Гусеница
Гу́сеница — личинка насекомых из отряда чешуекрылых (бабочек).
.jpg.webp)

.jpg.webp)
Тело состоит из головы, трёх грудных и десяти брюшных сегментов. Голова с мощно склеротизованными покровами, остальное тело у большинства мягкое, без крупных склеритов. На теле развиты щетинки, расположенные строго определённым образом, специфичным для отдельных семейств, родов и даже видов. Помимо этих первичных щетинок в старших возрастах у многих открытоживущих форм развивается густой покров из вторичных щетинок (наиболее мощный у гусениц представителей подсемейства медведиц). Размеры тела от нескольких миллиметров (у ряда молей) до 12—14 см (некоторые павлиноглазки)[1].
Для гусениц характерна парная шелкоотделительная железа, открывающаяся протоком на нижней губе. Выделяемый ею секрет при соприкосновении с воздухом образует шёлковую нить, используемую гусеницей для постройки кокона, изготовления защитной паутины, скрепления листьев и т. п.
На протяжении своей жизни гусеница интенсивно питается, запасая питательные вещества для последующего развития. Гусеницы большинства видов питаются зелёными частями растений. Некоторые развиваются внутри плодов; ветвей и стволов, питаясь древесиной; на лишайниках и на отмерших частях растений; на животных остатках (пух, перья, шерсть), а также на воске. Отдельные виды хищничают.
Стадия гусеницы может длиться от нескольких недель до нескольких лет. По мере роста гусеницы несколько раз линяют. По окончании своего развития гусеница превращается в куколку, из которой затем появляется взрослая бабочка.
В русском языке слово «гусеница» продолжает праслав. *ǫsěnica, образованное от *ǫsěnъ «волосатый», которое, в свою очередь, восходит к *ǫsъ «ус»[2][3]. Это слово в форме творительного падежа («гусѣницею») встречается в одной из древнейших русских летописей — Лаврентьевской летописи в записи под 1068 годом[4][5].
С гусеницами внешне сходны ложногусеницы — личинки насекомых семейства настоящих пилильщиков (Tenthredinidae). Название связано со значительным внешним сходством данных личинок с настоящими гусеницами бабочек. Часто ложногусеницами называют личинок всех семейств надсемейства пилильщиков (Tenthredinoidea) — группы семейств из отряда перепончатокрылых. В отличие от гусениц, у ложногусениц пилильщиков брюшные ноги развиты на II—X сегментах брюшка[6][7].
Строение
 |
|
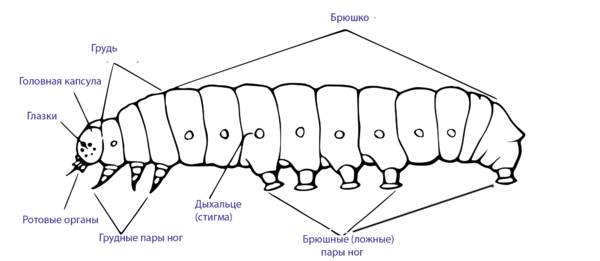 |
| Общее строение тела гусеницы, на примере Macroglossum stellatarum. | Строение тела гусеницы | |
Тело гусеницы состоит из головы, трёх грудных и десяти брюшных сегментов. Форма тела у различных видов довольно разнообразна. В большинстве случаев тело гусеницы более или менее цилиндрическое. Существуют варианты с постепенно утолщающимся (например Phytometrinae) или утончающимся (Hylophila) задним концом тела, суживающимся спереди или/и сзади, так называемая веретеновидная форма (серпокрылые моли). Также тело может быть несколько уплощённым (листовёртки, настоящие шелкопряды, моли выемчатокрылые). Более выраженными вариантами формы тела у гусениц являются сильное укорочение или утолщение тела с сильно суженными передним и задним концами, причём голова втянута в грудь, а ноги короткие — подобное строение характерно для голубянок и отчасти для пестрянок (мокрицевидная форма). Слизневидную форму тела имеют гусеницы слизневидок, которые могут быть совершенно округлыми, с плоской брюшной стороной; брюшные ноги у них могут отсутствовать, а голова быть целиком втянутой в грудь. Сильное сжатое тело в спинно-брюшном направлении, приводящее к плоской форме гусениц, отмечается у многих минирующих молей[8].
Голова

Голова образована плотной капсулой, состоящей из шести слившихся сегментов. Часто условно выделяют участки головы, занимающие сравнительно небольшую область между лбом и глазами, называемые щеками. С нижней стороны головы находится затылочное отверстие, которое в большинстве случаев имеет форму символа сердца[9].
По положению головы относительно тела различают следующие типы гусениц[10]:
- ортогнатический — продольная ось головы расположена более или менее перпендикулярно к оси тела, ротовые органы направлены вниз. Данный тип характерен для практически всех крупных гусениц, живущих открыто на растениях (булавоусые чешуекрылые, бражники, хохлатки, коконопряды, медведицы и многие другие).
- прогнатический — продольная ось головы совпадает с осью тела, ротовые органы направлены вперёд. Данный тип головы возник как приспособление к минирующему образу жизни. Он характерен для представителей беззубых молей, молей-малюток, Phyllocnistidae и ряда других семейств. Голова этого типа сильно уплощена, теменной шов отсутствует.
- полупрогнатический — данный тип занимает промежуточное положение между первыми двумя типами. Он характерен для гусениц, ведущих скрытный образ жизни.
Типичной формой головы гусениц является округлая. Иногда она может приобретать треугольную (многие бражники), прямоугольную (род орденские ленты) или сердцевидную форму. Фронтальная поверхность при этом становится плоской либо даже вдавленной. Теменные вершины могут значительно выдаваться над поверхностью тела, а иногда даже превращаться в большие «рога» или выросты (рода переливницы (Apatura) и Charaxes)[9].
Усики (антенны) гусениц короткие, трёхчлениковые. Располагаются они по бокам головы, между глазами и верхними челюстями, в так называемой усиковой впадине. Иногда усики подвергаются редукции — происходит сокращение общего числа члеников[11].
Ротовой аппарат гусениц грызущего типа. Верхние челюсти (мандибулы) всегда хорошо развитые и представляют собой сильно склеротизованные крепкие образования, варьирующие по своей форме. Их вершинный край обычно несёт на себе зубцы, служащие для откусывания или разрезания пищи. На внутреннем крае иногда могут находиться бугры, необходимые для разжёвывания пищи. Нижние челюсти (максиллы) и нижняя губа (лабиум), как и у многих других насекомых с полным превращением, слиты в единый лабио-максиллярный комплекс. Слюнные железы видоизменены в шёлкоотделительные[12].
Органы чувств

Органами зрения у гусениц являются глаза, которые представлены отдельными простыми глазками, находящимися по бокам головы. В большинстве случаев они располагаются в виде дугообразного ряда, состоящего из пяти простых глазков, и одного, стоящего внутри этой дуги. Иногда может наблюдаться их примитивность, либо, наоборот, специализация. Так, у новозеландских гусениц рода Sabatinca глаза образованы из пяти простых глазков, слившихся в один сложный глаз[13].
Простые глазки гусениц, вероятно, содержат только два светочувствительных пигмента, которые обеспечивают возможность различения цветов. Данный вывод согласуется с поведением гусениц дневных бабочек из родов ванессы и огородных белянок. Во время периода активного питания зелёные, жёлтые и оранжевые цвета привлекают гусениц, а во время периода окукливания привлекательными для них становятся тёмные тона: коричневый и чёрный[14]. Гусеницы способны различать форму и размер предметов. В экспериментах, из двух одинаковых по ширине вытянутых фигур прямоугольной формы, гусеницы монашенки (Lymantria monacha) предпочитают более высокую, также сужающаяся кверху фигура привлекает их больше, чем фигуры с теми же высотой и площадью, но суживающиеся книзу или имеющие параллельные края[14].
Орган вкуса представлен вкусовыми сенсиллами на максиллах. Исследования специфичности вкусовых сенсилл показали большое разнообразие количества специфических чувствительных клеток и широкий спектр распознаваемых ими химических соединений, многие из которых имеют сигнальное значение при распознавании гусеницами своего кормового растения. Большое количество специфических рецепторных клеток отражает разнообразие пищевых субстратов и приспособленность гусениц к переходу при питании с одного вида растений на другие[14].
На антеннах гусениц (экспериментально доказано для табачного бражника, репницы и Heliotis zea) имеются холодовые рецепторы и специализированные рецепторы влажности — чувствующие клетки, реагирующие на изменение влажности окружающего воздуха[14].
Грудь и брюшко
_Fausses_pattes.jpg.webp)
.jpg.webp)
Тело гусеницы заключено в мягкий перепончатый покров, благодаря чему обладает чрезвычайной подвижностью. Склеротизованными участками являются тергиты переднегруди и десятого брюшного сегмента. Каждый сегмент тела гусеницы может быть разделён на ряд вторичных колец, отделённых друг от друга бороздками, которые внешне не отличаются от границ настоящих сегментов. Переднеспинка (переднегрудной щит) крайне редко занимает весь тергит, и у большинства гусениц отделена от него небольшим склеритом, расположенным перед дыхальцем (стигмой) и носящим название предстигмального щитка. Средне- и заднеспинка никогда полностью не склеротизованы, а их боковые участки всегда разделены на несколько отдельных склеритов. Тергиты брюшных сегментов также всегда разделены на несколько склеритов[15].
Анальное отверстие располагается на последнем сегменте тела и окружено 4 лопастями. Одновременно хорошо развиты не все из них. Верхняя (наданальная) лопасть свешивается над анальным отверстием. Нижняя (поданальная) лопасть часто представлена в виде толстого мясистого образования конической формы; пара боковых (прианальных) лопастей — парапрокты — обычно хорошо развиты у гусениц пядениц и хохлаток и имеют вид относительно больших выростов со щетинками на концах[16].
Почти все гусеницы относятся к группе с одной закрытой стигмой (дыхальцем) на груди. Исключением являются отдельные виды, ведущие водный образ жизни. У них стигмы являются замкнутыми, и их заменяют трахейные жабры. Грудь гусениц несёт только одну открытую функционирующую стигму. Второе редуцированное дыхальце находится между средне- и заднегрудью. Грудное дыхальце обычно крупнее брюшных. Брюшко на I—VIII сегментах несёт восемь пар стигм, расположенных ниже грудной стигмы и более или менее посередине сегмента либо несколько ближе к его переднему краю. Стигма восьмого сегмента располагается выше остальных брюшных и крупнее их, в то время как стигма первого сегмента, наоборот, располагается несколько ниже остальных. По форме стигмы могут быть круглыми или овальными[17].
Конечности
.jpg.webp)
У большинства гусениц развиты три пары грудных ног (по паре на каждом из сегментов груди) и пять пар ложных брюшных ног на III—VI и X сегментах брюшка. Брюшные ноги несут мелкие крючочки, расположенные у представителей разных групп чешуекрылых по-разному — в виде круга, продольных или поперечных рядов. Нога состоит из пяти члеников: тазика, вертлуга, бедра, голени и лапки. Грудные ноги гусениц в некоторой мере редуцированы по сравнению с истинными ходильными ногами, и функцию передвижения несут преимущественно брюшные ноги. На конце грудной лапки находится неподвижно сочленённый с ней коготок, имеющий разную длину и форму. Конечной частью брюшной ноги является подошва, которая может втягиваться и выпячиваться, и несёт на своём конце коготки[18].
Существуют два типа строения подошвы[18]:
- край имеет более или менее округлое очертание с расположенными по периферии в форме кольца крючками, втягивающая мышца прикреплена в центре подошвы;
- внешняя часть подошвы редуцирована, и крючки располагаются только по её внутреннему краю, мышцы прикреплены у внешнего края подошвы; в этом случае наружный край подошвы, противостоящий краю, несущему крючки, иногда довольно сильно склеротизован.

В разных группах бабочек описаны отклонения от описанного варианта расположения ног. Наиболее известное — гусеницы пядениц, большинство из которых имеют только две пары брюшных ног (на VI и X сегментах). В результате они перемещаются как бы «шагая». Русское название, как и немецкое (нем. Spannern), происходит от сходства передвижения гусеницы с движениями кисти руки человека, измеряющего длину пядью. Латинское название семейства пядениц — Geometridae (от латинизированного греческого «землемер») также дано им в связи с этой особенностью. Брюшные ноги могут быть редуцированы на III и IV сегментах брюшка у гусениц некоторых совок (Noctuidae). У некоторых гусениц описано более пяти пар брюшных ног. У зубатых молей (Micropterigidae) — восемь, у мегалопигид (Megalopygidae) — семь (со II по VII и на X сегменте), у представителей рода Stigmella из семейства моли-малютки (Nepticulidae) — шесть (со II по VII сегменты) пар. Кроме того, ноги (как брюшные, так и грудные) могут быть полностью редуцированы у мелких минирующих чешуекрылых. У гусениц некоторых родов в брюшном отделе отсутствуют конечности, но имеются т. н. ложные или брюшные ноги[18].
Покровы тела и их придатки

.jpg.webp)
Тело гусеницы почти никогда не бывает полностью голым и обычно покрыто разнообразными образованиями, которые можно разделить на кутикулярные выросты, волоски и выросты тела[19].
Кутикулярные выросты представляют собой скульптурные элементы и небольшие выросты кутикулы: шипики, гранулы, звёздчатые образования, которые могут иметь вид мелких волосков — хетоидов. Волоски, щетинки и их производные отличаются от скульптурных элементов своим сочленением с кутикулой и развитием за счёт специальных клеток гиподермы. Основание волоска окружено кольцевым валиком, либо же сам волосок находится в углублении. В них волоски торчат свободно, в связи с чем легко выпадают или обламываются Условно волоски разделяют на собственно волоски и щетинки. Последние обычно развиты на теле гусениц и расположены строго определённым образом, специфичным для отдельных семейств, родов и даже видов. С точки зрения систематики щетинки и волоски гусениц имеют примерно такое же значение, как строение копулятивных органов у взрослых особей[19]. Например, форма вызывающих жгучие раздражения щетинок отличает гусениц из семейств Megalopygidae (мелкие щетинки разбросаны по всему телу) и павлиноглазки (Saturniidae: щетинки собраны в нескольких «веточках», иногда напоминающих хвоинки)[20]. Волоски чрезвычайно различны по своей форме. В большинстве случаев они представлены нитевидными или щетинковидными образованиями, иногда могут иметь боковые придатки[21].
Выросты кожи тела — образования, состоящие из выпячиваний кожи и имеющие внутри себя полость, сообщающуюся с полостью тела. К ним относятся бугорки — различные образования, связанные с первичными щетинками. Бородавка — выступ, покрытый пучком щетинок или волосков. Бородавки бывают сферическими или, наоборот, уплощёнными и овальными, часто очень крупными, например, у волнянок. Характерные выросты представляют также колючки. Волоски на теле гусениц могут принимать участие в терморегуляции. Так, что волоски на теле гусениц непарного шелкопряда эффективно снижают потери тепла, при этом не препятствуя поглощению солнечной радиации[19]. У гусениц бражников на последнем сегменте обычно имеется т. н. «рог»[22].
В редких случаях у гусениц, ведущих водный образ жизни, на теле развиваются трахейные жабры. Обычно они имеются на всех сегментах тела (кроме переднегруди и X сегмента брюшка) в виде пучков нежных нитей с входящими в них трахеями. Стигмы в этих случаях замкнуты[19].
Мягкая кутикула гусениц является складчатой и неплотно прилегает к телу. Благодаря этой особенности они могут расти между линьками, но только до тех пор, пока складки кутикулы не растянутся и тело гусеницы не заполнит весь объём наружного скелета[23].
Окраска гусениц играет важную роль в повышении их выживаемости. Пигменты, обуславливающие окраску, обычно находится в кутикуле хитинового покрова, либо в гиподерме — слое, находящемся под ней. Пигменты могут откладываться по-разному. Например, у гусениц бражников кутикула бесцветная и пигменты содержатся в гиподерме[24], у махаона яркие красные и чёрные элементы окраски гусеницы обусловлены скоплениями пигментов именно в кутикуле, а жёлтый пигмент сконцентрирован в гиподерме[24]. Основными элементами в окраске гусениц являются пигментные пятна и точки, а также полосы, проходящие вдоль тела. Точки могут увеличиваться и сливаться между собой, образуя различные пятна. Продольные пятна могут распадаться на вторичные отдельные. При слиянии частей продольных полос могут образовываться поперечные полосы, называемые перевязями. У гусениц бражников, берёзовых шелкопрядов и некоторых других имеется характерные косые полосы, каждая из которых пересекает по два сегмента[25].
В покровах гусениц имеются одноклеточные кожные железы. Они располагаются возле дыхалец и функционируют только при линьке — их выделения облегчают отслаивание старых кожных покровов[26].
Питание

.jpg.webp)
Большинство гусениц является фитофагами — они питаются листьями, цветками и плодами различных, преимущественно цветковых, реже — хвойных, растений[27][28]. Гусеницы целого ряда родов и видов, например пяденицы папоротниковой[29], совки Callopistria floridensis[30] и совки папоротниковой, тонкопряда орлякового питаются на листьях папоротников[31], а гусеницы некоторых видов пёстроногих молей питаются их спорангиями[32]. Малочисленное количество видов питаются мхами, например первичные зубатые моли, эребеиды рода Sinarella, моховая выемчатокрылая моль[33].
Всех растительноядных гусениц можно разделить на три группы: полифаги, олигофаги и монофаги. Наименее специализированная группа — гусеницы-полифаги, являющиеся многоядными и способными питаться различными видами и родами растений. Гусеницы-олигофаги кормятся растениями, принадлежащими к одному семейству или роду. Например, гусеницы поликсены питаются только четырьмя видами рода кирказон, а гусеницы махаона — исключительно зонтичными. Монофаги являются наиболее специализированной группой и способны питаться только лишь одним видом кормового растения. Например, гусеницы тутового шелкопряда питаются исключительно листьями шелковицы. Среди гусениц ночных бабочек полифагия встречается очень часто, а среди дневных бабочек — крайне редко (в качестве примера последних можно рассматривать репейницу и боярышницу). Большинство видов принадлежит к группе олигофагов[34].
Представители немногих семейства (стеклянницы, древоточцы, некоторые кастнии) являются ксилофагами и питаются древесиной, прогрызают ходы в стволах и ветвях древесных или корнях травянистых растений[32][35][36]. В коре молодых ясеней и маслин развивается гусеница огнёвки Euzophera pinguis, питаясь слоями луба и заболони[37]. Гусеницы пробковой моли развиваются во влажной гнилой древесине, на трутовых грибах, также известны случаи их развития в стенках деревянных колодцев, деревянных бочках и пчелиных ульях[38], в старых гнёздах германской и обыкновенной ос[39].
Переходом от растительноядных к нерастительноядным являются гусеницы, питающиеся лишайниками (лишайницы, Cleorodes lichenaria, род совок Bryophila)[40], и прежде всего грибами. Например, практически исключительно трутовиками питаются гусеницы эребеиды рода Parascotia[40]. Грибоядные гусеницы свойственны для семейства настоящие моли и, в особенности, для рода Tinea. Единственная гусеница, питающаяся шляпочными грибами является африканская мешочница Scirpophaga marginepunctella[40]. Интересен факт, что гусеницы амбарной моли могут развиваться на ядовитой спорынье[40].
Ряд видов является кератофагами и питается веществами животного происхождения: кожей, волосами, шерстью, роговыми веществами. Примерами служат представители семейства настоящие моли: платяная моль, ковровая моль (Trichophaga tapetiella), шубная моль, меховая моль и другие[40]. Гусеницы молей рода Ceratophaga живут в рогах живых и мёртвых африканских антилоп и буйволов, питаясь кератином[41]. А гусеницы вида Ceratophaga vicinella, обитающего в южной Флориде, питаются панцирями мёртвых черепах гофер-полифем[42].
Воском питаются гусеницы представителей семейства настоящие огнёвки. К ним относятся пчелиные огнёвки — большая восковая моль, малая восковая моль, восковая огневка и другие представители этого подсемейства — Melissoblaptes и Lamoria. В начале развития в восковых сотах медоносных пчёл гусеницы указанных бабочек питаются мёдом и пергой, а затем переходят к питанию восковыми рамками, смешанными с остатками коконов (чистый воск для питания не пригоден — в нём не хватает питательных веществ). Реже они встречаются гнёздах шмелей и общественных ос[43].
Гусеницы нескольких видов из группы Chrysauginae (семейство Огнёвки настоящие), как то виды Cryptoses choloepi, Bradipodicola hahneli, являющиеся комменсалами трёхпалых ленивцев[44] (форезия), питаются их экскрементами (копрофагия)[45].


Следующей группой являются хищные гусеницы, среди которых прослеживаются все переходы от некрофагии к паразитизму и хищничеству. Случайное хищничество и каннибализм, особенно в условиях совместного проживания и в случае недостатка нормальной пищи, отмечается весьма часто. Преимущественно при этом поедаются ослабленные и больные гусеницы (хлопковая совка, многие медведицы). Некоторые растительноядные виды могут являться факультативными хищниками и нападать на других гусениц, и при достаточном количестве растительной пищи, например совки Scopelosoma satelitia и виды рода Calymnia[44].
Собственно хищники встречаются среди гусениц сравнительно редко[44]. Гусеницы отдельных видов совок (Eublemma scitulum), узкокрылых молей (Coccidiphila gerasimovi), один вид малинных молей (Stathmopoda melanchra), некоторые представители Heliodinidae питаются червецами. Также отмечены случаи хищничества гусениц некоторых видов пёстроногих молей в колониях червецов и щитовок[32]. Гусеница совки Cosmia trapezina питается растениями, живёт между двумя сплетенными вместе листьями и из этого укрытия также нападает на голых гусениц разных видов[46].
Примерно 300 видов голубянок считаются энтомофагами (или предположительно, энтомофагами) на последних стадиях развития их гусениц. Они питаются равнокрылыми насекомыми и личинками муравьёв, находящимися с ними в мирмекофильной ассоциации. Это в основном (около 90 %), представители подсемейства Miletinae и родов Lepidochrysops, Maculinea, Phengaris (Polyommatinae), и лишь около 40 видов из других таксономических групп голубянок. Равнокрылыми насекомыми питаются гусеницы голубянок родов Feniseca, Logania, Miletus, Spalgis, Taraka, Thestor и другие[47]. Случаи питания тлями отмечены у гусениц голубянок видов Spalgis pius из Индии и Feniseca tarquinius из Северной Америки[48]. Гусеницы последней питаются тлями родов Neoprociphilus, Pemphigus, Prociphilus и Schizoneura[49]. Гусеницы пядениц рода Eupithecia, обитающие на Гавайских островах, являются насекомоядными и обладают целым арсеналом приспособлений для поимки своей добычи[50][51]. Гавайские гусеницы 4 видов роскошных молей из рода Hyposmocoma, например Hyposmocoma molluscivora, используют шёлковые ловушки, чтобы ловить улиток (преимущественно рода Tornatellides), которыми они питаются[52].
Интересны крайне редкие случаи настоящего паразитизма[44]. Гусеницы представителей семейства Epipyropidae живут исключительно на цикадках семейства фонарниц, где проводят весь цикл своего развития, иногда даже окукливаясь на них же[53]. Гусеницы паразитического вида огнёвок Sthenobaea parasitus из Бразилии живут среди колючек гусениц родов Automeris и Dirphia, встречаясь по несколько особей на одной гусенице-хозяине[54][44]. Оплетая паутинные нити между колючками боковых рядов, они сооружают подобие галереи и питаются колючками, отчего гусеница-хозяин погибает[55]. У гусениц Microfrenata отмечен паразитизм на личинках муравьиного льва Myrmeleon europaeus[44].
В 2014 году способность поедать полиэтилен и расщеплять его с помощью симбиотических бактерий пищеварительного тракта (Enterobacter asburiae и Bacillus) была обнаружена у гусениц Plodia interpunctella из семейства настоящих огнёвок[56]. В 2016 году аналогичная способность была выявлена у гусениц пчелиной огнёвки из того же семейства[57][58].
Вылупившаяся гусеница, как правило, сперва поедает остатки яйца из которого она появилась, а затем, спустя какое-то время, приступает к поеданию своего кормового субстрата[34]. Пищеварительный тракт гусениц соединяется с остальным телом только у переднего и заднего концов, благодаря чему, вероятно, движение остального туловища не мешает гусеницам переваривать пищу[59]. В слюнных железах гусениц выделяются три основные группы пищеварительных ферментов — протеазы, карбогидразы и липазы[60].
Жизненный цикл

Жизненный цикл чешуекрылых состоит из 4 фаз: яйца, гусеницы, куколки и имаго (взрослого насекомого)[61]. После спаривания, самки бабочек откладывают яйца. Стадия яйца у различных видов может длиться от нескольких дней в тёплое время года до нескольких месяцев в том случае, если яйца зимуют. По мере развития внутри яйца происходит формирование гусеницы, которая затем прогрызает оболочку и выходит наружу. У некоторых видов сформированная гусеница зимует внутри яйца и выходит из него только с наступлением весны. У многих видов гусеницы сразу же после выхода съедают оболочку своего яйца[61].
В зависимости от вида, развитие гусеницы может продолжаться от нескольких дней до нескольких лет (у высокогорных и полярных видов). От продолжительности развития гусениц будет зависеть и число генераций (поколений) за год. Гусеницы бабочек, живущих в северных широтах, порой не успевают завершить своё развитие за одно лето, из-за чего вынуждены впадать в зимнюю диапаузу до следующего лета. К примеру, обитающая за полярным кругом в Гренландии и Канаде волнянка Gynaephora groenlandica известна тем, что её стадия гусеницы может длиться до 14 лет (обычно продолжительность стадии гусеницы составляет до 7 лет)[62]. Вторая уникальная особенность гусениц данного вида заключается в том, что во время зимовок в стадии диапаузы они могут выдерживать температуру до −70 °C[62]. При снижении температуры окружающей среды в конце арктического лета гусеницы начинают синтезировать защищающие их от холода химические соединения, такие как глицерин и бетаин[62]. Гусеницы проводят около 90 % своей жизни в замёрзшем виде в состоянии диапаузы и лишь около 5 % — питаясь скудной растительностью. Развитие гусениц родственного вида — волнянки Кузнецова — эндемика острова Врангеля, длится до 6 лет[63].
Гусеницы высокогорных видов также могут завершать своё развитие за несколько сезонов. Так, некоторые высокогорные бабочки рода парнассиусы (Parnassius), например многочисленные подвиды аполлон Чарльтона летают только в чётные или нечётные годы, так как их гусеницам для полного завершения своего развития требуется два сезона[64].
Возрастные изменения

Гусеницы, как и другие голометаболические личинки, живущие в совершенно других, чем взрослое насекомое, условиях обитания, эволюционировали самостоятельно и в некоторой степени независимо от стадии имаго. Это вызвано резким разграничением функций между данными фазами жизненного цикла. Гусеницы одного и того же вида, но различного возраста часто отличаются существенными различиями между собой. Например, некоторые морфологические структуры развиваются только в определённом возрасте. Строение гусеницы первого возраста не всегда является первичным по отношению к гусенице последнего возраста. Фазы развития различных видов могут быть неэквивалентными. Например, третий возраст одного вида по своему развитию может соответствовать четвёртому у другого вида и т. п[65].

Оценка возрастных изменений гусениц была произведена американским энтомологом Мэри Ливингстон Риплей (Mary Livingston Ripley, 1914—1996). На основании этих исследований постэмбриональные изменения гусениц могут быть разделены на три группы: рекапитулятивные, нерекапитулятивные и сложные. Примером рекапитулятивных изменений является наличие первичных щетинок у гусениц первого возраста и т. п. Нерекапитулятивные (функциональные) могут быть вызваны адаптацией к различным функциям, или необходимостью роста самой гусеницы. Примером является редукция первых пар брюшных ног у молодых гусениц; а также случаи гиперметаморфоза. Сюда же можно отнести явления неравномерного роста — увеличение относительной величины головы, глаз, крючков брюшных ног (все эти структуры растут медленнее других частей тела, и у гусениц ранних возрастов они относительно крупнее, чем у взрослых). Комплексное воздействие различных факторов вызывает сложные изменения в одном или в разных направлениях[65].
Степень возрастных изменений находится в непосредственной зависимости от принадлежности гусеницы к той либо иной систематической группе. Данные изменения могут быть, как самыми незначительными, выражающимися в величине и окраске тела (например у многих совок, настоящих огнёвок), так и существенными, например превращение почти голой гусеницы в волосатую (медведицы) или шипоносную (нимфалиды). Либо же наоборот, превращение гусениц, несущих на себе различные выросты в молодом возрасте, в голых на взрослых стадиях развития. Наиболее значительными являются данные изменения при гиперметаморфозе[65].
Голова гусениц первого возраста всегда является непропорционально большой относительно величины всего тела, что объясняется неравномерным ростом различных тканей. С возрастом это соотношение изменяется[66]. Почти всегда гусеницы первых возрастов имеют голый покров тела и могут иметь на себе только первичные щетинки. Начиная со второго возраста покровы тела гусеницы резко изменяются по мере её приближения ко взрослому состоянию. Примером наиболее выраженных возрастных изменений являются гусеницы представителей групп волнянки, медведицы, ложные пестрянки. Примерами возрастных изменений служат колючки гусениц нимфалид и выросты гусениц слизневидок. Сами щетинки, волоски и выросты с возрастом гусеницы также могут сильно изменять свою форму и размер[67].
Линька
При своём развитии гусеница проходит несколько фаз, обычно называемых «возрастами», которые часто могут отличаться друг от друга не только окраской и величиной, но и структурными признаками. Наиболее часто наблюдаются выраженные различия между первой и второй фазами. Порой же различия между отдельными возрастами гусениц являются ничтожными. Вышедшая из яйца гусеница на протяжении своей жизни претерпевает несколько линек, количество которых зависит в первую очередь от её принадлежности к тому или иному биологическому виду. Обычным числом линек принято считать четыре, хотя наличие в цикле развития пять и даже семь линек встречается довольно часто. Наименьшее число линек отмечено у гусениц-минёров и составляет две[68].
Число линек даже у одного и того же вида порой подвержено колебаниям, причины которых пока мало изучены. Одним из влияющих факторов выступает окружающие условия; например, зимующее поколение вида может иметь одну дополнительную линьку по сравнению с летним. Неблагоприятные условия в период роста вызывают увеличение количества линек. Наиболее разительным примером является платяная моль, чьи гусеницы линяют от 4 до 40 раз. В некоторых случаях число линек различно у разных полов. Так, самки могут иметь дополнительные линьки (волнянки), что связано с резким половым диморфизмом, аналогичный случай отмечен у совки Coenurgia erechtea, с не выраженным половым диморфизмом[68][69].

К моменту линьки гусеница перестаёт питаться, становится неподвижной, прячется в укрытия и защищённые места. Здесь она выстилает своим шёлком субстрат. Затем прикрепляется к нему крючками брюшных пар ног. Гусеницы мешочниц перед процессом линьки прикрепляют шёлковой нитью свой чехлик к окружающему субстрату. Известны случаи плетения специального «кокона для линьки» у молей из рода Bucculatrix и совки Parastichtis oblonga[68].
Гусеницы перед началом процесса линьки характеризуются наличием целого ряда отличий в своём внешнем виде: их кожные покровы становятся сильно растянутыми, а голова относительно мала. К этому моменту голова гусеницы следующего возраста, постепенно вытягивается из старой головной капсулы, и располагается в передней части груди. При линьке сбрасываемые покровы тела гусеницы разрываются на тергитах груди, а головная капсула отрывается и отпадает. Через разрыв в покровах тела гусеница освобождается от своих старых покровов. Многие гусеницы съедают их после линьки[68].
Интересны особенности линьки зимующих гусениц пестрянок рода Zygaena. Перед зимовкой у них имеется дополнительная линька; при этом «зимние покровы тела» существенно отличаются от «покровов роста»: в отличие от нормальных линек, всегда сопровождающихся увеличением размеров головы гусеницы, при зимней линьке голова у них всегда уменьшается. Этот случай напоминает явление гиперметаморфоза, наблюдённого у некоторых видов[68].
Окукливание

Окукливанием называется линька, при которой гусеница превращается в куколку. Это предпоследняя линька в цикле развития бабочек; при последней линьке куколка превращается во взрослое насекомое (имаго). Перед линькой на куколку гусеница переходит в состояние предкуколки, или пронимфы. Поскольку снаружи пронимфа покрыта ещё не сброшенной личиночной кутикулой, она сохраняет внешний вид гусеницы, но при этом под её личиночной кутикулой происходит формирование куколочного строения. Как и перед всякой линькой, гиподерма отслаивается от старой (в данном случае личиночной) кутикулы и меняет свою форму, приобретая черты новой стадии — куколки. Перед окукливанием гусеница перестаёт питаться. Многие гусеницы становятся беспокойными и начинают ползать в поисках подходящего места для окукливания. Часто в поисках таких мест некоторые гусеницы могут перемещаться на значительные расстояния (например, коконопряд малинный, бражник прозерпина)[70][34].
Место и вид окукливания очень разнообразны. Инстинктивно многие гусеницы избирают для окукливания недоступные для хищников и скрытые места, другие же окукливаются открыто. Многие гусеницы окукливаются прямо на своём кормовом растении, например белянки, отдельные виды Zygaena и совок. Большинство бражников, шелкопрядов и совок окукливаются в земле — даже виды, живущие на деревьях. Перед окукливанием они спускаются по стволу или опускаются на шелковине. У Microlepidoptera наблюдается большое разнообразие способов окукливания, многие их гусеницы покидают места питания[71].
У гусениц, окукливающихся в почве или на её поверхности, куколка часто просто покоится в немного углублённой ямке. В простых ямках располагаются куколки бражников, шелкопрядов и совок. При проникновении в почву гусеницы скрепляют шелковинной нитью вокруг себя отдельные кусочки земли и песчинки, сооружая своеобразную куколочную колыбельку, сходную с таковой у окукливающихся жуков. Гусеницы многих видов, преимущественно ночных бабочек, перед окукливанием сооружают кокон, который может отличаться большим разнообразием у представителей различных групп видов. Часто он сооружается из слабо скреплённых комочков земли либо же из древесной трухи у гусениц, развивающихся внутри древесины (различные древоточцы и стеклянницы). Гусеницы лишайниц и медведиц сооружают неплотные коконы из своих волосков — сперва гусеница делает основу из нескольких шелковинных нитей, а затем плетёт более плотные стенки. Пестрянки рода Zygaena сплетают плотные коконы, кажущиеся сделанными из крепкой ткани с характерным блеском. Некоторые виды шелкопрядов и павлиноглазок сооружают своеобразные бочкообразные либо яйцевидные коконы, изнутри выстланные как бы пергаментом, а снаружи сплетённые с волосками гусеницы[71].
Образование шёлка
Все гусеницы умеют выделять шёлковое волокно (нить). Большинство использует его для прикрепления к субстрату при передвижении. Гусеница, ползущая по растению или по почве, постоянно оставляет за собой тонкую шёлковую дорожку. Если она упадёт с ветки, то останется висеть на шёлковой нити. Гусеницы некоторых семейств молей и огнёвок строят из шёлка туннели (шелковинные ходы). Каждый, кто видел повреждения, нанесённые гусеницами настоящих молей меховым или шерстяным изделиям, замечал шелковинные ходы в подшёрстке или на поверхности вязаных вещей. Мешочницы и некоторые другие используют шёлковую нить как основу для изготовления переносного чехлика. Гусеницы горностаевых молей и некоторых хохлаток строят шелковинные гнёзда на кормовых растениях. В некоторых семействах, например, у коконопрядов, павлиноглазок и настоящих шелкопрядов гусеница перед окукливанием строит шёлковый кокон[72].

Прядильный аппарат
Прядильный аппарат состоит из прядильного сосочка и несущего его склерита. Прядильный сосочек представляет собой трубку, верхняя стенка которой обыкновенно короче нижней, концевой край неровный. Края прядильного сосочка иногда обрамлены бахромой. Шелковыделительный проток, проходящий сквозь прядильный сосочек, открывается на его дистальном конце. В очень редких случаях, например у первичных зубатых молей и некоторых минёров, прядильный сосочек, по-видимому, отсутствует[73].
Прядильный сосочек чрезвычайно изменчив по форме и длине у представителей различных групп. Существует тесная связь между строением прядильного сосочка и шелкоотделительной деятельностью гусениц. Гусеницы, оплетающие свои ходы, например тонкопряды и большинство Microfrenata, имеют длинный, тонкий и цилиндрический прядильный сосочек. Наоборот, короткий и уплощённый прядильный сосочек встречается только у гусениц, которые не плетут коконов или шелкоотделительная деятельность которых ограничена, например у бражников, многих совок и минёров[73].
В развитии шелкоотделительных желёз гусениц наблюдаются некоторые особенности. В последние 4 дня жизни гусеницы, когда она ещё питается, железа развивается очень быстро и за короткие сроки достигает своего максимального веса. Через сутки после начала плетения кокона вес железы резко уменьшается, а затем продолжает уменьшаться дальше, вплоть до окончания плетения кокона гусеницей. Клетки, вырабатывающие шёлк, синтезируют его, видимо, за счёт накопленных веществ. У дубового шелкопряда плетение кокона находится в зависимости от влажности окружающего воздуха — так в атмосфере с повышенной влажностью, гусеницы не плетут кокон[73].
Химический состав и структура шёлка
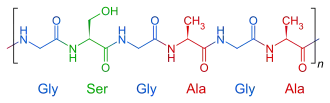
Шёлк образован белками — фиброином (75 %) и серицином, образующим поверхностный слой шёлкового волокна. Фиброин выделяется задней, а серицин — средней частью железы. Данные белки сильно отличаются друг от друга по своему аминокислотному составу. Серицин отличается от фиброина меньшим содержанием в своём составе гликокола, аланина и тирозина и значительно большим содержанием серина и диаминокислот. Шёлковое волокно сверху покрыто восковой пелликулой, по химическому составу сходной с кутикулином (белок, основной компонент наружного слоя кутикулы — эпикутикулы)[74][75].
Шёлковые волокна, выделяемые разными расами тутового шелкопряда, характеризуются различным химическим составом. Шёлковые волокна павлиноглазок Antheraea mylitta и Samia cynthia содержат меньше гликокола, чем волокна тутового шелкопряда. Шелковина мешочницы Oeceticus platensis не содержит в своём составе тирозин. Муравьиная кислота (40 %) входит в состав секрета прядильных желёз гусениц большой гарпии, из которого они сплетают прочный малопроницаемый кокон[74].
Образование волокна
Шёлковое волокно выходит из отверстия одной из губных желёз гусеницы, проходит через общий выводной проток этой железы. После этого оно проходит через «прессующий аппарат», который сжимает его и придаёт форму ленты. В то время, как два волокна, выделенных парными железами, проходят через общий выводной проток, добавочные железы производят клейкое вещество, которое склеивает их вместе. Механизм затвердевания шёлка мало изучен. Известно, что данный процесс осуществляется не путём высыхания, так как процесс затвердевания происходит даже в воде. В железе гусеницы шёлк является насыщенным водой и образует коллоидную суспензию. Её превращение в нити с ориентированными молекулами происходит путём натяжения в направлении продольной оси и давления, направленного перпендикулярно линии натяжения[74].
Образ жизни


_caterpillars.jpg.webp)
Преобладающее большинство гусениц ведёт наземный образ жизни[76]. Помимо этого существует небольшое количество видов с водными гусеницами[77][78]. К ним относятся некоторые представители семейства настоящих огнёвок. Гусеницы ряда видов подсемейств Pyraustidnae, Acentropinae и Nymphulinae семейства травяных огнёвок также обитают под водой[77], например белая водная огнёвка, кувшинковая водная огнёвка[79]. В тропических регионах наиболее многочисленны представители подсемейства Nymphulinae. Гусеницы, живущие в воде, имеются и в группе медведиц (род Palastra из Южной Америке)[77]. Некоторые гусеницы живут внутри водных и болотных растений[77]. Несколько видов гусениц гавайских роскошных молей рода Hyposmocoma являются земноводными и способны жить как под водой, так и на суше[80]. Приспособление органов дыхания к водяной жизни у гусениц преимущественно чисто внешнее. Строение дыхалец аналогично таковому у сухопутных видов. Гусеницы получает нужный для дыхания воздух время от времени всплывая на поверхность и унося его с собой в виде пузырьков на своём волосяном покрове, похожим на густой войлок. Другое приспособление имеется у гусениц рода Nymphula. В ранних возрастах они минируют листья водяных растений, а во взрослом состоянии сооружают «мешок» из двух кусочков листа. В младшем возрасте они лишены особых органов дыхания и дышат через кожу. По мере взросления у них открываются дыхальца, и дыхание осуществляется посредством трахей. Таким же образом дышат гусеницы родов Cataclysta и Acentropus[77][81].
Время суточной активности гусениц зависит от вида и стадии развития. Например, гусеницы представителей рода аполлонов (Parnassius) ведут исключительно дневной образ жизни и активны только в солнечную погоду[82], а гусеницы совок (Noctuidae) активны преимущественно ночью. Гусеницы непарного шелкопряда на первых возрастах активны преимущественно днём, а на поздних — в ночное время суток[83].
По образу жизни гусеницы условно разделяются на две большие группы[76]:
- гусеницы, ведущие свободный образ жизни, открыто питающиеся на кормовых растениях;
- гусеницы, ведущие скрытый образ жизни.
Гусеницы дневных, или булавоусых, бабочек, а также большинства других крупных чешуекрылых живут открыто на кормовых растениях. Гусеницы многих семейств молевидных чешуекрылых ведут скрытный образ жизни: в почве, подстилке или дерновине злаков (часто в шелковинных ходах); внутри кормовых растений, минируя листья, побеги и плоды; изготавливая разнообразные чехлики, которые гусеница, ползая, перетаскивает за собой (наиболее известны этим мешочницы, но ношение чехликов распространено гораздо шире)[76].
Скрытный образ жизни характерен для значительной части гусениц, принадлежащих к разным систематическим группам. Среди них выделяют несколько биологических типов, наиболее важнейшими среди которых являются следующие[76]:
- листоверты — живущие в сплетённых или свёрнутых листьях; к ним относятся не только представители семейства листоверток;
- карпофаги — гусеницы, которые живут в плодах;
- бурильщики (ксилофаги) — живущие внутри одеревеневших частей растения: в побегах, ветвях, стволах, а также в корнях и почках;
- минёры — живущие внутри тканей листьев, черешков, а также в коре побегов и оболочке плодов;
- галлообразователи — обладающие способностью вызывать разрастание тканей на повреждённых ими частях растений;
- подземные гусеницы — обитают преимущественно в почвенном слое;
- гусеницы, ведущие водный образ жизни.
Галлообразующие гусеницы малочисленные. Из них можно выделить группы таких, которые образуют галлы лишь при случае, и собственно гусениц, которые живут только в галлах и специально к ним приспособлены. Среди обитателей чужих галлов есть гусеницы, которые в них развиваются или только окукливаются. Например, Chionodes electella минируют еловую хвою, а затем живут в пустых галлах хермесов; Gelechia basipunctella питается почками и листьями ивы, а затем живёт в галлах ильмовой златки. Многие виды листовёрток рода Pammene живут на дубах в галлах других насекомых. К истинным галлообразователям относится, например, Incurvaria tenuicornis, живущая во вздутиях ветвей берёз, Argyresthia semifusca — в почках и вздутиях молодых ветвей черёмухи, а многие виды узкокрылых молей рода Mompha являются галлообоазователями на кипреи. Также большое число видов-галлообразователей известно среди семейства листовёрток[84].
В почве развивается ограниченное число видов гусениц из семейств тонкопряды, некоторые листовёртки, настоящие огнёвки (однако большинство из них проводит большую часть своей жизни в корнях), а также группа подгрызающих совок. Для обитающих в почве гусениц характерна широкая полифагия (многоядность). Подземный образ жизни гусениц является формой специализацией и обеспечивает лучшую защиту от паразитов и хищников[76].
Ряд гусениц образуют т. н. паутинные гнёзда, которые представляют собой скопления шелковистой нити, покрывающие развилки ветвей, целые ветви или в крайних случаях даже отдельные деревья целиком. В подобных гнёздах в течение лета питаются гусеницы горностаевых молей, американской белой бабочки, кольчатого шелкопряда. Небольшие паутинные гнёзда, называемые зимними, в виде нескольких сухих листьев, скреплённых шелковиной, служат для зимовки гусениц боярышницы и златогузки на ветвях деревьев[85].
Экология
Миграции

Миграционное поведение у гусениц отмечаются гораздо реже, чем у бабочек. Оно возникает при очень высокой численности, недостатке корма и ряде прочих факторов. Гусеницы некоторых видов, например капустницы, в поисках пищи способны на далёкие миграции, заселяя соседние участки с пригодной для питания растительностью. Миграции гусениц младшего возраста способствуют выживанию определённой части популяции даже в условиях негативного воздействия абиотических и биотических факторов. Наиболее известными, склонными к миграциям, являются гусеницы походных шелкопрядов (Thaumetopoeidae). Они обладают инстинктом миграций к новым местам кормёжки, собираясь обществами в виде длинных колонн или процессий, отчего и получили своё название. Во время таких миграций гусеницы походных шелкопрядов передвигаются цепочкой друг за другом, следуя за впереди передвигающейся гусеницей. Также они руководствуются шелковистой нитью, которую оставляет «гусеница-вожак», которая движется медленно и совершает ощупывающие, поисковые движения в различных направлениях[86].
Один из самых знаменитых экспериментов известного французского энтомолога Жана Анри Фабра, проделанный им в 1896 году, был посвящён именно данным особенностям поведения гусениц соснового походного шелкопряда. Направляемая колонна этих гусениц вползла на верхний край большого цветочного горшка. «Гусеница-вожак», достигнув его края, начала ползти по нему, описывая окружность; остальные гусеницы инстинктивно продолжали двигаться за ней. Когда «гусеница-вожак» проползла полный круг и наткнулась на очередную гусеницу, выползающую на край кадки, Фабр удалил всех остальных гусениц с боковой стенки горшка и щёточкой тщательно стёр нить, оставленную на ней гусеницами. После чего «гусеница-вожак», обнаружив нить, стала следовать за последней гусеницей, перестав быть «вожаком». Колонна гусениц «замкнула круг» и начала передвигаться по краю горшка. Примечательно, что гусеницы продолжали двигаться «по кругу» восемь дней подряд, распадаясь на две группы в период «отдыха» в ночное время и воссоединяясь вновь в «кольцо» с началом периода своей суточной активности, пока из-за истощения и голода не стали по очереди падать с края горшка[87].
Защита от хищников




Многие животные питаются гусеницами, поскольку они богаты белками. В результате, гусеницы развили различные средства защиты от хищников и способны разнообразно защищаться от природных врагов[88]. Многие гусеницы ведут ночной образ жизни, который действенно предохраняет их от возможных нападений птиц и других дневных хищников[88].
Окраска и форма тела гусениц как стратегия защиты обычно разделяется на три категории: предостерегающая окраска, покровительственная окраска и мимикрия несъедобным объектам или сходство с организмами опасным для хищников[88]. Открыто живущие гусеницы преимущественно имеют покровительственную окраску, что делает их малозаметными на кормовых растениях[88]. Либо же они, наоборот, отличаются яркой окраской, так как несъедобны для хищников, а заметная, запоминающаяся внешность служит для них защитой от хищников. Птица, которая попробовала съесть обладающую неприятным вкусом гусеницу, запоминает этот случай и больше не трогает гусениц с подобной окраской тела. Примеров покровительственной окраски среди гусениц имеется огромное множество: многие гусеницы, держащиеся среди листвы и трав окрашены в зелёный цвет; обитающие на коре деревьев — неотличимы от неё по окраске (например гусеницы орденских лент); живущие на соцветиях гусеницы также прекрасно скрываются благодаря своей окраске, сопоставимой с цветками. Интересно, что если один и тот же вид гусеницы обитает на цветах различной окраски, так порой их окраска изменяется. Особенно хорошо это явление отмечается у некоторых видов пядениц из рода Tephroclystia. Значительная изменчивость окраски, в зависимости от кормового растения, характерна для целого ряда гусениц[31].
Гусеницы многих пядениц характеризуются тонким палочковидным телом, голыми покровами, покрытыми бугорковидными возвышениями, и имеют сходство с сухими веточками. При этом у этих гусениц имеются особенные охранительные инстинкты, благодаря которым данное сходство ещё больше увеличивается — они способны часами находиться в неподвижном состоянии, прикрепившись последними парами конечностей к ветке, а остальной частью тела откинувшись назад, благодаря чему достигается полное сходство гусеницы с веточкой растения[31]. Гусеницы нимфалид из рода Adelpha на поздних возрастах выглядят как кусок мха[88]. Гусеницы многих пядениц обладают окраской под цвет коры, веточек, стеблей и т. п. Благодаря развитым мышцам они способны, закрепившись задними ногами на веточке, приподняв и вытянув тело, оставаться неподвижными длительное время, становясь похожими на обломанные веточки, сучки, черешки листьев и т. п. Для гусениц ряда видов как средство защиты характерен танатоз — временная неподвижность, при которой они притворяются мёртвыми. При вероятной опасности такие гусеницы обычно замирают и падают с растений на шелковинной нити[89].
.jpg.webp)
Волосяной покров служит для многих гусениц отличной защитой от большинства птиц, так как очень немногие из них поедают мохнатых гусениц, большинство же не трогает их[90]. Благодаря этой особенности, многие мохнатые и шиповатые гусеницы часто держатся открыто, иногда группами и имеют яркую предостерегающую или предупреждающую окраску[31]. У некоторых видов гусениц полые внутри хитиновые волоски сообщаются с кожными железами, ядовитый секрет которых вызывает у хищников сильное раздражение (например, шипики у гусениц павлиноглазки сатурнии ио). Помимо этого длинные волоски гусениц также могут выступать в качестве своеобразных «органов чувств», сообщающих об приближении хищника. Например, волосатые гусеницы эребиды Orgyia leucostigma в ответ на высокую скорость изгибания волосков падают с растения-хозяина, а при низкой скорости — уползают[88].
У гусениц бабочек из семейства парусников и хохлаток имеется особая вилкообразная железа — осметрий — имеющий вид двух длинных оранжево-красных «рожек»[91][92]. Гусеница выдвигает осметрий из своего тела при раздражении и возможной опасности. При этом из железы выделяется особый секрет оранжево-жёлтого цвета с неприятным резким и едким запахом, содержащий терпены. Подобным образом защищаются только гусеницы первых и средних возрастов, а гусеницы последних возрастов при опасности железу не выдвигают[93].

Гусеница большой гарпии при опасности принимает специальную угрожающую позу: надувает и поднимает передний конец тела, затем втягивая его в увеличенный первый сегмент брюшка, при этом на заднем конце тела вытягиваются две пахучие нити вилкообразного выроста. Сильно потревоженная гусеница выделяет капельки едкой гемолимфы из «вилки» на конце тела и может выбрызгивать её на расстояние до 10 см из поперечной щели, расположенной на первом грудном сегменте[23].
Гусеницы бражников вида Amorpha juglandis для отпугивания хищников в случае нападения издают свист, выпуская воздух через пару дыхалец на восьмом сегменте брюшка. Эти звуковые сигналы варьируются — начиная от звуков, различимых человеком, и до ультразвука. Они могут состоять из 1—8 звуков, различных по своему спектру: от простого монотонного сигнала до сложной звуковой композиции с пиками на частотах 9, 15 и 22 кГц[94][95][96].
Гусеницы некоторых представителей рода парусников-хвостоносцев, а также видов стрельчатка ольховая, Macrauzata maxima, Apochima juglansiaria и других имитируют своей окраской птичий помёт[97]. Многие гусеницы, не обладая необходимой окраской, всё равно принимают различные специфические позы, полагаясь на то, что птицы сочтут их несъедобными[97]. Гусеницы некоторых видов парусников-хвостоносцев (бианор, P. troilus и многие другие), бражника Hemeroplanes triptolemus и эребеид рода Phyllodes своей окраской имитирует змею[98]. При этом гусеницы Hemeroplanes triptolemus при опасности одновременно расширяют сегменты брюшка и груди, образуя форму головы змеи, что усиливает сходство[88].
Некоторые виды обладают скверным запахом и неприятным вкусом либо ядовиты, что делает их несъедобными. Попробовав такое насекомое один раз, хищники будут избегать подобный вид в дальнейшем. Химические средства защиты у гусениц представлены продукцией химических соединений и получением токсичных соединений из поедаемых растений-хозяев, которые служат защитой от естественных врагов[88].
Паразиты и паразитоиды



Среди настоящих паразитов следует отметить отдельные виды грибов, например, Cordyceps sinensis и другие представители рода Cordyceps, а также одноклеточных, например, микроспоридии родов Thelohania и Plistophora[99][100].
Существует большое число организмов, развивающихся за счёт гусениц, но не являющихся истинными паразитами, так как в результате их жизнедеятельности организм-хозяин в любом случае либо гибнет, либо теряет способность к продолжению рода. Принятое в науке название представителей группы, обладающей описанной формой биологических взаимоотношений, — паразитоиды[101]. Гусеницы подвержены нападению личиночных, личиночно-куколочных паразитоидов. Многие представители браконид, одного из семейств наездников, паразитируют на гусеницах[102]. Как и большинство других наездников, бракониды в основном откладывают яйца в свободно передвигающихся гусениц, некоторые — предварительно парализуют жертву, частично либо полностью. Последнее характерно для видов, специализирующихся на скрытоживущих хозяевах, вынужденных отыскивать и доставать при помощи своего длинного яйцеклада гусениц, например, под корой деревьев. Длительный или необратимый паралич наступает, как правило, от вводимого наездником в тело хозяина избыточного количества токсина. Из браконид наибольшее значение в России имеют Apanteles glomeratus — главный истребитель белянок, Habrobracon juglandis — паразит мучной огнёвки, Apanteles solitarius и Meteorus versicolor — паразиты ивовой волнянки. Бракониды рода Microgaster заражают гусениц капустницы, которые после перехода в стадию куколки поражаются представителями другого семейства — птеромалид (Pteromalidae) — Pteromalus puparum[103].
Самки наездников-ихневмонид из отряда перепончатокрылых, принадлежащие исключительно к личиночным и яйцеличиночным паразитам, откладывают свои яйца под кутикулу гусениц или непосредственно на неё. В последнем случае вылупившиеся личинки сами внедряются внутрь хозяина. Обычно молодые личинки питаются гемолимфой, а в конечных стадиях своего развития переходят на питание тканями и внутренними органами гусеницы. Окукливание происходит в теле гусеницы либо вне его[104][105].
Роющие осы рода аммофилы охотятся на гусениц, которых парализуют, после чего переносят в гнездо, где ими будут кормится личинки[106].
На гусеницах также паразитируют двукрылые, прежде всего тахины, или ежемухи (Tachinidae). Самки откладывают яйца или личинок, в случае живорождения, непосредственно на гусеницу. Другие виды ежемухи могут откладывать яйца на листву кормового растения гусениц. У гусениц, съевших их вместе с листвой, выход личинок происходит в пищеварительном тракте хозяина, откуда в дальнейшем они внедрятся в полость тела. Окукливание происходит внутри хризалиды, либо, в случае гибели гусеницы — в почве[107].
Симбионты

.jpg.webp)
Гусеницы примерно половины всех видов голубянок (Lycaenidae) так или иначе связаны в цикле своего развития с муравьями. Они живут в муравейниках, находясь с муравьями в симбиотических отношениях, например, с родом мирмик (Myrmica). Взаимосвязи между муравьями и гусеницами голубянок по своему типу могут быть мутуалистическими или паразитическими. Такие ассоциации варьируются от слабых факультативных взаимодействий, в которых личинки лишь изредка посещаются несколькими видами муравьёв (около 45 % ассоциаций), до сложных облигатных ассоциаций, в которых гусеницы всегда находятся под присмотром муравьёв, часто только одного единственного вида (30 %). 12 % облигатных ассоциаций с муравьями (3 % всех ассоциаций) являются паразитическими или иногда, даже, хищническими: гусеницы голубянок, принятые в муравейник, в некоторых случаях выступают в роли хищников, поедающих личинок муравьёв[108].
Облигатные ассоциации встречаются гораздо чаще в южном полушарии, чем в северном. В Австралии и ЮАР более распространены облигатные ассоциации (39 % и 59 %, соответственно), в то время как в Неарктике только 2 % голубянок считаются облигатными мирмекофилами, а более 80 % исследованных видов остаются без присмотра муравьёв. В Юго-Восточной Азии облигатные и факультативные ассоциации муравьёв и гусениц голубянок составляют 22 % и 53 %, соответственно, а в Палеарктике — 12 % и 67 %; в остальных случаях таковые связи в этих зоогеографических областях не выявлены (25 % и 20 %)[109].
Гусеницы мирмекофильных голубянок имеют целый ряд органов для облегчения общения с муравьями и манипулирования ими. Например, такие органы как дорзальная нектароносная железа (dorsal nectary organs, DNOs), куполовидные органы (pore cupola organs, PCOs), щупальцевые органы (tentacle organs, TOs), — обнаруживаются у большинства личинок голубянок, связанных с муравьями. Они действуют, чтобы умиротворять муравьёв или привлечь их внимание, выделяя летучие вещества, когда личинки ими потревожены. Нектароносная железа на спинной поверхности 7-го сегмента продуцирует сладкие выделения и имеет первостепенное значение в мирмекофильной ассоциации для «вознаграждения» привлечённых муравьёв. Эти органы являются экзокринными железами, которые производят «медвяную росу», сильно отличающуюся от сходных выделений равнокрылых насекомых. Функционально эти органы находятся ближе к внецветковым нектарникам растений, чем к экскреторной системе тлей. Вторичная потеря этих органов часто приводит к исчезновению мирмекофильных ассоциаций, что указывает на то, что выделение нектара имеет большое значение[109].
Гусеницы многих голубянок и риодинид обладают комплексом эволюционно выработанных химических и акустических сигналов для контролирования поведения муравьёв. Одним из примеров мирмекофилии являются взаимовыгодные отношения муравьёв и гусениц, проявляющееся в трофобиозе. Гусеницы выделяют из дорзальной нектароносной железы сладкую жидкость, привлекающую муравьёв. Муравьи, ощупывая усиками этот орган, вызывают рефлекторное выделение гусеницей жидкости, которая, вероятно, содержит феромоны лихневмоны, обусловливающие поведение муравьёв. В ответ муравьи обеспечивают защиту гусениц от потенциальных хищников и врагов. Известны формы трофобиоза муравьёв с гусеницами некоторых видов бабочек из семейств голубянки и риодиниды[110]. Некоторые гусеницы голубянок и риодинид обладают также звукоиздающими органами, влияющими на поведение муравьёв — одни виды имеют только шаровую щетинку на теле гусеницы, другие — вместе с звуковой ресничкой, при отсутствии нектароносной железы. Таким образом они производят вибрации и звук, воспринимаемые муравьями[111].
Все виды мирмекофильных голубянок приурочены к луговым муравейникам. Одним из таких видов является голубянка алькон (Maculinea alcon), самки которой откладывают яйца на цветы растений семейства горечавковые. Большинство видов голубянок развиваются в гнёздах лишь одного вида муравьёв, но гусеницы голубянки алькон и голубянки арион обитают в гнёздах муравьёв различных видов, на разных участках своего ареала[112][113]. Гусеницы огневок рода Niphopyralis (Wurthia) живут в гнёздах древесных муравьёв Oecophylla и полирахис[114]. Гораздо менее многочисленными являются случаи термитофилии — совместного обитания с термитами в их постройках. Среди примеров: 1 вид настоящих молей из Бразилии, 1 вид огнёвок на Шри-Ланке[44].
К симбиозу можно отнести гусениц огнёвки Zophodiopsis hyanella, обитающих в паутинных гнездах африканских походных шелкопрядов Anaphe, где они питаются, подобно видам экскрементами и другими отбросами гусениц шелкопрядов[115]. Также известны интересные случаи симбиоза гусениц с пауками. В Южной Африке в общественных паутинных гнёздах пауков Stegodyphus обитают гусеницы Coccidiphila stegodyphobius из семейства роскошные моли — в хорошую погоду они расползаются по паутине и питаются остатками добычи пауков, а в плохую прячутся в гнёздах совместно с пауками, которые также никогда не нападают на бабочек этого вида, постоянно находящихся здесь же. В Индии подобным же образом с пауками Stegodyphus живут гусеницы Philarachnis xerophaga[44].
На Шри-Ланке известен случай симбиоза гусениц Nepenthophilus tigrinus с насекомоядным растением Nepenthes distillatoria в жидкости «кувшинов» которого они обитают и питаются попавшими в него насекомыми, строят себе чехлик из их остатков[116][117].
Гусеницы-минёры Phyllonorycter blancardella из семейства молей-пестрянок (Gracillariidae) живут в симбиозе с бактериями, выделяющими цитокины, которые стимулируют деление клеток растения, продлевая фотосинтез. Образующиеся в результате этого «зелёные островки» позволяют гусенице пережить зиму[118].
Симбионты (бактерии и грибы) найдены в кишечнике гусениц древоточцев рода Cossus — они разлагают клетчатку, выделяя из неё целлобиозу, глюкозу и глюкуроновую кислоту, которые затем усваиваются гусеницей. Бактерии также могут вызывать дальнейший распад данных веществ с образованием летучих жирных кислот и молочной кислоты[119].
Роль в экосистемах
Роль гусениц в естественных экосистемах заключается в том, что, будучи консументами первого порядка, они являются неотъемлемыми участниками пищевых цепей. Будучи преимущественно растительноядными гусеницы потребляют большую часть растительного прироста. В свою очередь, они сами служат кормом многим насекомоядным беспозвоночных и позвоночных животных. Очень велика их роль в питании ряда насекомоядных птиц, которые не только поедают их сами, но и собирают в больших количествах в качестве корма для своих птенцов[120][121].
Гусеницы в культуре

В Ветхом Завете Библии гусениц опасаются как вредителей, которые пожирают урожай. Они являются частью «саранчи»[122][123][124]: псалом 104 описывает как Господь использовал гусениц и саранчу в одной из казней египетских[125]. В Европе в Средневековье гусеницы стали символом социальных иждивенцев[126]. Шекспировский «Болингброк» описывает друзей короля Ричарда II как «гусениц Содружества». В 1790 году английский поэт Уильям Блейк (1757—1827) сослался на этот популярный образ в «Бракосочетании Рая и Ада» («The Marriage of Heaven and Hell») в своих нападках на священников: «Поскольку гусеница выбирает самые прекрасные листья, чтобы отложить яйца, поэтому священник проклинает самые прекраснейшие радости»[126].
Роль гусениц, как этапа жизненного цикла бабочек, на протяжении веков была плохо понята. В 1679 году немецкая художница и энтомолог Мария Сибилла Мериан (1647—1717) опубликовала первый том книги с акварельными иллюстрациями собственного авторства, посвящённой гусеницам. Она содержала 50 иллюстраций и описаний насекомых, включая бабочек и их гусениц[127]. Изданное до этого популярное издание книги о бабочках и их гусеницах голландского натуралиста и художника Яна Гёдарта (1617—1668) не включало яйца на этапах жизненного цикла европейских видов бабочек, поскольку он полагал, что гусеницы возникают из воды. Когда Мериан опубликовала своё исследование гусениц, все ещё широко было распространено мнение, что насекомые зарождались самопроизвольно. Вторая книга Мериан была издана в 1683 году, а третий том в 1717 году завершила и опубликовала младшая дочь художницы — Доротея Мария[127].
Наибольшее значение для символики бабочки имеет цикл её развития. Превращения гусеницы в куколку, а потом в бабочку является символом возрождения, воскрешения и перерождения[128]. Эти же превращения представлялись в культурах многих народов символом трансформации, воскресения и бессмертия благодаря жизненному циклу: жизнь (гусеница) — смерть (куколка) — возрождение (бабочка). Метаморфозы гусеницы в бабочку в западных культурах ассоциировались с человеческими трансформациями в фольклоре и литературе[128]. Бабочки считались символом человеческой души с древних времён, а также в христианской традиции[129]. К XVIII веку в европейских культурах жизненный цикл бабочек ассоциировался как аллегория христианского вероучения о спасении души. При этом ненасытные гусеницы ассоциировались с людьми, обречёнными на земной труд в грязи и страданиях; кокон выступал своего рода гробом, в который природа заключала свои создания в конце их земного пути, а вылупившаяся из куколки бабочка получала «жизнь после смерти»[130]. Таким образом, гусеница в христианской традиции стала метафорой «перерождения»[130].
Ян Гёдарт утверждал, что метаморфоза из гусеницы в бабочку был символом и даже доказательством воскресения Христа[128]. Голландский натуралист и анатом Ян Сваммердам (1637—1680), продемонстрировавший в 1669 году, что внутри окукливающейся гусеницы имеются зачатки конечностей и крыльев будущей бабочки, указывал на мистическое и религиозное представление о том, что «смерть» гусеницы сменяется впоследствии «воскресением» бабочки[128].
В книге «Приключения Алисы в Стране чудес» (1865 год) английского математика и писателя Льюиса Кэрролла есть персонаж Гусеница (англ. Caterpillar), которая курит кальян[131]. Современные отсылки на гусениц имеются в популярной литературе и кино. «Очень голодная гусеница» — детская книжка с картинками американского писателя Эрика Карла. Вышла в 1969 году и впоследствии многократно переиздавалась. Текст книги представляет собой историю гусеницы перед её превращением в бабочку. Книга издаётся на картонных листах, яркая особенность — наличие дырок в картоне («проеденных» гусеницей)[132][133]. Гусеницы с антропоморфными чертами являются персонажами многих анимационных фильмов: «Путешествие муравья» (1983)[134], «Гагарин» (1994), «Приключения Флика» (1998), «Minuscule» (2006), «Приключений Лунтика и его друзей» (с 2006) и других.
Взаимодействие с человеком
Хозяйственное значение

Очень велико хозяйственное значение гусениц ряда видов как вредителей сельского и лесного хозяйства, повреждающих полевые, садовые или лесные культуры. Большую группу образуют виды, наносящие вред различным лесным и сельскохозяйственным растениям[135]. Так, вредителями хвойных лесов считаются сибирский и непарный шелкопряды. В периоды их массового размножения, когда численность гусениц может достигать тысячи особей на одно дерево, они уничтожают хвою, после чего деревья часто погибают[136]. Листья плодовых культур объедают гусеницы различных видов чешуекрылых: боярышницы, непарного и кольчатого шелкопряда, зимней пяденицы, яблонной горностаевой моли, златогузки и многих других видов[135]. Американская белая бабочка повреждает многие виды плодовых и ягодных кустарников[135]. Гусеницы Papilio demoleus является вредителем многих культурных видов цитрусовых в Южной Азии, Индии и на Ближнем Востоке[137]. Плоды повреждают гусеницы бабочек семейств Листовёртки (в том числе плодожорки — яблонная, грушевая, сливовая и пр.) и Огнёвки (крыжовниковая огнёвка и другие). Полевым культурам значительный ущерб наносят гусеницы многих совок и огнёвок, которым свойственно массовое размножение (озимая совка и луговой мотылёк)[138][139]. Вред овощным культурам могут наносить гусеницы бабочек, поедающие листья (капустная моль, капустница, многие совки), а также бутоны, цветки, семена (зонтичная моль и ряд видов огнёвок питаются на семенниках моркови, укропа, сельдерея и др.). На виноградниках могут вредить гусеницы листовёртки виноградной, двулётной[140]. Вредителями запасов являются гусеницы огнёвки зерновой, мучной, мельничной, моли амбарной и зерновой, и других[141]. Мехам, одежде из натуральных тканей и мебели вредят шубная моль, платяная моль и мебельная моль. Пчеловодству вредят малая пчелиная огнёвка и большая восковая моль[142].
В то же время гусеницы некоторых видов могут использоваться в борьбе с сорными растениями. Например, в борьбе с амброзией используют амброзиевую совку (Tarachidia candefacta), гусеница которой питается только листьями амброзии полынолистной, не повреждая других растений[143]. В США гусениц некоторых видов пальцекрылок используют для борьбы с сорными растениями вроде вербены кустарниковой. Самым же ярким примером является специально завезённая в 1925 году в Австралию из Уругвая и из северных районов Аргентины кактусовая огнёвка (Cactoblastis cactorum), которая помогла избавиться от завезённого кактуса опунции, которым заросли миллионы гектаров пастбищ[144]. В 1938 году в долине реки Дарлинг австралийские фермеры поставили специальный памятник гусеницам, «спасшим» Австралию[145].
Производство шёлка

Шелководство является отраслью сельского хозяйства, целью которой является производство сырья для выработки натурального шёлка. Оно представляет собой специальное разведение гусениц некоторых видов для получения шёлка. Текстильная промышленность отдаёт предпочтение тутовому шелкопряду (Bombyx mori), одомашненному человеком. Согласно конфуцианским текстам, производство шёлка с использованием тутового шелкопряда началось около XXVII века до н. э., хотя археологические исследования позволяют говорить ещё о периоде Яншао (5000 лет до н. э.)[146]. В первой половине I века н. э. шелководство пришло в древний Хотан[147] и в конце III века пришло в Индию. В Европе, включая Средиземноморье, шелководство возникло в VIII веке, а более трёхсот лет тому назад оно появляется и в Россию. Впоследствии основными центрами шелководства становятся республики Средней Азии и Закавказья. Тутовый шелкопряд играет важную экономическую роль в производстве шёлка. В результате селекции выведено много пород тутового шелкопряда, отличающихся продуктивностью, качеством шёлковой нити и цветом коконов. Для получения шёлковой нити куколок предварительно умерщвляют с помощью горячего пара и воды на десятый день после окукливания. В шёлковом коконе обычно содержится до 3500 метров волокна, но размотать его удаётся едва на треть[148]. Чтобы получить 1 килограмм шёлка-сырца, нужны коконы примерно тысячи гусениц, поедающих за полтора месяца 60 килограмм листьев тутового дерева. Из 100 кг коконов можно получить примерно 9 кг шёлковой нити[148]. В настоящее время во всём мире ежегодно производится 45 тысяч тонн шёлка. Главными поставщиками являются Китай, Индия, Япония, Корея и Узбекистан[149][150][151].
В шелководстве также используются такие виды, как китайская дубовая павлиноглазка (Antheraea pernyi), которую с XVII—XVIII века разводят в Китае (из её коконов получают шёлк, который идёт на изготовление чесучи)[152], а также японская дубовая павлиноглазка (Antheraea yamamai). Последняя выращивается в Японии[153] уже более 1000 лет. Её кокон легко разматывается, а шёлковая нить, полученная из него, является мягкой, толстой, блестящая. По прочности равна нити тутового шелкопряда, а по эластичности превосходит её. Павлиноглазка Samia cynthia даёт т. н. шёлк «ри», из которого изготавливают тонкотканные платки — шантунги[148].
В Индии также разводится павлиноглазка атлас, гусеницы которой выделяют шёлк отличающийся от такового у тутового шелкопряда — он обладает коричневым цветом, более крепкий и шерстистый — и носит название фагаровый шёлк (англ. fagara silk)[154].
Многие из других видов «шелкопрядов» плохо разводятся в неволе, поэтому обычно ограничиваются лишь сбором их коконов в дикой природе[148].
Использование в пищу

Энтомофагия (поедание насекомых) является широко распространённым явлением по всему миру[155]. Гусеницы представителей примерно восьмидесяти родов из двадцати семейств чешуекрылых употребляются в пищу как богатые белком пищевые продукты. Их едят в сыром или жареном виде[155].
Гусеницы употреблялись человеком в пищу с доисторических времён. Например, индейцы Северной Америки в Каскадных горах и горах Сьерра-Невады собирали гусениц сатурнии пандоры (Coloradia pandora), которых подсушивали на слое горячих углей или отваривали[155]. Австралийские аборигены с давних времён употребляют в пищу гусениц древоточца Xyleutes leucomochla, которых добывают, выкапывая корни кустов акации и разрубая их, чтобы добраться до находящихся в них насекомых[155][156].
В Китае гусениц тутового шелкопряда засаливают, добавляют в омлеты из куриных яиц либо просто жарят с луком и густыми соусами. В Индии ежегодно потребляется более 20 тысяч тонн этих насекомых[157]. В Африке деликатесом считаются жареные или копчёные гусеницы павлиноглазки Gonimbrasia belina, которые стоят в 4 раза дороже обычного мяса[157]. В отдельных районах Северной Африки эти гусеницы настолько популярны, что в сезон их появления существенно падают продажи говядины и мяса других животных. В Ботсване их продают большими пакетами в сушёном виде как любые другие сухие продукты. Из них готовят особый суп. Традиционный метод сохранения гусениц состоит в том, чтобы высушить их на солнце или закоптить, в результате чего они приобретают дополнительный аромат. Промышленный метод — консервирование гусениц в рассоле. В Мозамбике и Зимбабве сельские женщины собирают гусениц Gonimbrasia belina ранней весной и тушат их с овощами либо сушат. Ежегодно в одной только Южной Африке продаётся около 1 600 тонн этих гусениц[155].
Гусеницы и здоровье человека

Гусеничный дерматит — энтомоз, обусловленный раздражением кожи волосками гусениц некоторых видов[158]. Представляет собой вид аллергического контактного дерматита. Заболевание возникает при проникновении в кожу щетинок и волосков гусеницы, содержащих токсические вещества, аналогичные по своему химическому составу кантаридину. Также развивается при воздействии веществ, выделяемых железистыми клетками покровов гусеницы. Характеризуется местными кожными и общими токсико-аллергическими реакциями. Сопровождается сильными субъективными ощущениями и развитием воспалительно-экссудативной реакции кожных покровов: на поражённых участках кожи возникают ярко-красные, зудящие высыпания, на периферии которых отмечаются зудящие папуло-везикулёзные элементы. При попадании волосков в конъюнктиву глаза развивается острый конъюнктивит[159][160]. Наибольшим раздражающим свойством обладают гусеницы Thaumetopoea processionea[161][162], Megalopyge opercularis[163][164][165].
Лономиаз — энтомоз, вызываемый отравлением токсином гусениц павлиноглазок из рода Lonomia в странах Южной Америки — преимущественно в Бразилии, Аргентине, Венесуэле и Колумбии. Сведения об отравлениях, обусловленные гусеницами Lonomia и сопровождающиеся выраженными геморрагическими расстройствами, в том числе с летальным исходом, впервые появились в 1960-е годы в Венесуэле. У пострадавших отмечаются похожие на гангрену пятна по всему телу, появляются внутренние кровотечения, гемолиз, могут возникать кровоизлияния в головном мозге, в желудочно-кишечном тракте, в почках, что приводит к развитию острой почечной недостаточности[166][159].
Сушёные гусеницы, заражённые грибком кордицепс китайский (Ophiocordyceps sinensis), применяются в китайской и тибетской народной медицине[167][168] для лечения широкого спектра заболеваний различных органов, а также в качестве гипогликемического и иммуностимулирующего средства и афродизиака[169].
Примечания
- Hall, Donald W. Featured Creatures: hickory horned devil, Citheronia regalis. University of Florida, Entomology and Nematology Department (September 2014).
- Фасмер М. Этимологический словарь русского языка. — Прогресс. — М., 1964–1973. — Т. 1. — С. 477.
- Boryś W. Słownik etymologiczny języka polskiego (польск.). — Wydawnictwo Literackie. — Kraków, 2005. — P. 158. — ISBN 978-83-08-04191-8.
- Гусеница // Словарь русского языка XI—XVII веков Вып. 4. — М.: Наука, 1977. — С. 160. — 16 400 экз.
- Полное собрание русских летописей. Том 1. Лаврентьевская летопись. Вып. 1. Повесть временных лет. — 2-е изд. — Л., 1926. — Стб. 168
- Пилильщики // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- Мамаев Б. М. Определитель насекомых по личинкам / ред. В. И. Сучинская. — М.: Просвещение, 1972. — С. 353. — 414 с.
- Герасимов, 1952, с. 68.
- Герасимов, 1952, с. 6—7.
- Герасимов, 1952, с. 13.
- Герасимов, 1952, с. 18.
- Герасимов, 1952, с. 20—26.
- Герасимов, 1952, с. 17—18.
- Жантиев Р. Д., Елизаров Ю. А., Мазохин-Поршняков Г. А., Чернышев В. Б. Руководство по физиологии органов чувств насекомых. — М.: Изд-во Московского Университета, 1977. — С. 136, 189. — 224 с.
- Герасимов, 1952, с. 31—34.
- Герасимов, 1952, с. 33—34.
- Герасимов, 1952, с. 34—35.
- Герасимов, 1952, с. 38—53.
- Герасимов, 1952, с. 53—66.
- Vidal Haddad Jr, Joel Carlos Lastória. Envenomation by caterpillars (erucism): proposal for simple pain relief treatment (англ.) // Journal of Venomous Animals and Toxins including Tropical Diseases : Журнал. — 2014. — Vol. 20. — P. 21. — doi:10.1186/1678-9199-20-21.
- Ламперт, 2003, с. 40—42.
- Ламперт, 2003, с. 39.
- Акимушкин И. И. Шестиногие членистоногие // Мир животных: Насекомые. Пауки. Домашние животные. — 4-е. — М.: Мысль, 1995. — Т. 3. — С. 13. — 462 с. — 15 000 экз. — ISBN 5-244-00806-4.
- Ламперт, 2003, с. 39—40.
- Герасимов, 1952, с. 70—71.
- Ламперт, 2003, с. 45.
- Герасимов, 1952, с. 120.
- Савчук В. В. Атлас бабочек и гусениц Крыма. — Симферополь: Бизнес—Информ, 2013. — С. 7. — 296 с. — ISBN 978-966-648-331-0.
- Вийдалепп Я. Р. К фауне чешуекрылых Тувинской АССР. IV. Пяденицы (Lepidoptera, Geometridae) // Учёные записки Тартуского университета. — 1979. — № 483 (12). — С. 79.
- Jaffe B. D., Guédot Ch., Ketterer M. et al. A caterpillar (Callopistria floridensis G. (Lepidoptera: Noctuidae)) accumulates arsenic from an arsenic-hyperaccumulating fern (Pteris vittata L.) (англ.) // Ecological Entomology. — 1998. — Vol. 44, no. 4. — P. 480–487. — ISSN 1365-2311.
- Лавров С. Д. Наши гусеницы. Определитель.. — М.: Государственное учебно-педагогическое издательство, 1938. — С. 3—21. — 144 с.
- Определитель насекомых Дальнего Востока России. Т. V. Ручейники и чешуекрылые. Ч. 2 / под общ. ред. П. А. Лера. — Владивосток: Дальнаука, 1999. — С. 28. — 671 с. — ISBN 5-7442-0910-7.
- Manley Ch. British Moths: Second Edition: A Photographic Guide to the Moths of Britain and Ireland (англ.). — Bloomsbury Publishing, 2015. — P. 92. — 352 p.
- Моуха Йозеф. Бабочки. — Прага: Артия, 1979. — С. 15—17. — 189 с.
- Горбунов О. Г. Биоразнообразие, зоогеография и история фауны бабочек-стеклянниц (Lepidoptera, Sesiidae) России и сопредельных территорий // Известия Академии Наук. Серия биологическая. — 1998. — Т. 4. — С. 453—457.
- Герасимов, 1952, с. 122.
- Tzanakakēs M. E. Insects and Mites Feeding on Olive. — Brill, 2006. — P. 121. — 182 p.
- Загуляев А. К. Моли — вредители меха, шерсти и борьба с ними. — М.—Л.: Издательство академии наук СССР, 1958. — С. 66—71. — 195 с.
- Загуляев А. К. Настоящие моли (Tineidae). Ч. 2. Подсемейство Nemapogoninae // Фауна СССР. Насекомые чешуекрылые. — М.—Л.: Издательство АН СССР, 1964. — Т. 4, вып. 2. — С. 232—243. — 424 с. — (Новая серия № 86).
- Герасимов, 1952, с. 152.
- Балашов Ю. С. Паразитизм клещей и насекомых на наземных позвоночных. — СПб.: Наука, 2009. — С. 124. — 357 с.
- Deyrup M., Deyrup N.D., Eisner M., Eisner T. A Caterpillar that Eats Tortoise Shells (англ.) // American Entomologist. — 2005. — Vol. 51, no. 4. — P. 245—248. Архивировано 16 декабря 2010 года.
- Grabe A. Eigenartige Geschmacksrichtungen bei Kleinschmetterlingsraupen («Strange tastes among micromoth caterpillars») (нем.) // Zeitschrift des Wiener Entomologen-Vereins. — 1942. — Nr. 27. — P. 105—109.
- Герасимов, 1952, с. 153.
- Gilmore D. P., Da Costa C. P., Duarte D. P. F. Sloth biology: an update on their physiological ecology, behavior and role as vectors of arthropods and arboviruses (англ.) // Brazilian Journal of Medical and Biological Research. — 2001. — January (vol. 34, no. 1). — P. 9—25. — ISSN 0100-879X. — doi:10.1590/S0100-879X2001000100002. — PMID 11151024.
- Ламперт, 2003, с. 59.
- Pierce, 2002, с. 748—751.
- Martin Hering. Biologie der Schmetterlinge. — Berlin: Springer-Verlag, 1926. — P. 73. — 482 p.
- Kaufman, Kenn; Brock, Jim P. «Coppers and Harvester». Kaufman Field Guide to Butterflies of North America. — Houghton Mifflin Books, 2003. — P. 80—81.
- Montgomery S. L. Carnivorous caterpillars: the behavior, biogeography and conservation of Eupithecia (Lepidoptera: Geometridae) in the Hawaiian Islands (англ.) // GeoJournal. — 1983. — Iss. 184, no. 7. — P. 549—556.
- Arnett, Ross H. Jr. American Insects. A Handbook of the Insects of America North of Mexico (англ.). — 2. — CRC press LLC, 2000. — P. 631—632. — ISBN 0-8493-0212-9.
- Rubinoff D., Haines W. P. Web-spinning caterpillar stalks snails (англ.) // Science. — 2005. — Vol. 309, no. 5734. — P. 575. — doi:10.1126/science.1110397. — PMID 16040699.
- Pierce N. E. Predatory and parasitic Lepidoptera: Carnivores living on plants (англ.) // Journal of the Lepidopterist's Society : Журнал. — 1995. — Vol. 49, no. 4. — P. 412—453 (417).
- Peigler R. S. Catalog of parasitoids of Saturniidae of the world (англ.) // Journal of Research on the Lepidoptera. — 1994. — Iss. 33. — P. 1–121.
- Брем А. Жизнь животных / Предисловие и комментарии М. С. Галиной и М. Б. Корниловой. — М.: Олма-Пресс, 2004. — С. 962. — 1192 с. — 5000 экз. — ISBN 5-224-04422-7.
- Yang J., Yang Y., Wu W. M., Zhao J., Jiang L.. Evidence of Polyethylene Biodegradation by Bacterial Strains from the Guts of Plastic-Eating Waxworms (англ.) // Environmental Science and Technology : Журнал. — 2014. — Vol. 48, no. 23. — P. 13776—13784. — doi:10.1021/es504038a. — PMID 25384056.
- Bombelli, Paolo; Howe, Christopher J.; Bertocchini, Federica. Polyethylene bio-degradation by caterpillars of the wax moth Galleria mellonella (англ.) // Current Biology : Журнал. — Cell Press, 2017. — 24 April (vol. 27, no. 8). — P. R292—R293. — ISSN 0960-9822. — doi:10.1016/j.cub.2017.02.060.
- Could These Tiny Plastic-Eating Caterpillars Hold The Answer To Our Trash Problem? - Hydration Anywhere. hydrationanywhere.com.
- Открыто движение гусеницы внутренностями вперёд. Мембрана (23 июля 2010). Дата обращения: 20 мая 2012.
- Тыщенко В. П. Основы физиологии насекомых. Том 1. — Л., 1976. — С. 181. — 363 с.
- Ламперт, 2003, с. 34—35.
- Morewood, W. Dean & Richard A. Ring. Revision of the life history of the High Arctic moth Gynaephora groenlandica (Wocke) (Lepidoptera: Lymantriidae) (англ.) // Canadian Journal of Zoology : Журнал. — 1998. — Vol. 76, no. 7. — P. 1371—1381.
- Черешнев И. А. (отв ред.), А.В. Андреев, Д. И. Берман, Н.Е. Докучаев, В.А. Кашин. Красная книга Чукотского автономного округа. Том 1. Животные. — Магадан: Дикий север, 2008. — С. 34—35. — 244 с.
- Сочивко А.В. Колыбель Аполлона // Химия и жизнь. — М., 2006. — Вып. 184, № 6. — ISSN 0130-5972. Архивировано 4 марта 2016 года.
- Герасимов, 1952, с. 110—111.
- Герасимов, 1952, с. 113.
- Герасимов, 1952, с. 116—118.
- Ламперт, 2003, с. 111—112.
- Герасимов, 1952, с. 119.
- Ламперт, 2003, с. 47.
- Ламперт, 2003, с. 47—51.
- Ламперт, 2003, с. 46—52.
- Герасимов, 1952, с. 26—29.
- Шовен Р. Физиология насекомых. — М.: Издательство иностранной литературы, 1953. — С. 149—152. — 494 с.
- Zaoming W., Codina R., Fernández-Caldas E., Lockey R. F. Partial characterization of the silk allergens in mulberry silk extract (англ.) // Journal of investigational allergology & clinical immunology. — 1996. — Iss. 6, no. 4. — P. 237—241. — ISSN 1018-9068.
- Ламперт, 2003, с. 120—125.
- Ламперт, 2003, с. 44.
- Ламперт, 2003, с. 151—152.
- Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 5. Высшие насекомые / Под общ. ред. С. Я. Цалолихина. — СПб.: Наука, 2001. — С. 75—76. — 825 с.
- Milius, Susan. Hawaiian Caterpillars Are First Known Amphibious Insects (англ.). U.S. News & World Report (23 марта 2010). Архивировано 11 февраля 2012 года.
- Герасимов, 1952, с. 151—152.
- Львовский А. Л., Моргун Д. В. Определители по флоре и фауне России. Выпуск 8 // Булавоусые чешуекрылые Восточной Европы. — М.: Товарищество научных изданий КМК, 2007. — С. 165—172. — 2000 экз. — ISBN 978-5-87317-362-4.
- Lance D. R., Elkinton J. S., Schwalbe C. P. Behaviour of late-instar gypsy moth larvae in high and low density populations (англ.) // Ecological Entomology : Журнал. — 1987. — Vol. 12, no. 3. — P. 267. — ISSN 1365-2311. — doi:10.1111/j.1365-2311.1987.tb01005.x.
- Герасимов, 1952, с. 150.
- Митюшев И. М. Лесная энтомология. — М.: Юрайт, 2019. — С. 67. — 176 с. — ISBN 978-5-534-11373-0.
- Кохманюк Ф.С. О миграциях гусениц и имаго непарного шелкопряда Ocneria dispar L. // Поведение насекомых как основа для разработки мер борьбы с вредителями сельского и лесного хозяйства. — Киев: Наукова думка, 1975. — С. 73—74.
- Жан Анри Фабр. Жизнь насекомых (Рассказы энтомолога) / сокр. пер. с фр. и обработка Н. Н. Плавильщикова. — М.: Учпедгиз, 1963. — С. 380. — 460 с.
- Greeney H. F., Dyer L. A., Smilanich A. M. Feeding by lepidopteran larvae is dangerous: a review of caterpillars’ chemical, physiological, morphological, and behavioral defenses against natural enemies (англ.) // Invertebrate Survival Journal. — 2012. — Vol. 9. — P. 7—34.
- Верещагин Б. В., Плугару С. Г. Насекомые. — Кишинев: Штиинца, 1983. — С. 204. — 376 с.
- Тэннер О. Способы защиты у животных / Пер. с англ. И. Гуровой; Под ред. Е. Н. Панова. — М.: Мир, 1985. — 129 с. — (Удивительный мир диких животных).
- Eisner T., Meinwald Y. C. The defensive secretions of a caterpillar (Papilio) (англ.) // Science : Журнал. — 1965. — Iss. 150. — P. 1733—1735.
- Crossley A. C., Waterhouse D. F. The ultrastructure of the osmeterium and the nature of its secretion (англ.) // PaTissue & Cell : Журнал. — 1969. — Vol. 1, no. 3. — P. 525—554. — doi:10.1016/S0040-8166(69)80020-0.
- Chow-Chin L.; Shing Chow Y. Fine structure of the larval osmeterium of Papilio demoleus libanius (Lepidoptera: Papilionidae) (англ.) // Annals of the Entomological Society of America. — 1991. — Vol. 84, no. 3. — P. 294—302. — ISSN 0013-8746.
- Fullard, James H.; Napoleone, Nadia. Diel flight periodicity and the evolution of auditory defences in the Macrolepidoptera (англ.) // Animal Behaviour : Журнал. — Elsevier, 2001. — Vol. 62, no. 2. — P. 349. — ISSN 0003-3472. — doi:10.1006/anbe.2001.1753.
- Bura V. L., Rohwer V. G., Martin P. R., Yack J. E. Whistling in caterpillars (Amorpha juglandis, Bombycoidea): Sound-producing mechanism and function (англ.) // The Journal of Experimental Biology : Журнал. — The Company of Biologists, 2010. — Vol. 214, no. Pt 1. — P. 30—37. — ISSN 0022-0949. — doi:10.1242/jeb.046805. — PMID 21147966.
- Knight K. Whistling Caterpillars Startle Birds (англ.) // The Journal of Experimental Biology : Журнал. — The Company of Biologists, 2010. — Vol. 214, no. Pt 14. — doi:10.1242/jeb.054155. — PMID 21834205.
- Кирилл Стасевич. «Туалетная маскировка»: как гусеницы притворяются птичьим помётом. Наука и жизнь (28 июня 2015).
- Bernard D'Abrera. Sphingidae Mundi. Hawk Moths of the World (англ.). — Classey Ltd., 1986. — P. 100—101. — ISBN 0860960226.
- Евлахова А. А. Энтомопатогенные грибы. Систематика, биология, практическое значение. — Л.:: Наука, 1974. — С. 136—137. — 254 с.
- Вейзер Я. Микробиологические методы борьбы с вредными насекомыми (Болезни насекомых). Монография.. — М.:: Колос, 1966. — С. 486. — 640 с.
- Askew R. R., Shaw M.R. Parasitoid communities: their size, structure and development // Insect Parasitoids. — London, 1986. — P. 225—264.
- Определитель насекомых Дальнего Востока России. Т. IV. Сетчатокрылообразные, скорпионницы, перепончатокрылые. Ч. 5 / под общ. ред. П. А. Лера. — Владивосток: Дальнаука, 2007. — С. 9. — 1052 с. — ISBN 978-5-8044-0789-7.
- Cai J., Ye G.Y., Hu C. Parasitism of Pieris rapae (Lepidoptera: Pieridae) by a pupal endoparasitoid, Pteromalus puparum (Hymenoptera: Pteromalidae): effects of parasitization and venom on host hemocytes // Journal of Insect Physiology. — 2004. — Вып. 50. — P. 315—322.
- Жизнь животных. Членистоногие: трилобиты, хелицеровые, трахейнодышащие. Онихофоры / Под ред. Гилярова М. С., Правдина Ф. Н. — 2-е изд., перераб. — М.: Просвещение, 1984. — Т. 3. — С. 348. — 463 с.
- Каспарян Д. Р. Сем. Ichneumonidae — ихневмониды // Определитель насекомых Дальнего Востока России. Т. IV. Сетчатокрылообразные, скорпионницы, перепончатокрылые. Ч. 5 / под общ. ред. П. А. Лера. — Владивосток: Дальнаука, 2007. — С. 255. — 1052 с. — ISBN 978-5-8044-0789-7.
- Фабр Жан Анри. Жизнь насекомых. Рассказы энтомолога. — М.: Государственное учебно-педагогическое издательство, 1963. — С. 70. — 466 с.
- Мамаев Б. М. Отряд Двукрылые, или Мухи и комары (Diptera) // Жизнь животных. Том 3. Членистоногие: трилобиты, хелицеровые, трахейнодышащие. Онихофоры / под ред. М. С. Гилярова, Ф. Н. Правдина, гл. ред. В. Е. Соколов. — 2-е изд. — М.: Просвещение, 1984. — С. 416. — 463 с.
- Pierce, 2002, с. 734.
- Stadler B., Dixon A. Mutualism. Ants and their Insect Partners (англ.). — Cambridge: Cambridge University Press, 2008. — P. 57—88. — 219 p. — ISBN 978-0-521-86035-2.
- Phillip J. Torres, Aaron F. Pomerantz. Butterfly Kleptoparasitism and First Account of Immature Stages, Myrmecophily, and Bamboo Host Plant of the Metalmark Adelotypa annulifera (Riodinidae) (англ.) // The Journal of the Lepidopterists' Society : Журнал. — 2016. — Vol. 70, no. 2. — P. 130—138. — doi:10.18473/lepi.70i2.a8.
- Pierce, 2002, с. 743—745.
- Pierce, 2002, с. 752.
- Fiedler K., Holldobler B., Seufert P. Butterflies and ants: «The communicative domain» (англ.) // Experientia : Журнал. — 1996. — Iss. 52, no. 1. — P. 14—24.
- Hölldobler B., Wilson E. O. The Ants. — Harvard University Press, 1990. — P. 483. — 732 p. — ISBN 0-674-04075-9.
- Hering M. Biologie der Schmetterlinge (нем.). — Springer-Verlag, 2013. — P. 411. — 482 p.
- Kitching R. L. Food Webs and Container Habitats: The Natural History and Ecology of Phytotelmata (англ.). — Cambridge University Press, 2000. — P. 369. — 431 p.
- Ellison A. M., Adamec L. Carnivorous Plants: Physiology, Ecology, and Evolution (англ.). — Cambridge University Press, 2018. — P. 320. — 510 p.
- Wilfried Kaiser, Elisabeth Huguet, Jérôme Casas et. al. Plant green-island phenotype induced by leaf-miners is mediated by bacterial symbionts (англ.) // Proceedings of the Royal Society of London B: Biological Sciences : Журнал. — 2010. — Vol. 277.
- Stokland J. N., Siitonen J., Jonsson B. G. Biodiversity in Dead Wood. — Cambridge: Cambridge University Press, 2012. — P. 40. — 509 p.
- Калуцкий А. Л. О бабочках // Биология. — 2001. — № 22. — С. 14—18.
- Прокофьева И. В. Мохнатые гусеницы в пище птенцов насекомоядных птиц // Русский орнитологический журнал. — 2020. — Т. 29, № 1878. — С. 328—332.
- Архимандрит Никифор. Иллюстрированная полная популярная библейская энциклопедия. — Проспект, 2018. — С. 168. — 1565 с.
- Амос. 4:9
- Иоил. 1:4
- Фи Г., Стюарт Д. Как читать Библию и видеть всю ее ценность. — СПб.: Шандал, 2009. — С. 114. — 416 с.
- Ferber M. Dictionary of Literary Symbols (англ.). — Cambridge: Cambridge University Press, 1999. — P. 104. — 263 p. — ISBN 9781316780978.
- Andréolle D. S., Molinari V. Women and Science, 17th Century to Present: Pioneers, Activists and Protagonists (англ.). — Cambridge: Cambridge Scholars Publishing, 2011. — P. 35—47. — 300 p.
- Enenkel K. A. E., Smith M. S. Early Modern Zoology: The Construction of Animals in Science, Literature and the Visual Arts (англ.). — BRILL, 2007. — P. 157, 161-162. — 718 p. — ISBN 9789047422365.
- Sax B. The Serpent and the Swan: The Animal Bride in Folklore and Literature (англ.). — University of Tennessee Press, 1998. — 70 p. — ISBN 9780939923687.
- Настюк Е. На крыльях и ползком: бабочки и жуки как символ в искусстве. АРТхив.
- Hephzibah Anderson. Alice in Wonderland’s hidden messages. BBC-Culture (31 мая 2016).
- ASHLEY FETTERS. How The Very Hungry Caterpillar Became a Classic. The Atlantic (20 марта 2019).
- Kate Taylor. Eat your heart out. The Guardian (2004).
- Асенин С.В. Пути советской мультипликации // Мир мультфильма: Идеи и образы мультипликации социалистических стран. — Москва: Искусство, 1986.
- Васильев В. П., Лифшиц И. З. Вредители плодовых культур. — М.: Государственное издательство сельскохозяйственной литературы, 1958. — С. 161—162. — 392 с.
- Плешанов А. С., Рожков А. С. Насекомые-дефолианты лиственничных лесов Восточной Сибири. — Наука, Сибирское отделение, 1982. — С. 149. — 208 с.
- Homziak N. T., Homziak J. Papilio demoleus (Lepidoptera: Papilionidae): A new record for the United States, Commonwealth of Puerto Rico (англ.) // Florida Entomologist. — 2006. — Iss. 89, no. 4. — P. 485—488.
- Вредители сельскохозяйственных культур и лесных насаждений. — Киев: Урожай, 1988. — Т. 2. Вредные членистоногие. — С. 311. — 392 с.
- Кузнецов В. И. Насекомые и клещи — вредители сельскохозяйственных культур. Том III. Чешуекрылые. Ч. 2. — Л.: Наука, 1999. — С. 177. — 410 с.
- Козарь И. М. Болезни и вредители винограда. Меры борьбы.. — Одесса: Национальный научный центр «Институт винорадарства и виноделия им. В. Е. Таирова», 2005. — С. 43. — 65 с.
- Загуляев А. К. Моли и огневки вредители зерна и продовольственных запасов. — М-Л.: Наука, 1965. — С. 59, 122, 132, 142. — 274 с.
- Тоскнна И.Н., Проворова И. Н. Насекомые в музеях: Биология. Профилактика заражения. Меры борьбы. — М.: Товарищество научных изданий КМК, 2007. — С. 205. — 242 с.
- Есипенко Л. П. Использование насекомых-фитофагов в борьбе с амброзией полыннолистной в агроценозах юга России // Земледелие. — 2013. — № 5. — С. 39—40.
- Blanka Pophof, Gert Stange, Leif Abrell. Volatile Organic Compounds as Signals in a Plant-Herbivore System: Electrophysiological Responses in Olfactory Sensilla of the Moth Cactoblastis cactorum (англ.) // Senses : Журнал. — 2005, April 14. — Vol. 30, no. 1. — P. 51—68.
- Floyd J. Pest Alert: Cactus moth Cactoblastis cactorum (англ.) (недоступная ссылка). APHIS (2006). Дата обращения: 18 февраля 2007. Архивировано 26 ноября 2012 года.
- Ebrey B. P., Walthall A. East Asia: A Cultural, Social and Political History (англ.). — Wadsworth Publishing, 2013. — P. 53. — 600 p. — ISBN 0-618-13387-9.
- Hill J. E. Annotated Translation of the Chapter on the Western Regions according to the Hou Hanshu (англ.). — 2nd Draft Edition. Appendix A. — 2003.
- Ален Эд, 2001, с. 177.
- Mulberry Silk - Textile Fibres - Handloom Textiles | Handwoven Fabrics | Natural Fabrics | Cotton clothes in Chennai (недоступная ссылка). Brasstacksmadras.com. Дата обращения: 29 января 2016. Архивировано 9 ноября 2013 года.
- statistics silk production
- Ален Эд, 2001, с. 176.
- Лю Ши-Ци. География сельского хозяйства Китая. — М.:: Изд-во иностранной литературы, 1957. — С. 360—371. — 404 с.
- Narendra Reddy. Silk: Materials, Processes, and Applications. — Woodhead Publishing, 2019. — P. 2. — 226 p.
- Ryszard M. Kozlowski, Maria Mackiewicz-Talarczyk. Handbook of Natural Fibres: Volume 1: Types, Properties and Factors Affecting Breeding and Cultivation. — Woodhead Publishing, 2020. — P. 738—739. — 838 p.
- Джерри Хопкинс. Экстремальная кухня. Причудливые и удивительные блюда, которые едят люди. — М.: ФАИР-ПРЕСС, 2006. — С. 224—227. — 336 с. — ISBN 5-8183-1032-9.
- Dr. Cherry Ron. Use of Insects by Australian Aborigines. Cultural Entomology Digest – Insect Articles. Дата обращения: 7 сентября 2012. Архивировано 16 октября 2012 года.
- Ален Эд, 2001, с. 179.
- Хестанова М. С., Кертанов С. Р., Макиев Г. Г. Редкие заболевания, вызываемые членистоногими: саркопсиллез, лепидоптеризм // Colloquium-journal. — 2019. — № 15 (39). — doi:10.24411/2520-6990-2019-10466.
- Гелашвили Д. Б., Крылов В. Н., Романова Е. Б. Зоотоксинология: биоэкологические и биомедицинские аспекты. — Нижний Новгород: Изд-во ННГУ, 2015. — С. 355—371. — 770 с.
- Cardoso A. E. C., Haddad V. J. Accidents caused by lepidopterans (moth larvae and adult): study on the epidemiological, clinical and therapeutic aspects (англ.) // Brazilian Annals of Dermatology. — 2005. — No. 80. — P. 573—580.
- Rosen T. Caterpillar dermatitis (англ.) // Dermatol Clin : Журнал. — 1990. — April (vol. 8, no. 2). — P. 245—252. — PMID 2191798.
- Lee D., Pitetti R. D., Casselbrant M. L. Oropharyngeal manifestations of lepidopterism (англ.) // JAMA : Журнал. — 1999. — January (vol. 125, no. 1). — P. 50—52. — doi:10.1001/archotol.125.1.50. — PMID 9932587.
- Eagleman David M. Envenomation by the asp caterpillar (Megalopyge opercularis) // Clinical Toxicology. — 2007. — Vol. 46. — P. 201—205. — doi:10.1080/15563650701227729. — PMID 18344102.
- Hossler Eric W. Caterpillars and moths // Dermatologic Therapy. — 2009. — Vol. 22. — P. 353—366. — doi:10.1111/j.1529-8019.2009.01247.x. — PMID 19580579.
- McGovern J, P., Barkin G. D., McElhenney Th. R., Wende R. Megalopyge opercularis: Observations of Its Life History, Natural History of Its Sting in Man, and Report of an Epidemic // JAMA. — 1961. — Vol. 175. — P. 1155—1158. — doi:10.1001/jama.1961.03040130039009.
- Goddard J., Zhou L. Physician’s guide to arthropods of medical importance, 5th Edition. — Philadelphia: CRC Press, 2007. — P. 159. — 480 p.
- Winkler D. Yartsa Gunbu (Cordyceps sinensis) and the fungal commodification of the rural economy in Tibet AR (англ.) // Economic Botany : Журнал. — 2008. — Vol. 62, no. 3. — P. 291—305. — doi:10.1007/s12231-008-9038-3.
- Halpern G.M., Miller A. P. Medicinal Mushrooms. — New York: M. Evans and Company, Inc., 2002. — P. 64—65. — 184 p.
- Lo H. C., Hsieh C., Lin F. Y., Hsu T. H. A systematic review of the mysterious caterpillar fungus Ophiocordyceps sinensis in Dong-ChongXiaCao (冬蟲夏草 Dōng Chóng Xià Cǎo) and related bioactive ingredients (англ.) // Journal of Traditional and Complementary Medicine : Журнал. — 2013. — Vol. 3, no. 1. — P. 16—32. — doi:10.4103/2225-4110.106538. — PMID 24716152.
Литература
- Герасимов А. М. Гусеницы. — М.—Л.: Издательство академии наук СССР, 1952. — Т. 1. — 343 с. — (Фауна СССР).
- Лавров С. Д. Наши гусеницы. Определитель. — М.: Государственное учебно-педагогическое издательство, 1938. — 144 с.
- Определитель насекомых Дальнего Востока России. В 6 т. / под общ. ред. П. А. Лера. — Владивосток: Дальнаука, 1997. — Т. V. Ручейники и чешуекрылые. Ч. 1. — С. 207—238. — 540 с. — 500 экз. — ISBN 5-7442-0986-7.
- Ламперт К. Атлас бабочек и гусениц. — Минск: Харвест, 2003. — 735 с. — 5000 экз. — ISBN 985-13-1664-4.
- Ален Эд, Мишель Виар. Бабочки мира. — 2-е, стереотипное. — Интербук-бизнес, 2001. — 193 с. — ISBN 5-89164-090-2.
- The Book of Caterpillars: A Life-Size Guide to Six Hundred Species from Around the World. — Chicago: University of Chicago Press, 2017. — 656 с. — ISBN 978-0-226-28736-2.
- Sterling Ph., Henwood B., Lewington R. Field Guide to the Caterpillars of Great Britain and Ireland (Field Guides). — Bloomsbury Wildlife, 2020. — 448 с. — ISBN 978-1472933560.
- Carter D. J., Hargreaves B. A field guide to caterpillars of butterflies and moths in Britain and Europe. — Collins, 2001. — 296 с. — ISBN 978-0002190800.
- Miller J. C., Janzen D. H., and W. Hallwachs. 100 Caterpillars: Portraits from the Tropical Forests of Costa Rica. — Harvard University Press, 2006. — 272 с. — ISBN 978-0674034822.
- Minno M. C., Butler J. F., and D. W. Hall. Florida Butterfly Caterpillars and their Host Plants. — University Press of Florida, 2005. — 360 с. — ISBN 978-0-8130-2789-0.
- Pierce N. E., Braby M. F., Heath A., Lohman D. J., Mathew J., Rand D. B., Travassos M. A. The Ecology and Evolution of Ant Association in the Lycaenidae (Lepidoptera) (англ.) // Annual Review of Entomology : Журнал. — 2002. — Vol. 47. — P. 733—771. — ISSN 0066-4170.
Ссылки
- Изображения гусениц на Insecta.pro (англ.)
| В библиографических каталогах |
|---|
