Гаплогруппа HV (мтДНК)
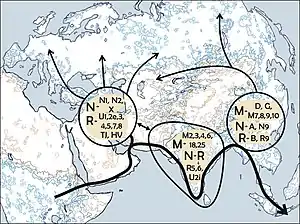
В популяционной генетике человека гаплогруппой HV называют одну из гаплогрупп, выявленных при анализе последовательности митохондриальной ДНК (mtDNA). Гаплогруппа pre-HV или RO, распространённая на Ближнем Востоке, особенно в Аравии, а также в Эфиопии и Сомали, происходит от гаплогруппы R, которая, в свою очередь, отделилась от гаплогруппы N около 40 тыс. лет назад.
| Гаплогруппа HV | |
|---|---|
 | |
| Тип | мтДНК |
| Время появления | 20—30 тыс. лет назад |
| Место появления | западная Евразия |
| Предковая группа | R0 |
| Субклады | HV0, HV1, HV2, HV4, HV5, HV6, HV7, HV8, HV9, H |
| Мутации-маркеры | C14766C[1] |
Эта гаплогруппа широко распространена в Западной Европе, её носители пришли туда через территорию Кавказа во время периода после последнего ледникового максимума и в неолите[2].
Параллельные филогеографии редких, но широко распространенных субкладов HV*(xH, V) обнаруживают связь между Апеннинским полуостровом и Южным Кавказом, обусловленную по меньшей мере двумя (после последнего ледникового периода и в эпоху неолита) волнами миграции. HV1b-152 поддерживает северное месопотамское происхождение для линий ашкенази HV1b2. В соответствии с древними находками ДНК, филогенетический анализ HV12 и HV14, двух исключительно азиатских субкладов HV*(xH,V), указывает на миграцию линий, происходящих из Ирана в Южную Азию до и в течение периода неолита[3].
Палеогенетика
Анализ ДНК кроманьонца Paglicci-25, жившего в пещере Пальиччи в Южной Италии около 24 тыс. лет назад, показал, что он принадлежал к гаплогруппе HV или её предку — preHV[4][5].
HV обнаружена у представителя группы Зальцмюнде (en:Salzmünde group) культуры воронковидных кубков[6], представителей андроновской культуры, культуры боевых топоров (шнуровой керамики), унетицкой культуры[7].
HV0 определена у представителя старчево-кришской культуры[8], гальштатской культуры[9].
HV1a'b'c определена у энеолитического (4500—3900/3800 гг. до н. э.) образца I1165 из израильской пещеры Пкиин (Peqi’in Cave)[10].
HV6’17 определена у представителя баальбергской культуры из немецкого Кведлинбурга (№ I0559), жившего 3645—3537 лет до н. э.[11], pre-HV и HV — у представителей трипольской культуры[12].
HV1a2 определили у представителя бабинской культуры (многоваликовой керамики) L112, жившего 3230±70 лет назад[13][14].
HV0a определили у неолитического образца NOE002 (5352±32 лет до н. в.) из некрополя Necropoli di Noeddale (коммуна Осси, SS) на Сардинии[15].
HV, HV9 и HV15 определили у представителей культуры колоколовидных кубков из Богемии (2400 лет до н. э.)[16].
HV0a определили у представителя унетицкой культуры I15642 (Czech_EBA_Unetice, 3850 л. н.) из Чехии[7]
HV0a1a определили у образца I18719 эпохи бронзы (Croatia_MBA_LBA, 3200 л. н.) из Хорватии[7]
HV0a определили у образца BRC024 (Broion, not dated, Италия)[17].
HV определена у представителя кобанской культуры из могильника Заюково-3, расположенного близ села Заюково в Баксанском районе Кабардино-Балкарии (VIII—VII века до н. э.)[18].
HV1a2a, HV1a'b'c, HV1b2, HV21 определены у мумий из Абусира[19]. HV обнаружена у древнеегипетской мумии YM S7 из художественного музея Тартуского университета, датируемой второй половиной 1 тыс. до нашей эры[20].
HV0j определили у пунического образца ORC002 (2255±22 л. н.) из S’Orcu ‘e Tueri (коммуна Пердасдефогу, NUO) на Сардинии[15]
HV0a определили у представителя латенской культуры I11712 (Slovakia_LIA_LaTene, 2046 л. н.) из Словакии[7].
HV0 и HV0a1 определены у двух обитателей Гнёздова X—XI веков[21].
HV0a и HV1b выявлены у крестоносцев из Сидона, живших примерно в XI—XIII веках[22].
HV0 определили у останков великой княгини Софьи Палеолог, дочери Ивана III великой княжны Евдокии Ивановны и племянницы Василия III Анастасии Петровны[23].
Примечания
- van Oven, Mannis; Manfred Kayser. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation (англ.) // Human Mutation : journal. — 2008. — 13 October (vol. 30, no. 2). — P. E386—E394. Архивировано 4 декабря 2012 года.
- Коробов Д. и др. Новые данные об этногенезе северокавказских алан: археология, антропология, палеогенетика (In Russian), April 2020
- Michel Shamoon-Pour, Mian Li, D. Andrew Merriwether . Rare human mitochondrial HV lineages spread from the Near East and Caucasus during post-LGM and Neolithic expansions, 2019
- Ancient DNA
- Evidence for a genetic discontinuity between Neandertals and 24,000-year-old anatomically modern Europeans
- Brandt G. et al. (2013) Ancient DNA Reveals Key Stages in the Formation of Central European Mitochondrial Genetic Diversity, Science, vol. 342, no. 6155 (2013), pp. 257-261.
- Nick Patterson et al. Large-scale migration into Britain during the Middle to Late Bronze Age // Nature, 22 December 2021
- Szécsényi-Nagy (2015), Molecular genetic investigation of the Neolithic population history in the western Carpathian Basin Архивная копия от 21 июля 2015 на Wayback Machine
- Peter de Barros Damgaard et al. 137 ancient human genomes from across the Eurasian steppes, 2018
- Éadaoin Harney et al. Ancient DNA from Chalcolithic Israel reveals the role of population mixture in cultural transformation, 2018
- Massive migration from the steppe is a source for Indo-European languages in Europe
- Nikitin A. G. et al. (2010) Comprehensive site chronology and ancient Mitochondrial DNA analysis from Verteba cave – a trypillian culture site of eneolithic Ukraine. http://www.iansa.eu/papers/IANSA-2010-01-02-nikitin.pdf
- Alexey G. Nikitin et al. Subdivisions of haplogroups U and C encompass mitochondrial DNA lineages of Eneolithic–Early Bronze Age Kurgan populations of western North Pontic steppe, 2 February 2017
- Jeff Pashnick. Table 1: Specimen data for individuals in this study. Genetic Analysis of Ancient Human Remains from the Early Bronze Age Cultures of the North Pontic Steppe Region, 8-2014
- Joseph H. Marcus et al. Genetic history from the Middle Neolithic to present on the Mediterranean island of Sardinia, 24 February 2020
- Luka Papac et al. Dynamic changes in genomic and social structures in third millennium BCE central Europe // Science Advances. Vol. 7, Issue 35, 25 Aug 2021
- Tina Saupe et al. Ancient genomes reveal structural shifts after the arrival of Steppe-related ancestry in the Italian Peninsula, May 10, 2021 (Table 1. Archaeological information, genome coverage, genetic sex, mtDNA, and Y chromosome haplogroups of the individuals of this study)
- Eugenia Boulygina et al. Mitochondrial and Y-chromosome diversity of the prehistoric Koban culture of the North Caucasus, 2020
- Verena J. Schuenemann et al. Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods, 30 May 2017
- Ester Oras et al. Multidisciplinary investigation of two Egyptian child mummies curated at the University of Tartu Art Museum, Estonia (Late/Graeco-Roman Periods), 2020
- Ashot Margaryan et al. Population genomics of the Viking world, 2019
- Marc Haber et al. A Transient Pulse of Genetic Admixture from the Crusaders in the Near East Identified from Ancient Genome Sequences, 2019
- Корниенко, И. В. Молекулярно-генетическая идентификация безымянных захоронений первой половины XVI в. из некрополя Вознесенского собора Московского Кремля : [рус.] : ст. / И. В. Корниенко, Т. Д. Панова, Т. Г. Фалеева … [и др.] // Генетика. — 2022. — Т. 58, № 2. — С. 206-218. — doi:10.31857/S0016675822020072.
Ссылки
- Spread of Haplogroup HV, from National Geographic
- [www.familytreedna.com/public/india The India Genealogical DNA Project]
- The mtDNA Haplogroup H Project (Home to HVs)
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| CZ | D | E | G | Q | R | O | A | S | X | Y | N1 | N2 | ||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | UK | I | N1a | W | ||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры IWX | ||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||