Пыльца
Пыльца́, устар. цветень, — скопление пыльцевых зёрен семенных растений[1]. Пыльцевое зерно представляет собой мужской гаметофит, развивающийся в микроспорангии из микроспоры и выполняющий функцию опыления, то есть оплодотворения женского гаметофита, находящегося в семязачатке.
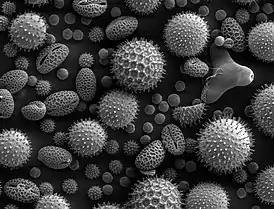

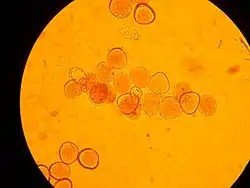
Строение
Пыльца развивается в пыльниках тычинок. Граничащий с наружной кожицей слой молодого пыльника перегородками делится на два слоя, из которых наружный производит стенку пыльцевой камеры, а внутренний даёт начало археспорию, состоящему из производящих, или материнских, клеток пыльцы. Затем материнские клетки пыльцы обычно увеличиваются в размере и утолщают оболочку. Они или остаются соединёнными между собой (большинство двудольных), или разъединяются (многие однодольные растения). Каждая производящая клетка затем делится на четыре специально производящие клетки или через повторное деление на два (у однодольных), или же вокруг получившихся путём деления четырёх ядер возникают сразу оболочки четырёх клеток (у большинства двудольных). Содержимое внутри каждой специальной производящей клетки облекается новой оболочкой, спородермой, дифференцирующейся на наружный толстый слой, экзину, и внутренний — интину. Спорам хвощей, папоротников и мхов свойственен ещё один слой над экзиной, называемый периной или периспорием. Наука, изучающая спородерму, называется палинологией, внутренняя же часть пыльцы изучается эмбриологией[2].
Большей частью получившиеся пыльцевые клетки вскоре разъединяются полностью, иногда же остаются соединёнными по четыре (тетрадами, или четвёрками) — например, у многих орхидей (тайник, гнездовка), y рогоза, анноны, рододендрона и др. У орхидей из трибы Orchideae пыльцевые клетки соединены в большом числе в пыльцевые тельца (лат. massulae), которые, в свою очередь, соединены между собой в одну массу, так называемый поллиний, заполняющую всю пыльцевую камеру. То же наблюдается у многих асклепиевых.
Величина пыльцевых клеток колеблется от 0,0025 до 0,25 мм. Они преимущественно эллипсоидальной или же шаровидной формы, иногда гранистые или угловатые. Внешний слой, экзина, часто бывает покрыт разнообразной скульптурой в виде гребешков, бугорков, шипов и т. д., иногда же сухой и гладкий. В спородерме может быть сквозное отверстие или легко растяжимое место, называемое апертурой. У пыльцы многих хвойных имеются воздушные мешки, являющиеся выростами спородермы и служащие для облегчения полёта пыльцы[2].
Пыльца как передатчик экологической информации у растений
По существующим в классической генетике представлениям, пыльца несёт только генетическую информацию, и количество пыльцы, попадающее на женский цветок, не играет роли, поскольку для оплодотворения достаточно одного пыльцевого зерна. В 1977 году В. Геодакян предположил, что количество пыльцы, попадающее на женский цветок, может нести также и экологическую информацию о ситуации в ареале[3][4]. Большое количество пыльцы говорит об оптимальных условиях среды (центр ареала, много мужских растений, хорошие условия для роста и погода), тогда как малое количество пыльцы, наоборот, несёт информацию о неблагоприятных условиях: это бывает либо на периферии, где сильно падает плотность популяции, либо в центре, при наступлении там экстремальных условий, которые элиминируют в первую очередь мужских особей.
Количество пыльцы может определять соотношение полов, дисперсию и половой диморфизм популяции растений. Большое количество пыльцы приводит к уменьшению этих характеристик и стабилизации популяции. Малое количество ведёт к их увеличению и дестабилизации популяции.
Зависимость вторичного соотношения полов от количества пыльцы было подтверждено у четырёх видов двудомных растений, принадлежащих к трём семействам, — Щавель кислый (Гречишные)[5][6], Дрёма белая (Гвоздичные)[7][8], Конопля посевная[9] и Хмель японский (Коноплёвые)[10][11]. С увеличением количества пыльцы уменьшается число мужских растений в потомстве и наоборот. Количество пыльцы зависит от плотности посадки растений, поэтому в густых посадках должна увеличиваться доля женских растений.
Зависимость разнообразия фенотипов потомства от количества пыльцы было обнаружено Д. Тер-Аванесяном в 1949 году у хлопчатника, вигны и пшеницы. У всех трёх видов при оплодотворении малым количеством пыльцы наблюдалось увеличение разнообразия потомства[12][13].
Роль пыльцы в жизни пчёл


После прохождения через пчелиные лапки пыльца становится обножкой, то есть пыльцой, которую пчёлы собрали и обработали своими ферментами.
Пыльца (пчелиная обножка) — это второй по объёму потребления и первый по значимости продукт питания пчелиной семьи. Пчёлы прикладывают немало усилий, чтобы запастись цветочной пыльцой — жизненно важным для них пищевым сырьём.
Пыльца как часть пищевой цепочки в животном мире — природный концентрат, содержащий белки, многие витамины, ценные минеральные вещества, а также полный набор незаменимых свободных аминокислот. По общему аминокислотному составу обножка близка к другим богатым белком пищевым продуктам — мясу, молоку, яйцам[14]. Внесение удобрений способствует увеличению количества белка в пыльце. У клевера красного в среднем за два года наблюдений в пыльце контрольных растений было 24,3 % белка, у мака восточного 33,4 %, а в пыльце растений под которые вносились удобрения, соответственно 30,8 и 36,4 %[15].
Благодаря пыльце масса вчерашней личинки возрастает в сотни раз всего за несколько дней, укрепляются и расправляются крылья, формируются все рабочие железы. Пыльца также служит сырьём для создания маточного молочка — продукта, предназначенного для кормления королевы-матки.
В пчелиной семье пыльца нужна в первую очередь пчёлам-кормилицам. Они интенсивно поедают этот белково-липидный корм, необходимый для выработки маточного молочка, которым питается молодая пчелиная матка, а в первые 3 дня также и личинки рабочих пчёл. Пыльцой питаются и только что родившиеся пчёлы: в их теле мало азота, они нуждаются в белках и витаминах. Пыльца нужна пчёлам-строителям для работы восковых желёз, трутням — для нормального полового созревания и функционирования. За сезон пчелиная семья собирает и потребляет 35—40 кг пыльцы обножки.
На поверхности тела пчёл находится антибиотическое вещество, которое оказывает ингибирующее действие на прорастание пыльцы и семян высших растений. В литературе есть сведения, доказывающие, что пыльца с тел и обножки быстро теряет всхожесть и для опыления не годится. При этом пыльца, попадающая на тело пчёл, погибает за разный промежуток времени. В опытах 1965 года внесение фосфорно-калийных удобрений способствовало сохранению жизнеспособности пыльцы клевера, находящейся на теле пчёл на 9,5—11 %[16]. Факт гибели пыльцы на теле пчёл представляет собой интересное явление природы. С одной, стороны, это противоречит целесообразности переноса пыльцы с помощью пчёл, а с другой, возможно, имеет значение в механизме отбора более сильного начала для будущего поколения[17].
Микробиота пыльцевых зёрен
Показано, что для пыльцевых зёрен разных видов растений характерен специфический набор бактерий, обитающих на их поверхности (микробиом)[18][19][20][21][22].
Организация микробиоты пыльцевых зёрен
Метагеномное изучение микробиоты пыльцы демонстрирует достаточно высокое разнообразие. Для четырёх видов растений — берёзы повислой (Betula pendula), рапса (Brassica napus), ржи (Secale cereale) и безвременника осеннего (Colchicum autumnale) — показано существование более тысячи различных видов бактерий, представляющих около ста семейств[21].
Существенно, что культивируемые в лабораторных условиях формы составляют меньшинство из всей микробиоты пыльцевого зерна (для указанных выше растений — 44 вида, то есть менее 5 %)[21]. Тем не менее, культивируемых бактерий на поверхности пыльцевых зёрен достаточно много — от 106 (берёза повислая) до 109 (безвременник осенний) клеток на грамм пыльцы[21].
Изучение поверхности пыльцевых зёрен с помощью сканирующей электронной микроскопии позволяет выявить колонии бактерий, организованные в том числе в виде биоплёнок[18][21].
Взаимодействие растений с собственной микробиотой
Показано, что растение взаимодействует с собственным микробиомом, синтезируя, в частности, антимикробные пептиды (англ. antimicrobial peptides, AMPs)[22]. Среди них наиболее изучены и, по-видимому, наиболее распространены дефензин-подобные пептиды. Так, у резуховидки Таля (Arabidopsis thaliana) из примерно 800 генов цистеин-обогащенных пептидов около трёхсот генов представляют собой именно гены дефензин-подобных пептидов[23]. Для дефензинов характерно наличие специфической третичной структуры с дисульфидным мостиком между остатками цистеина.
На важное значение антимикробных пептидов в пыльце указывает то, что 80 из 300 дефензин-подобных пептидов Arabidopsis thaliana гомологичны дефензин-подобному пептиду — PCP-A1, изолированному ранее из пыльцевой оболочки капусты (Brassica)[22][24]. Показано, что ряд антимикробных пептидов предотвращают самоопыление, взаимодействуя с рецепторами в тканях пестика[25], однако их влияние на микробиоту пыльцевого зерна изучено крайне мало.
У Arabidopsis thaliana показано, что по меньшей мере 200 из 800 генов цистеин-обогащённых пептидов экспрессируются в зародышевом мешке[26]. Предложена гипотеза, согласно которой данные пептиды обеспечивают противомикробный барьер для зародышевого мешка. Возможно, данный барьер изолирует зародышевый мешок от бактерий, способных проникнуть в него при росте пыльцевой трубки через завязь[22].
Следует уточнить, что также выявлены и функции некоторых антимикробных пептидов, не связанные с защитой от микроорганизмов и самоузнаванием при опылении. Изучен ряд пептидов, образующихся в клетке-синергиде и направляющих рост пыльцевой трубки; также определены антимикробные пептиды, инициирующие разрыв пыльцевой трубки при входе в зародышевый мешок[27].
Роль микробиома пыльцы во взаимодействии с животными
Остаётся плохо изученным взаимодействие микробиома пыльцы с опылителями, а также с иммунной системой человека при аллергической реакции на пыльцу[22].
См. также
Примечания
- Пыльца // Проба — Ременсы. — М. : Советская энциклопедия, 1975. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 21).
- Мейер Н. Р. Спородерма // Жизнь растений : в 6 т. / гл. ред. Ал. А. Фёдоров. — М. : Просвещение, 1978. — Т. 4 : Мхи. Плауны. Хвощи. Папоротники. Голосеменные растения / под ред. И. В. Грушвицкого, С. Г. Жилина. — С. 29—32. — 447 с. — 300 000 экз.
- Геодакян В. А. (1977) Количество пыльцы как регулятор эволюционной пластичности перекрестноопыляющихся растений. ДАН СССР, т. 234, № 6. с. 1460—1463.
- Геодакян В. А. (1978) Количество пыльцы как передатчик экологической информации и регулятор эволюционной пластичности растений. Жypн. общ. биологии. 39 № 5, с. 743—753.
- Correns С. (1922) Geschlechtsbestimmung und Zahlenverhaltnis der Geschlechter beim Sauerampfer (Rumex acetosa). «Biol. Zbl.» 42, 465—480.
- Rychlewski J., Kazlmierez Z. (1975) Sex ratio in seeds of Rumex acetosa L. as a result of sparse or abundant pollination. «Acta Biol. Cracov» Scr. Bot., 18, 101—114.
- Correns C. (1928) Bestimmung, Vererbung und Verteilung des Geschlechter bei den hoheren Pflanzen. Handb. Vererbungswiss., 2, 1—138.
- Mulcahy D. L. (1967) Optimal sex ratio in Silene alba. «Heredity» 22 № 3, 41.
- Riede W. (1925) Beitrage zum Geschlechts- und Anpassungs-problem. «Flora» 18/19
- Kihara H., Hirayoshi J. (1932) Die Geschlechtschromosomen von Humulus japonicus. Sieb. et. Zuce. In: 8th Congr. Jap. Ass. Adv. Sci., p. 363—367 (cit.: Plant Breeding Abstr., 1934, 5, № 3, p. 248, ref. № 768).
- Геодакян В. А., Геодакян С. В. (1958) Существует ли отрицательная обратная связь в определении пола? Журнал общей биологии. 46 № 2, с. 201—216.
- Тер-Аванесян Д. В. (1949) Тр. по прикл. ботанике, генетике и селекции. 28 119.
- Ter-Avanesian D. V. (1978) Significance of pollen amount for fertilization. «Bull.Torrey Bot.Club.» 105 N 1, 2—8.
- Хисматуллина Н. З. Апитерапия. — Пермь: Мобиле, 2005. — С. 74.
- Бухарева, 1971, с. 17—18.
- Бухарева, 1971, с. 17.
- Бухарева, 1971, с. 18.
- H. Colldahl, L. Nilsson. Possible relationship between some allergens (pollens, mites) and certain microorganisms (bacteria and fungi). A morphological study using the scanning electron-microscope // Acta Allergologica. — October 1973. — Т. 28, вып. 4. — С. 283–295. — ISSN 0001-5148.
- Robert R. Junker, Alexander Keller. Microhabitat heterogeneity across leaves and flower organs promotes bacterial diversity // FEMS Microbiology Ecology. — 2015-09-01. — Т. 91, вып. 9. — ISSN 0168-6496. — doi:10.1093/femsec/fiv097.
- Andrea Obersteiner, Stefanie Gilles, Ulrike Frank, Isabelle Beck, Franziska Häring. Pollen-Associated Microbiome Correlates with Pollution Parameters and the Allergenicity of Pollen // PLOS ONE. — 2016-02-24. — Т. 11, вып. 2. — С. e0149545. — ISSN 1932-6203. — doi:10.1371/journal.pone.0149545.
- Binoy Ambika Manirajan, Stefan Ratering, Volker Rusch, Andreas Schwiertz, Rita Geissler-Plaum. Bacterial microbiota associated with flower pollen is influenced by pollination type, and shows a high degree of diversity and species-specificity (англ.) // Environmental Microbiology. — 2016-12-01. — Vol. 18, iss. 12. — P. 5161–5174. — ISSN 1462-2920. — doi:10.1111/1462-2920.13524.
- Michael Zasloff. Pollen has a microbiome: implications for plant reproduction, insect pollination and human allergies (англ.) // Environmental Microbiology. — 2017-01-01. — Vol. 19, iss. 1. — P. 1–2. — ISSN 1462-2920. — doi:10.1111/1462-2920.13661.
- Kevin A. T. Silverstein, Michelle A. Graham, Timothy D. Paape, Kathryn A. VandenBosch. Genome Organization of More Than 300 Defensin-Like Genes in Arabidopsis (англ.) // Plant Physiology. — 2005-06-01. — Vol. 138, iss. 2. — P. 600–610. — ISSN 1532-2548 0032-0889, 1532-2548. — doi:10.1104/pp.105.060079.
- J. Doughty, S. Dixon, S. J. Hiscock, A. C. Willis, I. A. Parkin. PCP-A1, a defensin-like Brassica pollen coat protein that binds the S locus glycoprotein, is the product of gametophytic gene expression // The Plant Cell. — August 1998. — Т. 10, вып. 8. — С. 1333–1347. — ISSN 1040-4651.
- June B. Nasrallah. Recognition and Rejection of Self in Plant Reproduction (англ.) // Science. — 2002-04-12. — Vol. 296, iss. 5566. — P. 305–308. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.296.5566.305.
- Matthew W. Jones-Rhoades, Justin O. Borevitz, Daphne Preuss. Genome-Wide Expression Profiling of the Arabidopsis Female Gametophyte Identifies Families of Small, Secreted Proteins // PLOS Genetics. — 2007-10-12. — Т. 3, вып. 10. — С. e171. — ISSN 1553-7404. — doi:10.1371/journal.pgen.0030171.
- Tetsuya Higashiyama, Hidenori Takeuchi. The Mechanism and Key Molecules Involved in Pollen Tube Guidance // Annual Review of Plant Biology. — 2015-04-29. — Т. 66, вып. 1. — С. 393–413. — ISSN 1543-5008. — doi:10.1146/annurev-arplant-043014-115635.
Литература
- Бухарева Г. А. О пыльце и пчёлах // Пчеловодство : журнал. — 1971. — № 6. — С. 17—18.
- Сладков А. Н. Морфология пыльцы и спор современных растений в СССР в связи с методами её практического применения. — М.: Издательство Московского университета, 1962. — С. 256.
- Сладков А. Н. Введение в спорово-пыльцевой анализ. — М.: Наука, 1967.
- Косенко Я. Микромир пыльцы // Цветоводство. — 2008. — № 4. — С. 16—17.
- Пыльца // Словарь-справочник пчеловода / Сост. Н. Ф. Федосов. — М.: Гос. издат. сельскохоз. лит-ры, 1955. — С. 297—298. — 420 с.
- Пыльца // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
Ссылки
- База данных по морфологии пыльцы. Ботанический институт им. В. Л. Комарова РАН
- База данных австрийского общества — фотографии пыльцы растений около полутора тысяч видов, доступные после бесплатной регистрации
- Пыление (распространение пыльцы) крапивы на YouTube
| В библиографических каталогах |
|---|