7 | 10 | 125 | |
| Synthemistidae | 37 | 9 | 46 | |||||
| Incertae sedis | 37 | 24 | 21 | 15 | 2 | 99 | ||
Примечания
- Определитель насекомых Дальнего Востока СССР. Т. I. Первичнобескрылые, древнекрылые, с неполным превращением / под общ. ред. П. А. Лера. — Л.: Наука, 1988. — С. 147. — 452 с.
- Стриганова Б. Р., Захаров А. А. Пятиязычный словарь названий животных: Насекомые (латинский-русский-английский-немецкий-французский) / Под ред. д-ра биол. наук, проф. Б. Р. Стригановой. — М.: РУССО, 2000. — С. 6. — 1060 экз. — ISBN 5-88721-162-8.
- Tillyard, Robert John. The Biology of Dragonflies (англ.). — 1917. — P. 322—323.
- Dean, T. J. Chapter 1 — Fastest Flyer. Book of Insect Records. University of Florida (1 мая 2003). Архивировано 20 июля 2011 года.
- Hocking, B. 1953. The intrinsic range and speed of flight of insects: Trans. R. ent. Soc. lond., 104, 225-345.
- Frequently Asked Questions about Dragonflies. British Dragonfly Society. Дата обращения: 28 февраля 2015.
- Татаринов А.Г., Кулакова О.И. Фауна европейского северо-востока России. — СПб.: Наука, 2009. — Т. X Стрекозы. — С. 19—33. — 213 с. — ISBN 978-5-02-026252-9.
- Скворцов В. Э. Стрекозы Восточной Европы и Кавказа: Атлас-определитель. — М.: Товарищество научных изданий КМК, 2010. — С. 27—70. — 624 с. — 1000 экз. — ISBN 978-5-87317-657-1.
- Белышев Б. Ф. . Определитель стрекоз Сибири по имагинальным и личиночным фазам. — М.—Л.: Издательство Академии наук СССР, 1963. — С. 5—23. — 336 с. — 1100 экз.
- Акимушкин И. И. Насекомые. Пауки. Домашние животные. — 4-е изд., испр. и доп. — М.: Мысль, 1998. — С. 30. — 445 с. — (Мир животных). — ISBN 5-244-00899-4.
- Захваткин Ю. А. Курс общей энтомологии. — М.: Колос, 2001. — С. 37, 311—312. — 376 с. — ISBN 5-10-003598-6.
- Шванвич Б. Н. Курс общей энтомологии: Введение в изучение строения и функций тела насекомых. — М.—Л.: Советская наука, 1949. — С. 803. — 900 с.
- Кутикова Л. А., Старобогатов Я. И. (ред.). Определитель пресноводных беспозвоночных европейской части СССР. — Л.: Гидрометеоиздат, 1977. — С. 266—268. — 513 с.
- Райков Б. Е., Римский-Корсаков М. Н. Зоологические экскурсии. — 6-е. — Л.: Учпедгиз, 1956. — 694 с.
- Попова А. Н. Личинки стрекоз фауны СССР (Odonata). — Определители по фауне СССР, издаваемые Зоологическим институтом Академии наук СССР. Вып.50. — М.—Л.: Издательство Академии наук СССР, 1953. — С. 9—52. — 236 с. — 1500 экз.
- Corbet Philip S., Suhling Frank, Soendgerath Dagmar. Voltinism of Odonata: a review (англ.) // International Journal of Odonatology. — 2006. — Vol. 9, no. 1. — P. 1—44. — doi:10.1080/13887890.2006.9748261.
- Белышев Б. Ф. . Стрекозы Сибири, т. I, часть 1. — Новосибирск: Наука, Сибирское отделение, 1973. — С. 36—39. — 336 с. — 1100 экз.
- Борисов С. Н. Высотное распространение стрекоз (Odonata) на Памиро-Алае // Сибирский экологический журнал. — 2002. — № 6. — С. 685—696.
- Бродский А. К. Механика полёта насекомых и эволюция их крылового аппарата. — ЛГУ им. А. А. Жданова, 1988. — С. 206.
- Tillyard, Robert John. The Biology of Dragonflies (неопр.). — 1917. — С. 322—323. Архивированная копия (недоступная ссылка). Дата обращения: 25 марта 2017. Архивировано 3 июля 2017 года.
- Dean T. J. Chapter 1 — Fastest Flyer. Book of Insect Records. University of Florida (1 мая 2003). Дата обращения: 10 июля 2017. Архивировано 20 июля 2011 года.
- Казенас В. Л., Маликова Е. И., Борисов С. Н. Стрекозы. Животные Казахстана в фотографиях. — Алматы, 2014. — С. 3. — 176 с.
- Sluvko A. A., Chuikov Y. S. Diurnal activity, distribution and power spectrum of some species of dragonflies in Astrakhan region // Ecological and biological problems of the Volga region and the Northern Caspian Sea: Proceedings of the All-Russian scientific-practical conference. 2001 October 1st. — Astrakhan: Publisher of the Astrakhan State Pedagogical University, 2001. — P. 96—98.
- Кетенчиев Х. А., Тихонова А. В. Трофические связи стрекоз (Odonata) в биоценозах // Актуальные проблемы гуманитарных и естественных наук. — 2010. — № 12. — С. 29—31.
- Corbet Ph.S. Dragonflies: Behavior and ecology of Odonata. — Colchester: Harley Books, 1999. — P. 829.
- Michael L.-М. Thermoregulation and Adaptation to Temperature in Dragonflies (Odonata: Anisoptera) // Ecological Monographs. — 1976. — Vol. 46. — P. 1—32. — doi:10.2307/1942392.
- Белышев Б. Ф. Определитель стрекоз Сибири по имагинальным и личиночным фазам. — М.—Л.: Издательство Академии наук СССР, 1983. — 115 с.
- Харитонов А. Ю., Попова О. Н. Миграции стрекоз (Odonata) на юге Западно-Сибирской равнины // Зоологический журнал. — 2010. — Т. 89, № 11. — С. 1—9.
- Anderson R. C. Do dragonflies migrate across the western Indian Ocean? (англ.) // Journal of Tropical Ecology. — 2009. — Vol. 25. — P. 347—358. Архивировано 23 ноября 2017 года.
- Feng H-Q., Wu K-M., Ni Y-X., Cheng D-F., Guo Y–Y. Nocturnal migration of dragonflies over the Bohai Sea in northern China // Ecological Entomology. — 2006. — Vol. 31. — P. 511—520. — ISSN 1365—2311.
- May M. L., Matthews J. H. Migration in Odonata: a case study of Anax junius // Dragonflies and Damselflies. Model Organisms for Ecological and Evolutionary Research / Cordoba–Aguilar A. (ed.). — Oxford: Oxford University Press, 2008. — P. 63—77.
- Samways M. J., Osborn, R. Divergence in a transoceanic circumtropical dragonfly on a remote island (англ.) // Journal of Biogeography : Журнал. — 1998. — Vol. 25, no. 5. — P. 935—946. — ISSN 1365-2699. — doi:10.1046/j.1365-2699.1998.00245.x. (недоступная ссылка)
- Dingle H. Migration: The biology of life on the move. — Oxford: Oxford University Press, 1996.
- Dijkstra K. D. B., Bechly G., Bybee S. M., Dow R. A., Dumont H. J., et al. The classification and diversity of dragonflies and damselflies (Odonata) (англ.) // Zootaxa. — 2013. — Vol. 3703, no. 1. — P. 36—45. — doi:10.11646/zootaxa.3703.1.9.
- Suhling F., Sahlén G., Gorb S., Kalkman V.J., Dijkstra K-D.B., van Tol J. Order Odonata // Ecology and general biology. Thorp and Covich's Freshwater Invertebrates (англ.) / Thorp, James & D. Christopher Rogers. — 4 ed. — Academic Press, 2015. — P. 893—932. — ISBN 9780123850263.
Литература
- Alexander BLANKE, Carola GREVE, Rajmund MOKSO, Felix BECKMANN, Bernhard MISOF. 2013. An updated phylogeny of Anisoptera including formal convergence analysis of morphological characters. Systematic Entomology. Volume 38, Issue 3, July 2013 Pages 474–490. DOI: 10.1111/syen.12012
| Таксономия | |
|---|---|
| В библиографических каталогах |
Систематика современных стрекоз (Odonata) | ||
|---|---|---|
| ||
| Равнокрылые стрекозы |
|  |
| Разнокрылые стрекозы |
| |
| Anisozygoptera | ||