Терморегуляция насекомых
Терморегуляция насекомых — способность насекомых поддерживать температуру тела в определённых границах, даже если температура внешней среды сильно отличается.
Насекомые традиционно рассматриваются как пойкилотермные организмы (у которых температура тела варьируется и зависит от температуры окружающей среды), в отличие от теплокровных организмов (которые поддерживают стабильную внутреннюю температура тела, независимо от внешних воздействий). Тем не менее, термин «терморегуляциия», в настоящее время используется для описания способности насекомых поддерживать стабильную температуру (выше или ниже температуры окружающей среды), по меньшей мере, в части их тела, физиологическими или поведенческими средствами[1].
В то время как многие насекомые являются эктотермами (животные, у которых их источником тепла является окружающая среда), другие насекомые могут быть эндотермами (животные, которые могут самостоятельно производить тепло внутри своего тела биохимическими процессами). Эндотермических насекомых корректней описать как гетеротермов, потому что они не являются целиком эндотермическими. Когда тепло производится, различные части тела насекомых могут иметь разную температуру, например, ночные бабочки производят тепло в груди для полёта, но их брюшко при этом значительно не нагревается[2].
Терморегуляция в полёте
Полёт является очень энергозатратным способом передвижения, требующим высокого уровня обмена веществ. Чтобы летать, животному необходимо иметь мышцы, способные вырабатывать большие механические мощности, что, в свою очередь, вследствие неэффективности биохимических процессов, приводит к выработке больших количеств тепла[3]. Таким образом, насекомое в полёте нагревается, что допустимо, пока температура остается ниже определенного значения. Однако, если насекомое подвергается воздействию внешнего источника тепла (к примеру, солнечных лучей), или температура окружающей среды высока, ему необходима способность регулировать свою температуру, чтобы она оставалась в зоне комфорта.
Насекомое может терять тепло за счет простого теплообмена с окружающей средой, на более высоких скоростях отток тепла будет усиливаться. Тем не менее, увеличение скорости полёта приводит к повышению, а не понижению температуры[4]. Причина, вероятно, кроется в том, что мышцы работают усиленнее, а это вызывает повышения выделение тепла в области груди насекомого.
Первые свидетельства о терморегуляции насекомых в полёте были получены в ходе экспериментов, показавших, что у высших разноусых бабочек отвод тепла происходит за счет движения гемолимфы от груди к брюшку[5]. Сердце бабочек перекачивает гемолимфу через середину груди, что содействует теплообмену и превращает брюшко в теплоизлучатель, что позволяет насекомому поддерживать стабильную температуру груди при разных температурах окружающей среды. Таким образом, считалось, что терморегуляция достигается только за счет регулирования теплопотерь до того, как были получены свидетельства о регулировании теплопроизводства у пчёл.[6] Затем была высказана мысль, что постоянство температуры у пчёл, и, вероятно, многих других гетеротермных насекомых, в первую очередь достигается за счет изменения выработки тепла. Способны ли насекомые регулировать температуру груди путём регулирования производства тепла или только путём изменения потерь тепла, по-прежнему является предметом обсуждения.
Предполётное разогревание

Некоторые крупные насекомые научились разогреваться перед полётом, чтобы такой энергозатратный процесс как полёт был возможен.[7] Мышцы насекомого работают «вхолостую», производится большое количество тепла, благодаря чему достигается диапазон температур, в котором мышцы работают наиболее эффективно. Летательные мышцы потребляют большое количество химической энергии, но лишь очень малая её доля переходит в полезную механическую работу (движение крыльев).[8] Оставшаяся часть этой химической энергии преобразуется в тепло, вследствие чего температура тела получается значительно выше температуры окружающей среды.
То, что летательные мышцы работают лучше всего при высоких температурах, создает проблемы для низкотемпературного взлёта, поскольку мышцы в состоянии покоя имеют температуру окружающей среды, не оптимальную для их работы. Поэтому, гетеротермные насекомые приспособились использовать теплоту, вырабатываемую летательными мышцами, чтобы поднимать температуру груди перед полётом. И мышцы, тянущие крылья вниз, и мышцы, тянущие крылья вверх, участвуют в предполётном разогревании, но несколько иначе, чем в полёте. Во время полёта они работают как антагонисты, чтобы поддерживать движение крыльев, необходимое для обеспечения устойчивого полёта. Однако во время разогревания эти мышцы сокращаются одновременно (или, у некоторых насекомых, почти одновременно),[9] чтобы не приводить (или почти не приводить) крылья в движение и чтобы производить как можно больше тепла.
Интересно, что предполётное разогревание самцов бабочек вида Helicoverpa zea, как показано, зависит от получаемой обонятельной информации.[10][11][12] Как и у многих других ночных бабочек, самцы этого вида реагируют на феромоны, испускаемые самками, летя к самке и пытаясь с ней спариться. Во время разогрева летательных мышц при присутствии в воздухе феромона, самцы вырабатывают тепло усиленнее, с тем чтобы взлететь раньше и опередить других самцов, которые также могли почувствовать феромоны.
Получение повышенных температур так, как описано выше, называется «физиологической терморегуляцией», потому как тепло генерируется за счет физиологических процессов внутри насекомого. Другой описанный способ терморегуляции называется «поведенческой терморегуляцией», в этом случае температура тела регулируется за счет особого поведения насекомого, такого, как, например, грение на солнце. Бабочки — хороший пример таких насекомых, они берут тепло почти исключительно от Солнца.[13]
Другие примеры терморегуляции
У некоторых жуков, питающихся навозом, повышается скорость скатывания и скорость перекатывания навозных шариков когда температура их груди увеличивается.[14] Для этих жуков навоз — это ценный ресурс, позволяющий им найти пару и прокормить личинок. Скорейшее обнаружение навоза важно, чтобы жук смог скатывать шарик как можно быстрее и отнести его в отдаленное место для схоронения. Жуки сперва обнаруживают навоз с помощью обоняния и быстро летят на запах. Сразу после приземления температура их тела высока из-за того, что в полёте они нагреваются, что позволяет им скатывать и перемещать шарики быстрее. Чем больше получившийся шарик, тем выше их шансы найти пару. Однако по прошествии некоторого времени приземлившийся жук остывает, и ему становится сложнее увеличивать размер шарика, равно как и перемещать его. Таким образом, есть компромисс между скатыванием большого шарика, гарантирующего спаривание, но который сложнее перемещать, и шарика меньшего размера, который может не привлечь самку, но который может быть безопасно отнесен для схоронения. Кроме того, другие жуки, прибывшие позже, и, как следствие, более теплые, могут пытаться отнять шарики у уже остывших жуков, и, как показано, обычно успешно это делают.[15]

Другой интересный пример терморегуляции — это использование вырабатываемого тепла для защиты. На восковых пчёл (Apis cerana japonica) охотятся шершни, обычно ожидающие их у входа в улей. Хотя шершень значительно больше пчёл, пчёлы берут числом. Они могут выживать при температурах выше 46 °C, в то время как шершень — нет. Поэтому пчёлы могут убить шершня, облепив его собой со всех сторон и затем повысив свою температуру тела.[16]
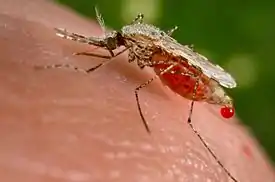
Малярийные комары вынуждены регулировать температуру тела каждый раз после того, как они забирают кровь у теплокровных животных. Во время всасывания они выделяют каплю жидкости, состоящую из мочи и крови жертвы, которую они оставляют прикрепленной к анусу. Капля испаряется, забирая излишек тепла. Такой механизм охлаждения позволяет комару избегать стресса, связанного с резким повышением температуры.[17]
Примечания
- Heinrich, Bernd (1993), The hot-blooded insects: Strategies and mechanisms of thermoregulation, Cambridge, Massachusetts: Harvard University Press, с. 601, ISBN 0-674-40838-1, <https://books.google.com/books?id=NHYuT94xvWYC&printsec=frontcover&source=gbs_ge_summary_r&cad=0#v=onepage&q&f=false>
- Heinrich, Bernd, ed. (1981), Insect thermoregulation, New York: John Wiley & Sons, Inc., с. 328, ISBN 0-471-05144-6, <https://books.google.com/books?id=ww6LAAAAIAAJ&dq=editions:VVOId1Kwp40C>
- Josephson, R.K. & Stevenson, R.D. (1991), The efficiency of a flight muscle from the locust Schistocerca americana, The Journal of Physiology Т. 442 (1): 413–429, PMID 1798034, <http://jp.physoc.org/content/442/1/413.full.pdf+html> (недоступная ссылка)
- Heinrich, B. (1971), Temperature regulation of the sphinx moth, Manduca sexta. I. Flight energetics and body temperature during free and tethered flight, Journal of Experimental Biology Т. 54: 141–152, <http://jeb.biologists.org/content/54/1/141.full.pdf+html>
- Heinrich, B. (1970), Nervous control of the heart during thoracic temperature regulation in a sphinx moth, Science Т. 169 (3945): 606–607, DOI 10.1126/science.169.3945.606
- Harrison, Jon F.; Fewell, Jennifer H.; Roberts, Stephen P. & Hall, H. Glenn (1996), Achievement of thermal stability by varying metabolic heat production in flying honeybees, Science Т. 274 (5284): 88–90, PMID 8810252, DOI 10.1126/science.274.5284.88
- Heinrich, Bernd (1974), Thermoregulation in Endothermic Insects, Science Т. 185 (4153): 747–756, PMID 4602075, DOI 10.1126/science.185.4153.747
- Josephson, R.K. & Stevenson, R.D. (1991), The efficiency of a flight muscle from the locust Schistocerca americana, The Journal of Physiology Т. 442 (1): 413–429, PMID 1798034, <http://jp.physoc.org/content/442/1/413.full.pdf+html> (недоступная ссылка)
- Kammer, Ann E. (1968), Motor patterns during flight and warm-up in Lepidoptera, Journal of Experimental Biology Т. 48: 89–109, <http://jeb.biologists.org/content/48/1/89.full.pdf+html>
- Crespo, Jose G.; Goller, Franz & Vickers, Neil J. (2012), Pheromone mediated modulation of pre-flight warm-up behavior in male moths, Journal of Experimental Biology Т. 215: 2203–2209, doi:10.1242/jeb.067215, <http://jeb.biologists.org/content/215/13/2203.full.pdf+html>
- Crespo, Jose G.; Vickers, Neil J. & Goller, Franz (2013), Female pheromones modulate flight muscle activation patterns during preflight warm-up, Journal of Neurophysiology Т. 110: 862–871, doi:10.1152/jn.00871.2012, <http://jn.physiology.org/content/110/4/862.full.pdf+html>
- Crespo, Jose G.; Vickers, Neil J. & Goller, Franz (2014), Male moths optimally balance take-off thoracic temperature and warm-up duration to reach a pheromone source quickly, Animal Behaviour Т. 98: 79–85, doi:10.1016/j.anbehav.2014.09.031, <http://www.sciencedirect.com/science/article/pii/S0003347214003765>
- Clench, N. S. (1966), Behavioral thermoregulation in butterflies, Ecology Т. 47 (6): 1021–1034, DOI 10.2307/1935649
- Bartholomew, George A. & Heinrich, Bernd (1978), Endothermy in African dung beetles during flight, ball making, and ball rolling, Journal of Experimental Biology Т. 73: 65–83, <http://jeb.biologists.org/content/73/1/65.full.pdf>
- Heinrich, Bernd & Bartholomew, George A. (1979), Role of endothermy and size in inter- and intraspecific competition for elephant dung in an African dung beetle, Scarabaeus laevistriatus, Physiological Zoology Т. 52 (4): 484–496, <https://www.jstor.org/stable/10.2307/30155939>
- Ono, M.; Okada, I. & Sasaki, M. (1987), Heat production by balling in the Japanese noneybee, Apis cerana japonica as a defensive behavior against the hornet, Vespa simillima xanthoptera (Hymenoptera: Vespidae), Cellular and Molecular Life Sciences Т. 43 (9): 1031–1034, DOI 10.1007/BF01952231
- Lahondère, Chloé & Lazzari, Claudio R. (2012), Mosquitoes Cool Down during Blood Feeding to Avoid Overheating, Current Biology Т. 22: 40–45, DOI 10.1016/j.cub.2011.11.029