Нимфалиды
Нимфалиды[2][3] (лат. Nymphalidae) — семейство чешуекрылых, состоящее из около 6100 видов[4]. Название семейству впервые было дано немецким зоологом, ботаником и специалистом по мезоамериканским языкам, Константином Самуэлем Рафинеск-Шмальцем. Основным отличительным признаком представителей семейства нимфалид является укороченная передняя пара ног. Данный признак наблюдается практически у всех таксонов семейства, за редким исключением.
| Нимфалиды | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Отряд: Подотряд: Инфраотряд: Клада: Клада: Apoditrysia Клада: Obtectomera Надсемейство: Семейство: Нимфалиды |
||||||||||||
| Международное научное название | ||||||||||||
| Nymphalidae Rafinesque, 1815 | ||||||||||||
| Подсемейства | ||||||||||||
|
||||||||||||
| ||||||||||||
Общая характеристика

Бабочки средних и крупных размеров. Размах крыльев представителей семейства 25—180 мм, у большинства — 50—80 мм. Бабочки из этого семейства характеризуются богатым разнообразием окраски[5] и рисунка крыльев. Наиболее ярко и пёстро окрашены представители подсемейства Nymphalinae. Нижняя сторона крыльев часто покровительственной окраски[5], порой подражающая сухим листьям и другим сухим растительным остаткам.
Голова округлая. Глаза голые или волосистые. Усики головчатые или булавовидные[5], достигающие половины костального края переднего крыла. Губные щупики большие и мощные, с коротким концевым сегментом, косо торчащие вперед. Грудь мощная и развитая.

Отличительной особенностью представителей семейства являются передние ноги, которые и у самцов и самок редуцированы (недоразвиты) и не используются при хождении[6][7]. Передние ноги лишены коготков и покрыты густыми волосками. Задние голени с одной парой шпор. Крылья широкие, их наружный край часто с угловатыми выступами или зазубринами. Переднее крыло треугольной формы, с выпуклым, несколько вогнутым или зубчатым (волнистым) внешним краем. На переднем крыле жилки R3, R4 и R5 ответвляются от общtго ствола, который берёт начало от внешнего края центральной ячейки. В ряде случаев от этого же ствола ответвляется и жилка R2 (Melitaeinae). Жилка R3 доходит к вершине крыла, a R4 и R5 — к внешнему краю. Жилка R1 имеет самостоятельное начало от переднего края центральной ячейки. Сама центральная ячейка замкнутая (кроме Neptis). Анальная жилка одна. Заднее крыло округлой или угловатой формы, с двумя развитыми анальными жилками. Центральная ячейка заднего крыла не замкнутая (кроме Argynninae). Задние крылья, например у хараксиновых могут иметь «хвостики».
Брюшко узкое и короткое. Гениталии самцов характеризуются значительным разнообразием, а иногда и высокой сложностью строения. Тегумен относительно крупный, имеет вид купола, сильно выступает вперёд. Ункус может иметь разнообразное строение — от сложного у некоторых Argynninae до полной редукции у Melitaeinae. Части гнатоса не всегда хорошо дифференцируются, иногда отсутствуют. Саккус короче ункуса, у Melitaea он уплощенный, имеет различную форму центральной вырезкой. Вальва широкая либо, со сложной структурой внутренней поверхности у Argynninae, мощно развитыми гарпами и характерным отростком у Melitaea. Эдеагус прямой или изогнутый, имеет заостренную или тупую вершину. Для строения гениталий самок характерно разнообразие строения вагинальной области. Генитальная пластинка не всегда дифференцируется. Бурса перепончатая, всегда без аппендикса[8].
Особенности биологии и экологии
Почти все нимфалиды — активные посетители цветущих растений. Ряду видов из семейства для жизнедеятельности необходимы микроэлементы, в первую очередь натрий. Они охотно летят на влажную минеральную глину, на экскременты и мочу крупных животных, влажный древесный уголь, человеческий пот — откуда получают влагу и необходимые микроэлементы. Часто самцы данных видов собираются группами на влажном песке и глинистых почвах, по берегам ручьёв, возле луж.
Гусеницы средних или крупных размеров, 30—50 мм длиной, различной окраски — от тёмных цветов до зелёной, с различным рисунком, умеренно волосистые, с шипами или выростами на теле. Гусеницы младших, а иногда и средних возрастов часто живут группами, однако есть и такие, которые живут поодиночке. Питаются травами, листьями кустарников и деревьев, в том числе плодовых, иногда нанося заметный вред. Куколки угловатые или в бугорках, висят на листе или стебле кормовых растений или рядом с ними на субстрате[5].
Для некоторых нимфалид характерно территориальное поведение, как правило, отмечаемое у самцов. При этом они могут занимать в ожидании самок наивысшую точку ландшафта, либо же облетать свой участок обитания. Часто что со своих территориальных участков эти самцы прогоняют не только других самцов своего вида, но и представителей других видов бабочек со схожей окраской. В ряде случаев самцы воспринимают в качестве соперников неодушевлённые предметы соизмеримые с бабочками их вида размерами и цветом[9] — например, данное поведение отмечается у самцов морфо, хараксес и других.
У многих видов выражен половой диморфизм в размерах и окраске[5].
Миграции
Шашечницы и малые перламутровки преимущественно оседлые и держатся, как правило, на определённых участках своего места обитания. Многие же виды нимфалид склонны к миграционному поведению в поисках пищи и кормовых растений для потомства. Некоторые способны при миграциях преодолевать большие расстояния, например, репейница. При миграциях репейницы летят группами, со скоростью 25—30 км/ч и могут преодолевать до 500 км в день[10]. Бабочки летнего поколения репейниц зимуют в Северной и Центральной Африке, где они размножаются и уже новое поколение мигрирует на север, где выводятся летние поколения бабочек. В конце лета особи одного из этих поколений мигрируют обратно в Африку[11]. Весной цикл повторяется снова. Общая протяженность их перелета достигает 4000-5000 км[11]. Как и все насекомые данный вид периодически дает массовые вспышки численности в результате массового размножения резко увеличивает количество мигрирующих особей. Таким образом ежегодно особи репейниц совершают огромные массовые миграции из поколения в поколение. В период миграций бабочки продвигаются на север широким фронтом от Урала до Скандинавии[11], повсеместно достигая побережий окраинных морей Северного Ледовитого океана[12]. В соответствующих районах на севере они образуют временные поселения (псевдопопуляции) и успешно размножаются. Миграции позволяют бабочкам воспользоваться изменениями ресурсов по мере смены сезонов, перемещаясь на север летом, чтобы избежать жарких температур в Африке, и снова перемещаясь на юг, чтобы избежать холодных зим в северной Европе[11][13]. Эта картина также отражена в США, где репейницы мигрирует из Мексики в северную часть Соединенных Штатов и Канады[14].
Ареал и виды
Около 6100 видов и 559 родов[4]. Широко распространены по всему миру, большей части в тропиках. В России более 130 видов[5].
Замечания по систематике
Состав семейства и его систематика является предметом научных дискуссий и исследований. Многие таксоны, рассматриваемые сейчас в ранге подсемейств Нимфалид, как то Danainae, Heliconiinae, Libytheinae, Morphinae, ранее рассматривались в ранге самостоятельных семейств[15]. Спорным является систематическое положение группы Сатириды. Ряд энтомологов включают данный таксон в ранге подсемейства в составе семейства Нимфалид — именно в таком ранге рассматриваются представители данной группы в англоязычной научной литературе. На территории стран бывшего СССР, по традиционной систематике, на основании характерного узнаваемого внешнего облика и особенностей морфологии гениталий самцов за этой группой бабочек преимущественно сохраняют статус самостоятельного семейства[16][17].
Филогения и эволюция
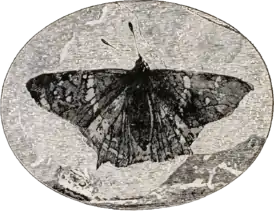
Филогения нимфалид сложна. Некоторые таксоны до сих пор имеют неясную позицию, что отражает тот факт, что некоторые подсемейства ранее были признаны в качестве отдельных семейств из-за недостаточного изучения. Наиболее древний известный ископаемый представитель семейства — Prodryas persephone, известный по единственной необычайно хорошо сохранившейся окаменелости, обнаруженной в 1878 году в отложениях Северной Америки[18]. Окамелесть P. persephone найдена в отложениях позднего эоцена в известной флориссантской формации у Флориссант (Колорадо)[19][20]. Длина переднего крыла бабочки 24,5 мм. Особь полностью сохранилась, включая ротовой аппарат, антенны и даже цветовой рисунок чешуек на крыльях[21]. Ближайшие родственные нимфалиды происходят из современных родов Hypanartia[22] и Antanartia[23].
Представители подсемейств
_male_(35510927695).jpg.webp)
.jpg.webp) Catonephele orites
Catonephele orites
Biblidinae




_Dos.jpg.webp)




Примечания
- Ряд энтомологов выделяют данный таксон в ранге подсемейства Satyrinae в составе семейства нимфалиды — именно в таком статусе рассматриваются представители данной группы в зарубежной литературе. На территории стран бывшего СССР, по сохранившейся систематике, на основании особенностей анатомии и биологии, а также характерного узнаваемого внешнего облика, за этой группой бабочек обычно сохраняют статус семейства.
- Горностаев Г. Н. Насекомые СССР. — Москва: Мысль, 1970. — 372 с. — (Справочники-определители географа и путешественника).
- Определитель насекомых Дальнего Востока России. Т. V. Ручейники и чешуекрылые. Ч. 5 / под общ. ред. П. А. Лера. — Владивосток: Дальнаука, 2005. — 575 с. — 500 экз. — ISBN 5-8044-0597-7.
- Van Nieukerken, Erik J. et al. Order Lepidoptera Linnaeus, 1758 (англ.) // In: Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. — Zootaxa. — Magnolia Press, 2011. — Vol. 3148. — P. 212—221. — ISSN 1175-5326.
- Девяткин, 2013.
- Wolfe, Joanna M.; Oliver, Jeffrey C.; Monteiro, Antónia. Evolutionary reduction of the first thoracic limb in butterflies (англ.) // Journal of Insect Science : journal. — 2011. — 1 January (vol. 11, no. 1). — P. 66. — ISSN 1536-2442. — doi:10.1673/031.011.6601. — PMID 21867433.
- Bingham, C. T. The Fauna of British India, Including Ceylon and Burma: Butterflies Volume I (англ.). — London: Taylor and Francis, Ltd., 1905.
- Некрутенко Ю. П. Булавоусые чешуекрылые Крыма. Определитель. — Киев: Наукова думка, 1985. — 152 с.
- Тинберген Н. Поведение животных. — М.: Мир, 1978. — 192 с., илл. с.
- Harlan A.Ch., A Quantitative Study of the Migration of the Painted Lady Butterfly, Vanessa Cardui L. Ecology, Vol. 32, No. 2 (Avril 1951), pp. 155—171.
- Gerard Talavera, Roger Vila. Discovery of mass migration and breeding of the painted lady butterfly Vanessa cardui in the Sub‐Sahara: the Europe-Africa migration revisited. Biological Journal of the Linnean Society, Volume 120, Issue 2, 2017, pp. 274—285, https://doi.org/10.1111/bij.12873
- Болотов И. Н. Многолетние изменения фауны булавоусых чешуекрылых (Lepidoptera, Diurna) северной тайги запада Русской равнины. — Экология, 2004. № 2. С. 141—147
- Southwood T. R. E. Habitat, the templet for ecological strategies? Journal of Animal Ecology. 1977. № 46. р. 337—365.
- Stefanescu, C., R. Askew, J. Corbera, M. Shaw. 2012. Parasitism and migration in southern Palaearctic populations of the painted lady butterfly, Vanessa cardui (Lepidoptera: Nymphalidae). European Journal of Entomology, 109/1: 85-94.)
- Smart P. The Illustrated Encyclopedia of the Butterfly World. — Transworld Publishers Ltd, 1981. — 274 p. — ISBN 0-552-98206-7. (англ.)
- Кузнецов В. И., Стекольников А. А. 2001. Сравнительная морфология гениталий самцов в подсемействах Morphinae и Brassolinae и восстановление сем. Satyridae, status resurr. (Lepidoptera) в новом объёме // Энтомологическое обозрение Т. 80, вып. 1. С. 121—136.
- Сочивко А.В., Каабак Л.В. Определитель бабочек России. Дневные бабочки. — М.: Аванта+, 2012. — С. 106—107. — 320 с. — (Мир энциклопедий). — 5000 экз. — ISBN 978-5-98986-669-4.
- H. Goss & W. W. Fowler. Societies: Entomological Society of London (неопр.) // The Entomologist's Record and Journal of Variation. — 1894. — Т. 5. — С. 19—20.
- Thomas C. Emmel, Marc C. Minno & Boyce A. Drummond. The fossil butterflies of Florissant // Florissant butterflies: a guide to the fossil and present-day species of central Colorado (англ.). — Stanford University Press, 1992. — P. 4—12. — ISBN 9780804720182.
- Steven W. Veatch & Herbert W. Meyer. History of paleontology at the Florissant fossil beds, Colorado // Paleontology of the Upper Eocene florissant formation, Colorado (англ.) / Herbert William Meyer & Dena M. Smith. — Geological Society of America, 2008. — P. 1—18. — (Volume 435 of Special papers). — ISBN 9780813724355.
- Гримальди, Дэвид & Michael S. Engel. Fossil insects // Evolution of the insects (неопр.). — Cambridge University Press, 2005. — С. 42—92. — (Volume 1 of Cambridge Evolution Series). — ISBN 9780521821490.
- Niklas Wahlberg (2006). «That awkward age for butterflies: insights from the age of the butterfly subfamily Nymphalinae (Lepidoptera: Nymphalidae)». Systematic Biology. 55 (5): 703—714. doi:10.1080/10635150600913235. PMID 16952908
- Keith R. Willmott, Jason P. W. Hall & Gerardo Lamas (2001). «Systematics of Hypanartia (Lepidoptera: Nymphalidae: Nymphalinae), with a test for geographical speciation mechanisms in the Andes». Systematic Entomology. 26 (4): 369—399. doi:10.1046/j.1365-3113.2001.00157.x
Литература
- Нимфалиды / Девяткин А. Л. // Николай Кузанский — Океан. — М. : Большая российская энциклопедия, 2013. — С. 51. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 23). — ISBN 978-5-85270-360-6.
Ссылки
- Нимфалиды (Nymphalidae) Архивная копия от 5 мая 2009 на Wayback Machine на lepidoptera.ru
| Таксономия | |
|---|---|
| В библиографических каталогах |
