Гаплогруппа R1a (Y-ДНК)
Гаплогруппа R1a (М420) — Y-хромосомная гаплогруппа, распространённая в Восточной и Центральной Европе, Средней и Южной Азии и Южной Сибири.
| Гаплогруппа R1a | |
|---|---|
| Тип | Y-ДНК |
| Время появления | 22,8 тысячи лет BP[1] |
| Место появления | Южная Сибирь[2] |
| Время до БОП | 18,3 тысячи лет[1] |
| Предковая группа | R1 |
| Сестринские группы | R1b |
| Субклады | Гаплогруппа R1a1 (Z282 (Европа) и Z93 (Азия)), R1a2 |
| Мутации-маркеры |
R1a : L62, L63, L120, M420, M449, M511, M513 R1a1a : M17, M198, M512, M514, M515, L168, L449, L457, L566 |
R1a возникла около 22 тыс. лет назад[3] (по другим данным — около 25 тыс. лет назад[4]) от мутации гаплогруппы R1, произошедшей у мужчины, жившего около 22,8 тысячи лет назад (дата определена по снипам компанией YFull[5]) предположительно в Азии (исходя из распространения линий R2 и R*). R1a-М420 делится на ветви R1a1-M459 и R1a2-YP4141. R1a2-YP4141 делится на две ветви R1a2a-YP5018 и R1a2b-YP4132.
Мутация SNP R1a-M420 была обнаружена после R1a1a-M17, что привело к реорганизации линии, в частности, к созданию новой парагруппы (обозначенной R-M420*) для относительно редких линий, которые не находятся в субкладе R1a1-M459/SRY10831.2, ведущем к линии R1a1a-M512/M17. Субклад R1a1a1-M417 имеет древность 5800 лет[4]. Распределение субкладов R1a1a1b1a-Z282 (включая R1a1a1b1a2-Z280) в Центральной и Восточной Европе и R1a1a1b2-Z93 в Азии предполагает, что R1a1a-M17 выделилась в пределах евразийских степей. Место происхождения и развития этих субкладов значимо для определения происхождения и путей расселения протоиндоевропейцев.
Гипотезы происхождения R1a
Происхождение R1a1a1 (M417), древние миграции
Корневая и первая ветвь, пошедшая от гаплогруппы R1a — M420. Она разделилась между 25 и 22 тыс. лет назад. Это период последнего ледникового максимума. В исследовании от 2014 года, проведённом Питером А. Андерхиллом и соавторами с участием 16 244 человек из более чем 126 популяций со всей Евразии, авторы пришли к предположению о том, что «начальные эпизоды диверсификации гаплогруппы R1a, вероятно, произошли в окрестностях современного Ирана». Согласно расчётам Андерхилла и др. (2014), субклад R1a1a1-M417, диверсифицировался в R1a1a1b1a-Z282 и R1a1a1b2-Z93 около 5800 лет назад[6].
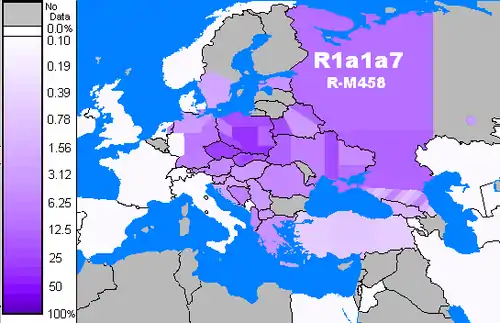
R1a показывает сильную корреляцию с индоевропейскими языками Южной и Западной Азии, Центральной и Восточной Европы[7][8]. Наибольшее распространение она имеет в Восточной Европе, Западной и Южной Азии и Центральной Азии. В Европе преобладает R1a1a1b1a-Z282, в то время как в Азии доминирует R1a1a1b2-Z93. Связь между Y-хромосомной гаплогруппой R1a1a-M17 и распространением индоевропейских языков впервые была отмечена Татьяной Зерджаль и её коллегами в 1999 году[9].
Взрывообразный рост числа потомков основателя субклады R1a1a1b2-Z93 произошёл в Южной Азии примерно 4,5—4 тыс. лет назад. Расширение маркеров R1a1a1b2-Z93 в промежуток между 4500 и 4000 лет назад на несколько столетий предшествует угасанию и краху Индской цивилизации[10].
Согласно исследованию В. Нарасимхана и др. (2018), степные скотоводы являются вероятным источником Y-хромосомной гаплогруппы R1a в Индии[11].
По данным на 2017—2018 год по исследованиям, проводимым Европейскими генетиками, известно следующее.
1 — Гаплогруппа R1a через гаплогруппу R1 происходит от старшей по возрасту гаплогруппы R. Гаплогруппа R произошла в Северной Азии перед Последним Ледниковым максимумом, в промежутке между 26 599 и 19 000 лет назад. Наиболее древний найденный образец данной гаплогруппы (R*) найден у мальчика со стоянки Мальта в Иркутской области. Этот мальчик жил в племени охотников на мамонтов — племена специализировались на одном из видов животных того периода — мамонты, олени, бизоны и так далее. В силу постоянных миграций эти носители гаплогруппы R доходили до современной Европы и Южной Азии и таким образом способствовали её распространению. В современный период именно эти регионы наиболее богаты данной гаплогруппой и её производными — R1b в Западной Европе, R1a в Восточной Европе, Центральной и Южной Азии и R2 в Южной Азии.
2 — В результате серии мутаций корневой гаплогруппы R развилась гаплогруппа R1 и из неё — R1a. Срок возникновения — приблизительно после последнего ледникового максимума. Точное место возникновения именно гаплогруппы R1a на данном этапе изучения не известно. Разброс возможных географических регионов — Пакистан, Северо-Западная Индия, Балканы — данные регионы имеют наибольшее генетическое разнообразие этой гаплогруппы.
3 — Существует версия, что на Балканы гаплогруппа R1a попала от миграционных потоков, идущих из Евразийских степей, и в силу того, что миграции шли волнами, было обеспечено высокое разнообразие мутаций. В Южной Азии уже на протяжении 10 000 лет плотность и количество населения наибольшие на планете и потому разнообразие гаплогруппы так же велико. Исходя из этого, генетики предполагают, что гаплогруппа R1а могла возникнуть либо в Средней Азии, либо в Южной России — в Сибири.
При первом варианте R1a с носителями сначала попала в Восточную Европу. Второй вариант — предварительно люди мигрировали в период палеолита на юг, в Среднюю Азию, затем — в Иран, затем — на Кавказ во время неолита, после Кавказа через Каспийские степи уже попали в Восточную Европу. За данный вариант говорит то, что в Иране обнаруживаются очень старые субклады R1a — М420, в регионе Загрос. При третьем варианте развития событий люди из места происхождения гаплогруппы разделились на два потока — один в Казахстанские степи и затем в Восточную Европу, другой отклонился и попал в Иран.
4 — Ряд теорий, упиравших на то, что гаплогруппа R1a попала в Европу через Анатолию в период миграции неолитических земледельцев, натолкнулись на несостыковки. 99 % современной R1a происходит от гаплогруппы R1a-M417 — а она, в свою очередь, имела расширение в период Бронзового века, но не в период неолита. Филогенетика данной гаплогруппы выводит к месту её происхождения — Восточной Европе. Большая часть R1a на Ближнем востоке — глубокая субклада, отделившаяся от гаплогруппы R1a-Z93, имевшей место происхождения в России.
При исследовании образцов, максимально близких к неолитическим земледельцам, не обнаружено мутаций, связывающих их с R1a, даже относительно поздний земледелец Эци, живший 5 300 лет назад, не несёт в себе никаких признаков гаплогруппы R1a (G2a), так же R1a отсутствует у современных сардинцев, являющихся наиболее близкими потомками земледельцев, мигрировавших в период неолита. Данные факты указывают, что гаплогруппа могла произойти именно в Восточной Европе и затем в период Бронзового века распространиться.
Теория происхождения из Азии
Квислед в 2003 году предположил либо Южную, либо Западную Азию, а Мирабал в 2009 году указывал на Южную и Центральную Азии[7]. Другие исследователи предполагают происхождение R1a на территории современной Украины[12][13], в Средней Азии[14] и в Западной Азии[15][16].
В 2000 году предположено происхождение на территории Украины и постледниковое распространение R1a1 во время последнего ледникового максимума. Распространение генома по этой теории обеспечивалось в том числе благодаря экспансии курганной культуры в Европе и на Восток[17].
Спенсер Уэлс предлагает центрально-азиатское происхождение, предполагая, что распределение и возраст R1a1 указывают на древнюю миграцию, соответствующую распространению Культуры курганных погребений в их экспансии из Евразийской степи[18].
В 2012 году предположено, что R1a1a выделилась в евразийских степях или на Ближнем Востоке и на Кавказе. Внутренняя и Центральная Азия, является перекрывающейся зоной для линий R1a1-Z280 и R1a1-Z93, что подразумевает раннюю зону дифференциации R1a1a-M198, предположительно произошла в пределах Евразийских степей или на Ближнем Востоке и на Кавказе, поскольку они лежат между Южной Азией и Центральной и Восточной Европой[19].
Три генетических исследования, проведённые к 2015 году, подтвердили курганную гипотезу Гимбутас. Согласно этим исследованиям, гаплогруппы R1b и R1a, теперь наиболее распространенные в Европе, были распространены из российских степей наряду с индоевропейскими языками. Исследования также обнаружили аутосомный компонент, присутствующий в современных европейцах, которого не было у неолитических европейцев, которые были бы введены с отцовскими линиями R1b и R1a, а также с индоевропейскими языками[20][21][22].
_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg.webp)
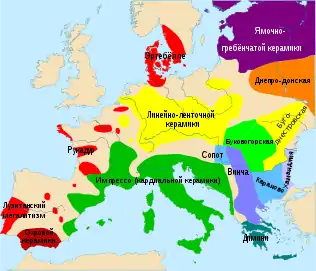 Европейский средний неолитический период. Культура ямочно-гребенчатой керамики, между 4200 г. до н. э. - ок. 2000 г. до нашей эры.
Европейский средний неолитический период. Культура ямочно-гребенчатой керамики, между 4200 г. до н. э. - ок. 2000 г. до нашей эры. Культура боевых топоров - с. 2900 до н.э. – по. 2350 до н.э.
Культура боевых топоров - с. 2900 до н.э. – по. 2350 до н.э. Трипольская культура - границы 1.
Трипольская культура - границы 1. Трипольская культура - границы 2.
Трипольская культура - границы 2.
Связь с ямной культурой
Дэвид Энтони считал людей ямной культуры предками индоевропейцев[23][24]. Согласно исследованию 2015 года, массовая миграция людей ямной культуры имела место около 2500 лет до нашей эры. Её связывают с тем, что данная культура одной из первых массово научилась использовать лошадей для передвижения людей и перевозки грузов, а также активно применять бронзовое оружие. Генетики из Стэнфордского университета и др. (Poznik et al. 2016) выявили, что с демографической экспансией в Западную Европу в бронзовом веке связана не гаплогруппа R1a, а наплогруппа R1b1a1a2a1a-L151/L11 (L23 и её субклады), в то время как субклад R1b-Z2103, представленный в ямных погребениях, в этой экспансии не участвовал[25][26].
Ямную культуру сменила катакомбная культура, у представителей которой также определили Y-хромосомную гаплогруппу R1b, а не R1a[27][28].
Выделение германских племён
Первое сильное распространение гаплогруппы R1a коррелирует с миграциями именно культуры боевых топоров. С этой миграцией в Германию и Нидерланды попал субклад R1a1a1b1-Z283. В Скандинавию — R1a1a1b1a3-Z284. Люди этой культуры смешались с племенами, населявшими Германию до их прихода и носившими гаплогруппы I1 и I2, таким образом образовав первую индоевропейскую культуру на территориях нынешних Германии и Скандинавии. Однако эта культура не была первично германской, а была первичной индоевропейской общностью и скорее всего несла в себе основы и для германцев и для прото-балто-славян. Языковой анализ подтверждает такой подход — первичный германский язык развивался как смесь прото-балто-славянских языков культуры шнуровой керамики (R1a1a1b1-Z283) и более поздней унетицкой культуры, люди этой культуры были гибридными носителями R1a/R1b.
Эта первая волна миграции большей частью состояла из носителей R1b, но в её массе были также племена, носившие R1a в виде субклады R1a1a1a1-L664. Эти племена смешались затем с носителями R1b, в особенности с её субкладой R1b1a1a2a1a1-U106, от которой спустя много веков произойдут в итоге германцы. Данная миграция поглотила и закончила в итоге культуру колоколовидных кубков и культуру шнуровой керамики в Центральной Европе. Из получившегося процесса вышла унетицкая культура и стала основой для германских, кельтских и итальянских культур.
Унетицкая культура расширила ареал для Скандинавии и таким образом дала начало скандинавскому Бронзовому веку. В этот же период люди, носившие R1a1a1a1-L664 и R1b (L11 и U106), достигли Скандинавии. Люди в тот период в Скандинавии говорили на первично германском языке, основанном на собственном лексиконе, лексике поглощённой Культуры шнуровой керамики, являвшейся носителем R1a1a1b1a3-Z284 и говорившей на смеси прото-балто-славянских языков и третий компонент это носители неиндоевропейских языков и гаплогрупп G2a, I1, I2. В итоге современная Скандинавия состоит из 40 % I1, 20 % R1a и 40 % R1b.
Выделение славянских племён
Миграция из Ямной культуры через Северо-Восточную Европу и связанная с этим Культура шнуровой керамики. Эти племена заселили наиболее плотно нынешние страны — Польшу, Белоруссию и Украину вкупе с Западной Россией, образовав ядро прото балто-славян. Высокая концентрация гаплогруппы R1a в балтийских и в славянских странах связана не только с миграцией этой культуры, а также объясняется несколькими последовавшими волнами переселений с территории нынешней России, последняя из которых имела место между 5 и 10 веками нашей эры[25][26].
В процессе движения Культуры шнуровой керамики она поглотила в себя Трипольскую культуру на Западной Украине и в Северо Восточной Румынии, люди которой были носителями G2a-U1 и I2a1b-M423. Люди Трипольской культуры, таким образом, были непосредственными потомками европейцев времён Палеолита. В них также содержались следы от генетики земледельцев с Ближнего востока — гаплогруппы V13, J2a и T1a. Культура шнуровой керамики в период своего развития соседствовала в степях между Днепром и Уралом со Срубной культурой и в Польше с Тшинецкой культурой. Последняя славянская миграция, внесшая заметный вклад, произошла начиная с 6-го века нашей эры с территорий Украины до Польши, Словакии и Словении. Эта миграция заполнила людьми образовавшиеся свободные пространства, которые получились после миграции и вторжения древних восточных германских племён в Римскую империю[29].
Выделение балтийских племён
Балтийская ветвь народов развилась из нескольких культур входивших в культуру шнуровой керамики и боевых топоров, в том числе, по мнению нескольких историков, в балтских племенах заметны схожие с фатьяновской культурой 3200 — 2300 лет до н. э. культурные традиции, включая керамику. Фатьяновская культура (R1a1a1b2-Z93) является восточным вариантом культуры боевых топоров — шнуровой керамики, который появился на Северо-Востоке Европы. В раннем бронзовом веке кочевники, носители R1a, смешались с носителями уральских языков N1c1. Этот подход подтверждается наличием гаплогрупп R1a и N1c1 в Южной Финляндии, до Литвы и на Северо-Западе России. Латышские и литовские носители R1a так же несут в себе балто-славянские линии, такие как R1a1a1b1a1-M458, R1a1a1b1a2b-CTS1211 и R1a1a1b1a2a-Z92.
Индо-иранское выделение R1a
Носители протоиндоиранского языка, впоследствии ставшие ариями в «Ригведе» и «Авесте», возникли в синташтинской культуре, 2100—1750 лет до н. э. в долинах рек Тобол и Ишим к востоку от Уральских гор. Синташтинская культура была наследницей более старой культуры — абашевской 2500—2900 лет до н. э., пришедшими с Верхенго Дона и Волги на Урал. Так же в образовании синташтинской культуры принимала участие Полтавская культура 2700—2100 лет до н. э. Синташтинская культура и её носители R1a1a1b2-Z93 были первыми носителями индоевропейского языка к западу от Урала в период бронзового века. Эти племена в итоге вышли к обширным равнинам и пустыням Средней Азии и дошли до богатых металлами Алтайских гор. Арии быстро распространились по всей Центральной Азии, от берегов Каспийского моря до Южной Сибири и Тянь Шаня. Этому способствовала активная торговля, сезонные миграции населения вслед за стадами и грабительские походы. Именно синташтинцам приписывают одни из самых ранних образцов боевых колесниц — 2100 лет до н. э. Благодаря колесницам они смогли распространиться на богатый металлами горнодобывающий регион Бактрию — современная граница Туркменистана, Узбекистана, Таджикистана и Афганистана. В итоге они могли добывать большое количество меди на Урале и протоиндоиранцы из синташтинской культуры активно экспортировали её на Ближний Восток.
Индоиранцы освоили Заревшанскую долину, основав в итоге Петровскую медно-рудную колонию в Тугае в 1900 году до н. э. из-за большого содержания в ней медной руды. Олово добывалось в Карнабе и Мушистоне. Олово во времена позднего бронзового века было ценным ресурсом — медно-оловянный сплав прочнее на излом и удар, чем обычная бронза с мышьяком. К 1700 годам до н. э. индоиранцы расширили ареал до нижней долины Амударьи и основали общины, занимавшиеся земледелием с орошением — тазабагьябская культура. К 1600 годам до н. э. старые города Бактрии были заброшены в силу новых волн степных кочевников с севера, которые наступали на поселения. Индоиранской культурой в Центральной Азии являлась андроновская культура, существовавшая до 800 г. до н. э. Индо иранцы продвигались дальше на юг и достигли Гиндукуша.
К 1700 г. до н. э. кочевники-скотоводы проникли в Белуджистан — Юго-Западный Пакистан. В долину реки Инд они пришли к 1500 г. до н. э. Северная и центральная части Индийского субконтинета были заселены ими к 500 г. до н. э. Взаимопроникновение культуры степных скотоводов, имевших на вооружении колесницы и носителей Индской культуры и языка, привело к взаимному обмену знаниями и языками. В итоге к 1600 г. до н. э. была захвачена Ассирия и инды стали правителями Митанни к 1500 г. до н. э. Мидийцы, парфяне и персы — все носители иранских языков из Андроновской культуры — перешли жить на Иранское плато к 800 г. до н. э. Оставшиеся в Центральной Азии племена стали в итоге скифами, таковыми их и зафиксировала историческая наука, зародившаяся в Древней Греции.
Оставшиеся после миграций представители Ямной культуры, которые обитали в Понтийско — Каспийских степях, стали сарматами и так же были зафиксированы в исторической науке Древней Грецией и Древним Римом. Эти перемещения и миграции населения привели к распространению гаплогруппы R1a в Южной части Центральной Азии и в Индийском субконтиненте. Наиболее высокая концентрация R1a — до 65 % — наблюдается на стыке границ Киргизии, Таджикистана и Северного Афганистана. В Индии и Пакистане.
R1a составляет от 15 до 50 % генетического материала у населения в заивисимости от региона, этнической группы и кастового сословия. R1a наиболее часто представлена на Северо-Западе Индийского субконтинента, наименее часто — На юге — Тамил, Керала, Карнатака, Андхра-Праде, Бенгалия. Более 70 % браминов — высшей касты в Индии — несут в себе значительную часть R1a1. Однако митохондриальные ДНК-линии в Южной Азии в подавляющем большинстве случаев являются доиндоевропейскими. Например, в Индии более 75 % мтДНК М и R — это местные линии и только 10 % — Восточно-Азиатские линии. Оставшиеся 15 % гаплогрупп — это 7 — 8 % ближневосточных линий и только 7 % могут быть отнесены к линиям, попавшим с носителями из Понтийско-Каспийских степей, в основном в форме гаплогрупп U2 и W. Европейские линии мтДНК гораздо чаще представлены в Центральной Азии, в Афганистане и на Севере Пакистана. Первые крупные поселения, в которых постоянно проживали женщины индо-арийской языковой группы и генетики, это Северный Пакистан, Западная Индия — Пенджаб и Гуджарата и Северная Индия — Уттар — Прадеш, где гаплогруппы U2 и W являются наиболее распространенными сегодня.
Таримские мумии
В 1934 году шведский археолог Фольке Бергман на северо-западе Китая, в районе реки Тарим (ныне Синьцзян-Уйгурский автономный район), обнаружил около 200 мумий европеоидного типа. Наиболее старая из мумий датируется примерно 2 тыс. лет до н. э. При анализе ДНК у 11 из 12 мумий мужчин была найдена гаплогруппа R1a1 по нестабильному снипу M17. Современные жители Таримского бассейна — уйгуры. Они несут в себе до 20 % R1b1a1a1-M73 и 30 % R1a1. У таримский мумий определили митохондриальные гапогруппы U4, U4’9, U5b2c, U5a2, T1a1b, I1b, G3b, H15b1, D4j1b и A17.
Есть две базовые теории о происхождении этой культуры. Первая — это группа ранних скотоводов из репинской культуры 3700 — 3300 лет до нашей эры, которые мигрировали из Донских и Поволжских степей до гор Алтая, затем основали афанасьевскую культуру 3600—2400 лет до нашей эры и уже от неё распространились до Таримского бассейна. Вторая — таримские мумии, это люди, происходившие из протоиндо-иранского народа, которые мигрировали по всей Центральной Азии примерно 2000 лет до нашей эры из синташтинской культуры, что не подверждается ископаемой ДНК.
Исследование 2021 года показывает, что популяция, связанная с мумиями, восходит не к индоевропейцам, а к древним северным евразийцам и была значительно изолирована от соседей начиная с 9 тыс. л. н. 4,5 тыс. л. н. Она стала первыми обитателями бассейна Тарима. У них определили Y-хромосомную гаплогруппу R1b1c-PH155/PH4796.
Гунны
При исследовании ДНК установлено, что гунны — это народ, появившийся при смешивании европейских носителей и северо-востока Азии. При этом большая часть генетики в гуннах была от Хунну — древнего кочевого народа, жившего в Китае. Современные жители территорий, где раньше жили Хунну, несут 90 % монгольских родов против 10 % европейских. Самая старая находка мтДНК европейского рода найдена на границах Монголии и рядом с озером Байкал, датируется 4000 лет до нашей эры. Данные выводы свидетельствуют, что носители тюркского языка быстро заменили языки скифов и иранские диалекты по всей Центральной Азии.
Таким образом, многие народы переняли тюркский язык, но оставались почти не затронуты тюрками в плане генетики. В итоге болгарский язык, хоть и назван в честь тюркского племени — булгар, не является тюркским. Это славянский язык с незначительным влиянием тюркского. Венгерский язык, иногда ошибочно принимаемый за язык, оставшийся от гуннов-хуннов, на самом деле является языком уральской группы — мадьяр. Тюркский язык, несмотря на огромную площадь распространения и большое количество разных народов-носителей, лингвистически не сильно отличается между носителями и разные народы понимают его довольно хорошо. Это связано с относительно недавним распространением языка и кочевым характером его носителей.
При этом генетическое совпадение между представителями ямной культуры и культуры боевых топоров достигает 75 %. Исследователи отмечают, что R1a и R1b могли распространиться в Европу с Востока после 3000 лет до нашей эры[24]. Однако при исследованиях все семь образцов из ямной культуры принадлежали к R1b1a1a2-M269, а R1a1a не была обнаружена в образцах, отнесённых к ямной культуре[24]. Это поставило вопрос — откуда возникла R1a1a в культуре боевых топоров, учитывая что ямная культура не являлась носителем данной субклады.
Семёнов и Булат спорят о происхождении R1a1a в культуре боевых топоров, отмечая, что несколько исследований указывают на присутствие R1a1a в генетике данной культуры[30][31][32][33].
Исследования от 2015 года обнаружили, что часть предков людей ямной культуры ведёт свою генетику с Ближнего Востока, и что неолитический период в ямной культуре, связанный с освоением соответствующих навыков, начался на Балканах. При этом исследователь Хаак, поддерживающий эту версию, отмечает, что генетически люди ямной культуры не происходят от земледельцев Ближнего Востока, а ведут корни от людей с Ближнего Востока.
Рёссен — культура среднего неолита, существовавшая между 4600—4300 годами до н. э., была распространена в Германии и предшествовала культуре боевых топоров, но не напрямую, а через последовательно сменяющиеся культуры — культуру воронковидных кубков и культуру шаровидных амфор. Рёссен может иметь связь с древним подклассом R1a, а именно R1a1a1a1-L664 — согласно ДНК-исследованиям, она сформировалась к 2700 году до нашей эры.
Некоторые исследователи полагают, что R1a попала на Балканы через Анатолию, а уже оттуда распространялась дальше на северо-запад, до культуры рёссен, затем на восток от трипольской культуры до ямной культуры и афанасьевской культуры. При этом исследователи признают — R1a отсутствует в промежуточных культурах между Ближним Востоком, Анатолией и Балканами. Аско Парпола, исследуя эту проблему, предположил, что трипольская культура соотносится с поздним вариантом праиндоевропейского этноса. Он отмечает, что в трипольской культуре, возможно, изобретено колесо, а также что, вероятно, люди трипольской культуры была захвачены или ассимилированы носителями праиндоевропейского языка в 4-м тысячелетии до н. э., после чего культура распространилась до Понтийской степи к 3400 годам до нашей эры и в результате развития перетекла в ямную культуре[34].
Выделение греческой линии
На данный момент мало информации о прибытии протогреческих носителей языка из степей. Микенская культура возникла около 1650 лет до нашей эры и явно связана с прибытием переселенцев от степных культур. Микенский язык тесно связан с протоиндо-иранскими языками и разделение их на отдельные языки произошло относительно поздно, между 2500—2000 годами до нашей эры. Своеобразные микенские колесницы, наконечники копий, кинжалы из бронзы и другие изделия из неё показывают большое сходство с необычным археологическим памятником, называемым в научной литературе Сейминско-турбинский феномен (с 1900 по 1600 лет до нашей эры) — в нём известна Володарская стоянка и в том числе в ней — Сейминский могильник.
Изделия и предметы из этого археологического памятника, расположенного рядом с городом Володарск (Нижегородская область), напрямую коррелируют с находками в ареале Микенской цивилизации — это южная часть Балканского полуострова. Также схожие признаки культуры, оставившей Сейминский могильник, найдены в Монголии, что говорит о высокой мобильности кочевых степных народов. Данные факты указывают на возможную миграцию культур из лесостепной зоны центральной части Восточно-Европейской равнины на Балканы, смешивание их с местным населением и затем основание ими Микенской цивилизации, положившей начало Греческой культурной общности.
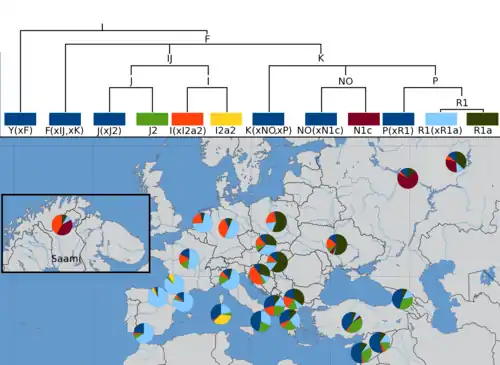
Закавказье и западно-европейское происхождение, возможное влияние Индской цивилизации
Часть генетической составляющей в жителях Южной Азии происходит от западно-европейского населения, и некоторые исследователи подразумевают, что субклад R1a1a1b2-Z93, возможно, прибыл в Индию через Иран[35] и расширился там во время Индской — Хараппской цивилизации[36][37].
В 2015 году предположено[38], что корни -R1a1a1b2-Z93 лежат в Западной Азии и её распространение вместе с L342.2 проходило в Юго-Восточном направлении от Закавказья до Южной Азии"[35], мотивируя это археологическими данными о восточной популяции Западной Азии в IV тысячелетии до нашей эры, кульминацией которого стали так называемые миграции куро-араксской культуры в период после Урука IV (период истории Месопотамии между 4000 — 3100 лет до нашей эры)[35]. Тем не менее Лазаридис отметил, что в куро-аракский образец I1635 была привнесена Y-гаплогруппа R1b1-M415 (xM269)[39] также называемая R1b1a1b-CTS3187[40].
Согласно результатам исследования 2014—2015 годов[41], диверсификация-отделение R1a1a1b2-Z93 и процессы урбанизации в долине Инда произошли 5600 лет назад, и географическое распределение R1a1a1b2a1~-M780 (нижестоящего к R1a1a1b2a-Z94)[42] может отражать это. Согласно ДНК, прослеживаемого через генеалогическое древо, R1a1a1b2a1~-M780 сформировалась 4700 лет назад. Этот процесс по возрасту совпадает с движением на восток между 2800 и 2600 годами до н. э. ямной культуры в область полтавкинской культуры, предшественницы синташтинской культуры, из которой возникли индоиранцы. Субклад R1a1a1b2a1~-M780 сконцентрирован в долине Ганг, центре классической ведийской цивилизации[5].
В 2016 году отмечено[43], что «произошло поразительное расширение» субклады R1a1a1b2-Z93 в промежуток между 4500—4000 л. н., что на несколько столетий предшествует угасанию, краху Индской цивилизации[37].
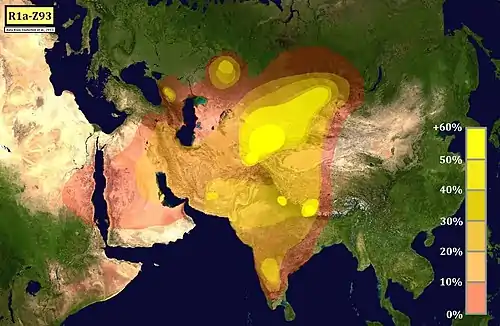
Теория южно-азиатского происхождения
В 2003 году высказано предположение о происхождении либо в Южной, либо в Западной Азии[44]. Это связано с фактом: гаплогруппа R1a, ранее уже увязываемая как маркер с предполагаемым индоарийским вторжением, была найдена в самой высокой частоте генома в Пенджабе и также в высокой частоте (26 %) — в племени чэнчу, одно из коренных индийских племён группы — адиваси. Это открытие вместе с более высоким составляющим маркера R1a — тандемное повторное разнообразие в Индии и Иране, по сравнению с Европой и Центральной Азией, предполагает, что источником этой гаплогруппы могут быть Южная и Западная Азии.
В исследовании 2009 года указывается в качестве возможной «прародины» гаплогруппы как Южная, так и Центральная Азия[45].
Южноазиатские популяции имеют наивысшее разброс микросателлитов в пределах R1a1a[45][45][46][47][48], и последующих, более поздних датировок на ближайшего общего предка. А R1a1a присутствует как среди высших каст (брахманов), так и низших каст, хотя присутствие значительно выше среди брахманов[48]. Исходя из этих данных, некоторые исследователи пришли к выводу, что R1a1a возникла в Южной Азии[49], исключая существенный генетический приток от индоевропейских мигрантов[49][50][51].
Однако это разнообразие и последующие более ранние датировки на выявление ближайшего общего предка также могут быть объяснены исторически большим числом населения, что увеличивает вероятность диверсификации и изменения микросателлитов[52][53][54]. R1a1 и R2, возможно, распространялись в Южную Индию из юго-западного региона Азии несколько раз[50].
В 2017 году отмечено[55], что R1a в Южной Азии наиболее вероятно распространилась от одного центрального пучка источников в Центральной Азии, по крайней мере — трёх. И, вероятно, эти генетические линии в дальнейшем развивались в границах Индийского субконтинента, что согласуется с несколькими волнами миграций[52].
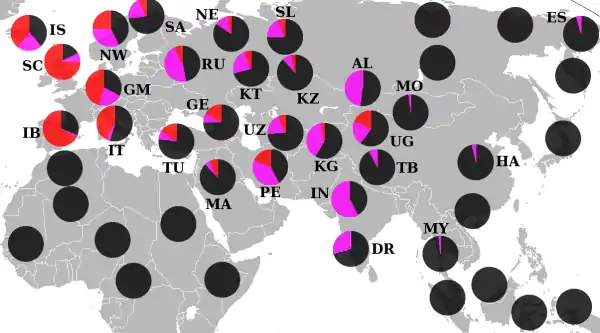
Филогенетика
Генеалогическое древо гаплогруппы R1a имеет три основных уровня ветвления с наибольшим количеством определённых субкладов в доминирующей и наиболее известной — R1a1a, она же R1a1.
Топология
Топология R1a выглядит следующим образом[56][57][58][59].
Согласно работе Tatiana Karafet et al. (2014), быстрый процесс диверсификации гаплогруппы K-M526, вероятно, произошел в Юго-Восточной Азии с последующими расширениями на западе предков гаплогрупп R и Q[59][60].
- P P295/PF5866/S8 (ранее известная как K2b2).
- R (R-M207)[5][61]
- R*
- R1 (R-M173)
- R1*[61]
- R1a (M420)[61] (Eastern Europe, Asia)[62]
- R1a*[5]
- R1a1[61] (M459/PF6235,[61] SRY1532.2/SRY10831.2[61])
- R1a1 (M459)[5][61]
- R1a1b (YP1272) https://web.archive.org/web/20150428004519/http://forum.molgen.org/index.php?topic=7835.0
- R1a1a (M17, M198)[61]
- R1a1a1 (M417, page7)[61]
- R1a1a1a (CTS7083/L664/S298)[61]
- R1a1a1b (S224/Z645, S441/Z647)[61]
- R1a1a1b1 (PF6217/S339/Z283)[61]
- R1a1a1b2 (F992/S202/Z93)[61] [R1a1a2*] (Z93, M746)(Asia)[63]
- [R1a1a1c] (M64.2, M87, M204)[64]
- [R1a1a1d] (P98)[64]
- [R1a1a1d2a][66]
- [R1a1a1e] (PK5)[64]
- R1a1a1 (M417, page7)[61]
- R1b (M343) (Western Europe)
- R2
Палеогенетика
Самая древняя известная R1a (субклад R1a1b~-YP1272>YP1301) определена у представителя культуры Веретье из могильника Песчаница 1 PES001 (10785—10626 лет до н. э.) на озере Лача (Каргопольский район Архангельской области)[68].
Гаплогруппа R1a обнаружена в останках следующих культур и народов: культура боевых топоров[69][70][71], Культура полей погребальных урн[72], синташтинская культура[73], андроновская культура[74], пазырыкская культура[75], тагарская культура[74], таштыкская культура[74], у жителей древнего Танаиса (город)[76], в таримских мумиях[77], у аристократов Хунну[78], в двух древних останках хазар.
Субклад R1a1a1b1-Z283 обнаружен у представителя культуры боевых топоров, в захоронении Бергсгравен (Bergsgraven), в Швеции, номер теста ber1[79].
Линия R1a1a1b2a1~-Y3, предковая для субклада R1a1a1b2a1a-L657, в настоящее время распространена особенно в Индии в высшей варне — у браминов. Она найдена в Поднепровье, у реки Оскол, в Харьковской области (Alexandria, Ukraine_MBA, Украина). Датировка захоронения 2134—1950 гг. до н. э.[80], номер образца палео-ДНК — I6561[81].
У темнокожих представителей фатьяновской культуры определена Y-хромосомная гаплогруппа R1a1a1-M417, при этом у половины образцов удалось уточнить субклад R1a1a1b2-Z93[68].
Гаплогруппа R1a, обнаружена у населения срубной и алакульской археологических культур, субклады R1a-Z280 и R1a-Z93:
Номер теста: kzb005, захоронение Казбурун 1, срубно-алакульская культура, датировка 1880—1690 гг. до н. э., R1a-M459, R1a-Z283, Z280[82],
Номер теста: kzb008, захоронение Казбурун 1, срубно-алакульская культура, датировка 1880 −1690 гг. до н. э., R1a-M459, R1a-Z283, Z280?[82],
Номер теста: kzb002, захоронение Казбурун 1, срубно-алакульская культура, датировка 1875—1665 гг. до н. э. R1a-M459, R1a-Z93, YP5585[82],
Номер теста: kzb003, захоронение Казбурун 1, срубно-алакульская культура, датировка 1765—1630 гг. до н. э. R1a-M459, R1a-Z93, Z94[82],
Номер теста: mur002, захоронение Мурадым 8, срубно-алакульская культура, R1a-M459[82],
Номер теста: mur003, захоронение Мурадым 8, срубно-алакульская культура, датировка 1880—1685 гг. до н. э. R1a-M459, R1a-Z93[82],
Номер теста: kzb007, захоронение Казбурун 1, срубно-алакульская культура, датировка 1755—1630 гг. до н. э. R1a-M459, R1a-Z93, Z94, Z2124, Z2125, Z2123, Y934, BY30762?[82]
у киммерийцев, субклад R1a-Z93:
Номер теста: cim357, захоронение Глиное Сад, киммерийцы, датировка 914—805 гг. до н. э., R1a-M459, R1a-Z93[82],
у скифов, субклад R1a-Z93:
Номер теста: scy193*, захоронение Глиное, скифы, R1a-M459, R1a-Z93, Z94, Z2124, Z2125, Z2123, Y934, BY30762, BY30764?[82]
у сармат, субклад R1a1a1b2-Z93:
Номер теста: chy002, захоронение Чёрный Яр, сарматы, датировка 65 — 220 гг. н. э. R1a-M459, R1a-Z93, Z94, Z2124, Z2122, Y57, Y52[82]
Номер теста: tem002, захоронение Темясово, сарматы, датировка 125—240 гг. н. э. R1a-M459, R1a-Z93, Z94, Z2124, Z2125, S23592, YP1558, Y73758, YP5844, FGC48758[82]
Номер теста: tem003 захоронение Темясово, сарматы, датировка 130—320 гг. н. э. R1a-M459, R1a-Z93, Z94, Z2124, Z2125, Z2123, YP3920[82]
Скелеты отца и двух его сыновей с археологического объекта, обнаруженного в 2005 году недалеко от Ойлау (Саксония-Анхальт, Германия) и датируемых примерно 2600 годом до н. э., показали положительный результат для маркера Y-SNP SRY10831.2. Таким образом предковая клада присутствовала в Европе по меньшей мере 4600 лет назад и была связана с одним из археологических памятников Культуры боевых топоров[83]. Номер Ysearch для останков Eulau — 2C46S. Таким образом, предковая клада присутствовала в Европе по меньшей мере 4600 лет назад, в связи с широко распространенной культуры шнуровой керамики.
Так же гаплогруппа R1a была выявлена в нескольких случаях останков населения межовской[84], хвалынской, срубной, потаповской, полтавкинской[85], тшинецкой[86]. археологических культур, на стоянке Афонтова гора (бронза)[84], у скифов[87][88][89], у населения срубной культурно-исторической общности[89], среди аланских захоронений (субклад Z2124 — R1a1a1b2a2)[90], у населения салтово-маяцкой культуры (субклад Z2124 — R1a1a1b2a2)[90], у обитателя Южного Оленьего острова на Онежском озере (R1a1-SRY10831.2), у одного представителя культуры шнуровой керамики (боевых топоров) из Эсперштедта и у двух из саксонской пещеры Лихтенштейн, у одного представителя культуры полей погребальных урн из Хальберштадта и у двух из Эйлау[91][92]. Гаплогруппа R1a1 прослежена у обитателей верховий Западной Двины с 5120±120 лет назад (Сертея VIII) до VIII—V веков до нашей эры (городище Анашкино)[93].
Ближний Восток и Передняя Азия
Распространение в странах Ближнего Востока и Передней Азии:
Центральная Азия
У киргизов R1а1а — 63 % [94]. В Афганистане R1a1a встречается у 51 % пуштунов и 30 % таджиков, реже — среди хазарейцев (7 %). У казахов родов — ошакты 26,7 %, суан — 31,7 % и кожа 35 %. У узбеков (18 %)[95].
Южная Азия
В Южной Азии R1a1a часто наблюдается с высокой частотой в ряде демографических групп.
В Индии высокие частоты этой гаплогруппы наблюдаются у западных бенгальских брахманов (72 %) на восток, лоханов (60 %) на западе, кхатри (67 %) на севере и айенгары (31 %) на юге. Она также была найдена у нескольких южно-индийских дравидийских племён адиваси, включая ченчу (26 %), валмики из Андхра-Прадеша и калларов в Тамилнаде, что говорит о том, что R1a1a широко распространена у племенных южных индийцев.
Кроме того, исследования показывают высокие проценты в региональных группах, таких как Манипури (50 %), до крайнего северо-востока и среди пенджабцев (47 %) до крайнего северо-запада.
В Пакистане он встречается у 71 % среди племени Моханны в провинции Синд на юге и 46 % среди балтийцев Гилгит-Балтистан на севере. Среди сингальцев Шри-Ланки 23 % были обнаружены как R1a1a (R-SRY1532). Индусы района Читван в регионе Тераи Непал показывают его на уровне 69 %[49][50][51][96].
Восточная Азия
Частота R1a1a сравнительно низкая среди некоторых тюркоязычных групп, включая турок, азербайджанцев, казахов и якутов.
Но выше (от 19 до 28 %) в некоторых тюркских или монгольских группах северо-западного Китая — баоань, дунсян, саларов, уйгуров.
В Восточной Сибири R1a1a встречается среди некоторых коренных этнических групп, включая камчатцев и чукчей, и достигает у ительменов 22 %[97].
Западная Азия
R1a1a был обнаружен в различных формах, в большинстве районов Западной Азии, в самых разных концентрациях. От почти не присутствующих в таких районах, как Иордания, до гораздо более высоких уровней в некоторых частях Кувейта, Турции и Ирана. Племена бедуинов Шаммар в Кувейте показывают самую высокую частоту на Ближнем Востоке — до 43 %[98].
В 2001 году отмечено[99], что в западной части страны у иранцев низкие уровни R1a1a, в то время как мужчины в восточной части Ирана имеют до 35 % R1a1a. В 2004 году R1a1a найдена примерно у 20 % иранских мужчин из городов Тегеран и Исфахан. В исследовании Ирана 2006 года отмечены гораздо более высокие частоты на юге, чем на севере.
Новое исследование показало 20,3 % R-M17 * среди курдских образцов, которые были взяты в провинции Курдистан в западном Иране, 9,7 % среди Мазандараниса в Северном Иране в провинции Мазандаран, 9,4 % среди гилаков в провинции Гилан, 12,8 % Персидский и 17,6 % среди зороастрийцев в Йезде, 18,2 % среди персов в Исфахане, 20,3 % среди персов в Хорасане, 16,7 % афро-иранцев, 18,4 % Кешми «Гешми», 21,4 % среди персидских говорящих людей Бандари в Хормозгане и 25 % среди Жители Балоха в Систане и провинции Балухстан.
Гаплогруппа R1a1a была найдена в повышенных уровнях среди выборки израильского населения, которые сами себя обозначили как левиты и ашкеназские евреи (левиты составляют около 4 % евреев). В исследовании Бехара установлено, что R1a1a является доминирующей гаплогруппой у ашкеназских левитов (52 %), хотя она редко встречается у ашкеназов-коэнов (1,3 %).
С другой стороны, к северу от этих ближневосточных регионов, уровни R1a1a начинают расти на Кавказе неравномерно. Несколько изученных популяций не показали никаких признаков R1a1a, в то время как самые высокие уровни, обнаруженные до сих пор в регионе, среди карачаевцев ок. 40 % и среди балкарцев ок. 35 % представлены доминирующей гаплогруппой R1a1a[8][97][100][101][102][103].
Известные представители гаплогруппы R1a
См. также
Примечания
- YFull Tree
- Maanasa Raghavan, Pontus Skoglund, Kelly E. Graf, Mait Metspalu, Anders Albrechtsen, Ida Moltke, Simon Rasmussen, Thomas W. Stafford Jr, Ludovic Orlando, Ene Metspalu, Monika Karmin, Kristiina Tambets, Siiri Rootsi, Reedik Mägi, Paula F. Campos, Elena Balanovska, Oleg Balanovsky, Elza Khusnutdinova, Sergey Litvinov, Ludmila P. Osipova, Sardana A. Fedorova, Mikhail I. Voevoda, Michael DeGiorgio, Thomas Sicheritz-Ponten, Søren Brunak et al. «Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans», 2014
- Sharma, S; Rai, E; Sharma, P; Jena, M; Singh, S; Darvishi, K; Bhat, AK; Bhanwer, AJ. et al. (2009). «The Indian origin of paternal haplogroup R1a1(*)substantiates the autochthonous origin of Brahmins and the caste system». Journal of Human Genetics. 54 (1): 47-55
- Underhill, Peter A. (2014), «The phylogenetic and geographic structure of Y-chromosome haplogroup R1a» (PDF), European Journal of Human Genetics, 23 (1): 124—131, doi:10.1038/ejhg.2014.50, ISSN 1018-4813, PMC 4266736 ,PMID 24667786
- R1a YTree
- .
- Mirabal, Sheyla; Regueiro, M; Cadenas, AM; Cavalli-Sforza, LL; Underhill, PA; Verbenko, DA; Limborska, SA; Herrera, RJ; et al. (2009). «Y-Chromosome distribution within the geo-linguistic landscape of northwestern Russia». European Journal of Human Genetics. 17 (10): 1260—1273. doi:10.1038/ejhg.2009.6
- Underhill, Peter A; Myres, Natalie M; Rootsi, Siiri; Metspalu, Mait; Zhivotovsky, Lev A; King, Roy J; Lin, Alice A; Chow, Cheryl-Emiliane T; et al. (2009). «Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a». European Journal of Human Genetics.
- Zerjal T. et al. The use of Y-chromosomal DNA variation to investigate population history: recent male spread in Asia and Europe // in S.S. Papiha, R. Deka and R. Chakraborty (eds.), Genomic diversity: applications in human population genetics (1999), pp. 91-101.
- Poznik et al. Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences, 2016
- Narasimhan, Vagheesh M.; Anthony, David; Mallory, James; Reich, David (2018), The Genomic Formation of South and Central Asia, bioRxiv 10.1101/292581, doi:10.1101/292581
- Semino, O.; Passarino, G; Oefner, PJ; Lin, AA; Arbuzova, S; Beckman, LE; De Benedictis, G; Francalacci, P; et al. (2000).
- https://web.archive.org/web/20031125151213/http://hpgl.stanford.edu/publications/Science_2000_v290_p1155.pdf
- Wells, R.S. (2001), «The Eurasian Heartland: A continental perspective on Y-chromosome diversity», Proceedings of the National Academy of Sciences of the USA, 98 (18): 10244-10249, doi:10.1073/pnas.171305098, PMC 56946
- Iran: Tricontinental Nexus for Y-Chromosome Driven Migration — Abstract — Human Heredity 2006, Vol. 61, No. 3 — Karger Publishers
- Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a
- Ornella Semino, Giuseppe Passarino, Peter J. Oefner, Alice A. Lin, Svetlana Arbuzova, Lars E. Beckman, Giovanna De Benedictis, Paolo Francalacci, Anastasia Kouvatsi, Svetlana Limborska, Mladen Marciki, Anna Mika, Barbara Mika, Dragan Primorac, A. Silvana Santachiara-Benerecetti, L. Luca Cavalli-Sforza, Peter A. Underhill, The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective, Science, vol. 290 (10 November 2000), pp. 1155—1159.
- The Eurasian Heartland: A continental perspective on Y-chromosome diversity // PNAS
- Pamjav 2012.
- Massive migration from the steppe is a source for Indo-European languages in Europe // bioRxiv
- Population genomics of Bronze Age Eurasia | Nature
- Eight thousand years of natural selection in Europe | bioRxiv
- The Horse The Wheel And Language. How Bronze-Age Riders From the Eurasian Steppes Shaped The Modern World, Princeton University Press
- Ringe, Don (2015), «The Indo-European Homeland from Linguistic and Archaeological Perspectives», Annual Review of Linguistics
- Дискуссия о происхождении индоевропейских языков из миграции ямной культуры на запад.
- Ямная культура.
- Chuan-Chao Wang et al. The genetic prehistory of the Greater Caucasus, 2018
- Chuan-Chao Wang et al. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions, 04 February 2019
- Трипольская культура.
- Haak W. et al. Massive migration from the steppe is a source for Indo-European languages in Europe.
- Mathieson I et al. Eight thousand years of natural selection in Europe.
- Chekunova Е. М., Yartseva N.V., Chekunov М. К., Mazurkevich А.N. The First Results of the Genotyping of the Aboriginals and Human Bone Remains of the Archeological Memorials of the Upper Podvin’e. // Archeology of the lake settlements of IV—II Thousands BC: The chronology of cultures and natural environment and climatic rhythms. Proceedings of the International Conference, Devoted to the 50-year Research of the Pile Settlements on the North-West of Russia. St. Petersburg, 13-15 November 2014.
- Eppie R. Jones et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians. Nature Communications.
- The Roots of Hinduism. The early Aryans and the Indus Civilisation, Oxford University Press
- Mascarenhas 2015, p. 9
- Underhill 2014.
- Pozink 2016, p. 5.
- Mascarenhas et al. 2015
- Eurogenes Blog: Big deal of 2016: the territory of present-day Iran cannot be the Indo-European homeland
- Arame’s english blog: November 2016
- Underhill et al., 2014/2015
- Dienekes’ Anthropology Blog: Major new article on the deep origins of Y-haplogroup R1a (Underhill et al. 2014)
- Poznik et al. (2016)
- Kivisild 2003
- Y-Chromosome distribution within the geo-linguistic landscape of northwestern Russia
- Sengupta 2006
- A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios | PNAS
- Thangaraj, Kumarasamy; Naidu, B. Prathap; Crivellaro, Federica; Tamang, Rakesh; Upadhyay, Shashank; Sharma, Varun Kumar; Reddy, Alla G.; Walimbe, S. R.; Chaubey, Gyaneshwer; Kivisild, T; Singh, L; et al. (2010). Cordaux, Richard, ed. «The Influence of Natural Barriers in Shaping the Genetic Structure of Maharashtra Populations»
- Sahoo, S; Singh, A; Himabindu, G; Banerjee, J; Sitalaximi, T; Gaikwad, S; Trivedi, R; Endicott, P; et al. (2006). «A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios»
- Sengupta, S; Zhivotovsky, LA; King, R; Mehdi, SQ; Edmonds, CA; Chow, CE; Lin, AA; Mitra, M; et al. (2006). «Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists»
- Underhill, Peter A; Myres, Natalie M; Rootsi, Siiri; Metspalu, Mait; Zhivotovsky, Lev A; King, Roy J; Lin, Alice A; Chow, Cheryl-Emiliane T; et al. (2009). «Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a»
- Silva, Marina; et al. (2017), «A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals», BMC Evolutionary Biology,
- A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals | BMC Ecology and Evolution | Full Text
- How genetics is settling the Aryan migration debate — The Hindu
- Silva et al. (2017)
- ISOGG 2017 Y-DNA Haplogroup R
- Underhill, Peter A. (2015), «The phylogenetic and geographic structure of Y-chromosome haplogroup R1a», European Journal
- Haplogroup R (Y-DNA) — SNPedia
- Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia | European Journal of Human Genetics
- Karafet, Tatiana; Mendez, Fernando; Sudoyo, Herawati (2014). «Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia». Nature. 23
- ISOGG 2016 Y-DNA Haplogroup R
- Underhill, 2015.
- Underhill, 2015, p. 125.
- Haplogroup R (Y-DNA) - SNPedia. www.snpedia.com. Дата обращения: 20 декабря 2019.
- [www.familytreedna.com/groups/r-1a/about/results About Us]. Family Tree DNA. Дата обращения: 20 декабря 2019.
- eurogenes.blogspot, R1a in Yamnaya
- Eurogenes Blog: R1a in Yamnaya
- Lehti Saag, Sergey V. Vasilyev, Svetlana V. Oshibkina et al. Genetic ancestry changes in Stone to Bronze Age transition in the East European plain (Table 1), July 03, 2020 (bioRxiv)
- Haak, Wolfgang; Brandt, Guido; Jong, Hylke N. de; Meyer, Christian; Ganslmeier, Robert; Heyd, Volker; Hawkesworth, Chris; Pike, Alistair W. G.; Meller, Harald; Alt, Kurt W. (25 November 2008). «Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age». PNAS. 105 (47): 18226-18231. doi:10.1073/pnas.0807592105. PMC 2587582 . PMID 19015520. Retrieved 15 June 2016 — via www.pnas.org.
- Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age | PNAS
- Ancient DNA reveals key stages in the formation of Central European mitochondrial genetic diversity
- https://web.archive.org/web/20110814164431/http://dirkschweitzer.net/LichtensteinCaveAnalysis0804DS.pdf
- Allentoft; et al. (2015), «Population genomics of Bronze Age Eurasia», Nature, 522: 167—172,
- Keyser, Christine; Bouakaze, Caroline; Crubézy, Eric; Nikolaev, Valery G.; Montagnon, Daniel; Reis, Tatiana; Ludes, Bertrand (2009). «Ancient DNA provides new insights into the history of south Siberian Kurgan people». Human Genetics.
- Ricaut, F.; et al. (2004). «Genetic Analysis of a Scytho-Siberian Skeleton and Its Implications for Ancient Central Asian Migrations». Human Biology. 76: 1.
- Корниенко И. В., Водолажский Д. И. Использование нерекомбинантных маркеров Y-хромосомы в исследованиях древних популяций (на примере поселения Танаис) // Материалы Донских антропологических чтений. Ростов-на-Дону, Ростовский научно-исследовательский онкологический институт, Ростов-на-Дону, 2013.
- https://web.archive.org/web/20110427172440/http://www.biomedcentral.com/content/pdf/1741-7007-8-15.pdf
- Kim, Kijeong; Brenner, Charles H.; Mair, Victor H.; Lee, Kwang-Ho; Kim, Jae-Hyun; Gelegdorj, Eregzen; Batbold, Natsag; Song, Yi-Chung; Yun, Hyeung-Won; Chang, Eun-Jeong; Lkhagvasuren, Gavaachimed; Bazarragchaa, Munkhtsetseg; Park, Ae-Ja; Lim, Inja; Hong, Yun-Pyo; Kim, Wonyong; Chung, Sang-In; Kim, Dae-Jin; Chung, Yoon-Hee; Kim, Sung-Su; Lee, Won-Bok; Kim, Kyung-Yong (2010). «A western Eurasian male is found in 2000-year-old elite Xiongnu cemetery in Northeast Mongolia». American Journal of Physical Anthropology.
- PgmNr 2363: Multiple waves of peopling northeast Europe: An ancient DNA study through Mesolithic to the Middle Ages. Authors: M. Metspalu,; L. Saag 1,2; K. Tambets. При более детальном исследовании тест ber1 — относится к линии R1a-Y2395 * (YP694, YP3896). Одиночные производные чтения для R1a-Z662 (уровень R1a1a1b1-Z283) и R1a1a1b1a-Z282, три SNP для Y2395, нет чтения на SNP Z289. Отрицательно для всех SNP из трех известных нисходящих ветвей.
- Allen Ancient DNA Resource (AADR): Downloadable genotypes of present-day and ancient DNA data. V50.0: New data release: Oct 10 2021
- Iain Mathieson et al. The Genomic History Of Southeastern Europe, 2017
- Summary sequencing statistics for mitochondrial variants for individuals sequenced in this study. (*) Individuals previously used in a study focusing on mitochondrial genomes (Juras et al.2017) and are thus reported elsewhere.
- Haak 2008.
- Morten E. Allentoft et al. «Population genomics of Bronze Age Eurasia»
- Iain Mathieson et al. Eight thousand years of natural selection in Europe, 2015
- Fernandes D. M. et al. A genomic Neolithic time transect of hunter-farmer admixture in central Poland, 2018
- Ricaut F. et al. 2004. Genetic Analysis of a Scytho-Siberian Skeleton and Its Implications for Ancient Central Asian Migrations. Human Biology. 76 (1)
- F.X. et al. Genetic analysis and ethnic affinities from two Scytho-Siberian skeletons, 2004.
- Samples, Mathieson - Google Таблицы. docs.google.com. Дата обращения: 12 октября 2015.
- Афанасьев Г. Е. и др. Хазарские конфедераты в бассейне Дона // Тезисы докладов на Всероссийской научной конференции «Естественнонаучные методы исследования и парадигма современной археологии». М.: ИА РАН. 2015, С.9.]
- Massive migration from the steppe is a source for Indo-European languages in Europe
- Mitochondrial and Y-chromosome haplogroups extracted from historic and prehistoric human remains in Europe and related remains in Asia, arranged chronologically
- Чекунова Е. М. Ярцева, Н. В., Чекунов М. К., Мазуркевич А. Н. «Первые результаты генотипирования коренных жителей и человеческих костных останков из археологических памятников Верхнего Подвинья». С. 287—294. // Археология озёрных поселений IV—II тыс. до н. э.: хронология культур и природно-климатические ритмы. — СПб.: ООО «Периферия», 2014.
- Shou et. al. 2010, Y-chromosome distributions among populations in Northwest China identify significant contribution from Central Asian pastoralists and lesser influence of western Eurasians
- Haber 2012
- http://evolutsioon.ut.ee/publications/Kivisild2003a.pdf
- Wells, R.S. (2001), «The Eurasian Heartland: A continental perspective on Y-chromosome diversity», Proceedings of the National Academy of Sciences of the USA
- Nasidze 2005.
- Wells 2001
- Nasidze 2004.
- Mohammad 2009.
- Grugni 2012.
- Behar 2003.
- Peter L. Nagy and etc. Determination of the phylogenetic origins of the Árpád Dynasty based on Y chromosome sequencing of Béla the Third
- Judit Olasz. DNA profiling of Hungarian King Béla III and other skeletal remains originating from the Royal Basilica of Székesfehérvár, 2018
Литература
- Underhill, Peter A. (2015), The phylogenetic and geographic structure of Y-chromosome haplogroup R1a, European Journal of Human Genetics Т. 23: 124–131, PMID 24667786, DOI 10.1038/ejhg.2014.50
Ссылки
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 GHIJK | |||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||