Гаплогруппа D (мтДНК)
Гаплогруппа D — гаплогруппа митохондриальной ДНК человека.
| Гаплогруппа D | |
|---|---|
| Тип | мтДНК |
| Время появления | 60 тыс. лет назад |
| Место появления | Азия |
| Предковая группа | Гаплогруппа M |
И. А. Захаров-Гезехус с коллегами предложили дать гаплогруппе D имя Дарый (Daryy)[1].
Происхождение
Предполагается, что гаплогруппа D возникла в Азии около 60 тыс. лет тому назад. Она является потомком гаплогруппы M.
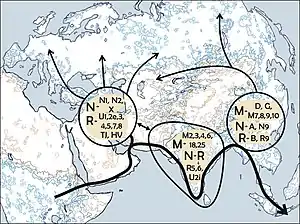
По оценке Н. В. Володько, митохондриальная гаплогруппа D4h3 возникла на Дальнем Востоке около 17 тыс. лет назад[2][3].
Филогенетическое дерево
- D
- D4
- D1
- D1a
- D1b
- D1c
- D1d
- D4a
- D4a1
- D4a1a
- D4a1b
- D4a1c
- D4a2
- D4a2a
- D4a3
- D4a3a
- D4a4
- D4a1
- D4b
- D4b1
- D3
- D4b1a
- D4b1a1
- D4b1a2
- D4b1a2a
- D4b1a2a1
- D4b1a2a1a
- D4b1a2a1
- D4b1a2a
- D4b1b
- D4b1b1
- D4b1b1a
- D4b1b2
- D4b2
- D4b2a
- D4b2a1
- D4b2a2
- D4b2a2a
- D4b2a2b
- D4b2b
- D4b2b1
- D4b2b1a
- D4b2b1b
- D4b2b2
- D4b2b3
- D4b2b1
- D4b2a
- D4b1
- D4c
- D4c1
- D4c1a
- D4c1a1
- D4c1b
- D4c1b1
- D4c1a
- D4c2
- D4c1
- D4d
- D4e
- D4e1
- D2
- D2a
- D2a1
- D2a1a
- D2a1b
- D2a1
- D2b
- D2a
- D4e1a
- D4e1a1
- D4e1a2
- D2
- D4e1
- D4e2
- D4e2a
- D4e2b
- D4e2c
- D4f
- D4g
- D4g1
- D4g1a
- D4g1b
- D4g1c
- D4g2
- D4g2a
- D4g2a1
- D4g2a
- D4g1
- D4h
- D4h1
- D4h1a
- D4h3
- D4h1
- D4i
- D4i1
- D4i2
- D4j
- D4k
- D4l
- D4l1
- D4l2
- D4l2a
- D4m
- D4m1
- D4m2
- D4n
- D4n1
- D4o
- D4o1
- D4o2
- D1
- D5’6
- D5
- D5a’b
- D5a
- D5a1
- D5a1a
- D5a2
- D5a2a
- D5a2a1
- D5a2a1a
- D5a2a1b
- D5a2a1
- D5a2a
- D5a1
- D5b
- D5b1
- D5b1a
- D5b1b
- D5b1b1
- D5b2
- D5b1
- D5a
- D5c
- D5d
- D5d1
- D5a’b
- D6
- D5
- D4
Распространение
Встречается в Северо-Восточной Азии (в том числе в Сибири). Также является одной из 5 мтДНК-гаплогрупп, обнаруженных у коренных народов Америки[4], наряду с такими, как A, B, C и X2a. D1 это базальная ветвь D4, которая широко распространена и разнообразна в Северной и Южной Америке. Субклады D4b1, D4e1 и D4h встречаются как в Азии, так и в Северной и Южной Америке. D2, встречается с высокой частотой в некоторых арктических и субарктических популяциях (особенно у алеутов) и является подклассом D4e1, параллельным D4e1a и D4e1c, поэтому его правильно следует называть D4e1b. D3 был найден главным образом в некоторых сибирских популяциях и у эскимосов Канады и Гренландии и является ветвью D4b1c.
Также гаплогруппа D довольно часто встречается в Центральной Азии[5], где она является второй по частоте кладой мтДНК (после митохондриальной гаплогруппы H). С низкой частотой гаплогруппа D встречается на северо-востоке Европы и в Юго-Западной Азии.
У монголов Китая митохондриальная гаплогруппа D достигает 27,07 %, у китайцев-хань — 23,53 %, у монголоязычных дауров — 10,53 %. Митохондриальная гаплогруппа D выявлена у 3 из 10, изученных в Китае носителей тунгусского языка[6].
Палеогенетика
- D1 определена у девочки-подростка из пещеры Хойо Негро (полуостров Юкатан), жившей 13—12 тыс. лет назад[7][8]
- D4h3a определена у мальчика Анзик-1 (en:Anzick-1), жившего 12,7 тыс. лет назад[9]
- D1 определили у 10 600-летней мумии Пещеры Духов (AHUR_2064), из штата Невада[10]
- В Южном конусе Южной Америки в раннем голоцене была распространена митохондриальная гаплогруппа D1j, в среднем голоцене — митохондриальная гаплогруппа D1g[11]
- D1j определили у образца LLP.S2.E1 (Laguna Chica, 10 223—9764 л. н.) из Аргентины[12]
- D4h3 определена у индивида Шука Каа (Shuká Káa) из пещеры «На коленях» (en:On Your Knees Cave) на острове принца Уэльского на юго-востоке штата Аляска, жившего 9730±60 — 9880±50 лет назад[13][14]
- D4h3a определили у образцов из бразильских Lapa do Santo возрастом ок. 9,5 тыс. л. н., Moraes возрастом ок. 5,8 тыс. л. н. и у образца из белизского Mayahak Cab Pek (ок. 9,3 тыс. л. н.)[15]
- D5a2a1h1 FY-HT-2 (9380±90 л. н.) из китайской пещеры Фуянь (en:Fuyan Cave)[16][17][18]
- D1 определили у образца ASO_B9_S14a (Arroyo Seco 2, 7832—7573 л. н.) из Аргентины[12]
- D4 определена у образца DevilsGate1 (5726—5622 лет до н. э.) из пещеры Чёртовы Ворота в Приморье[19]
- D4 определена у представителей культуры Бойсмана (~5000 лет до н. э.) в Приморье[20]
- D* и D1 выявлены у древних южноамериканцев (D* и D1 в Huaca Pucllana, D1 в Pasamayo, Pueblo Viejo (Перу) и в Llullaillaco (Аргентина)[21][22]
- D4b1a определили у представителей культуры Луншань (2275—1844 лет до н. э.)[23]
- D4j определили у образца KPT005 (4142—3980 л. н., ранний бронзовый век) из могильника Khaptsagai у деревни Люры (Иркутская область)[24]
- D2a1a определена у древних алеутов с Алеутских островов (от 2320—1900 до 500—140 л. н.)[25]
- D2a1 обнаружена у представителя палеоэскимосской культуры Саккак, жившего в Гренландии ок. 4 тыс. лет назад[26]
- D4b2b обнаружена у образцов JJD301.1 и JJD301.6 (3300 л. н.) из китайской пещеры Яндзиапо (Yangjiapo Cave)[16]
- D5a1a2ab определена у образца FY-HT-1 (2510±140 л. н.) из китайской пещеры Фуянь (en:Fuyan Cave)[16]
- Митохондриальная гаплогруппа D была обнаружена у мужчины-воина из погребения в Покровске (Якутия), жившего 2400—2200 лет назад[27]
- У представителей среднего периода Яёй определили митохондриальные гаплогруппы D4b2b1 (образцы из Doigahama DH-S01 (2,306—2,238 тыс. л. н.) и DH-A) и D4b2a1 (образец HN-SJ002, Hanaura, Япония)[28].
- У мужчины и женщины хунну из Duurlig Nars в Северо-Восточной Монголии, живших 2000 лет назад, определили митохондриальная гаплогруппа D4[29]
- D1g5 определили у образца LCH.E4.4 (Laguna Chica, 1627—1565 л. н.) из Аргентины[12]
- Урджарская принцесса из погребения сакского времени (III—IV век до н. э.) в Урджарском районе Восточно-Казахстанской области имела субкладу D4o[30][31]
- Исследование ДНК скелета гуннского периода из Музея естественной истории (г. Будапешт), датированного средней третью V века, показало, что он имел субкладу D4j12[31]
- D2a1 определили у образцов токаревской культуры (вторая половина 1 тыс. до н. э. — начало 1 тыс. н. э.) со стоянки Ольская на мысе Восточный (Ольский) (Магаданская область)[24]
- D5c1a определили у образца имперского периода Кофун JpIw31 (1303—1377 л. н.)[32].
- D4o2a определили у образца Yana _young (766 л. н.) с Янской стоянки (Якутия)[24]
- D4b1 определили у мужчины из погребения 688 гунно-сарматского времени памятника Сопка-2 в Барабинской лесостепи[33]
- D4j определена у образца Uyelgi21 из могильника IX века на озере Уелги (Челябинская область)[34]
- D4b1a2a1a обнаружена у образцов с кладбища в Нувуке (Барроу) на Аляске с калиброванными датами между 981 и 1885 годами нашей эры[35]
- D1 определена у образца I1742 (880 л. н.) из перуанского Tranca (Laramate, Highlands)[15]
См. также
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| CZ | D | E | G | Q | R | O | A | S | X | Y | N1 | N2 | ||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | UK | I | N1a | W | ||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры IWX | ||||||||||||||||||||||
Примечания
- Захаров И. А. Центральноазиатское происхождение предков первых американцев // Первые американцы. 2003. № 11. С. 139-144.
- Васильев С. А., Березкин Ю. Е., Козинцев А. Г., Пейрос И. И., Слободин С. Б., Табарев А. В. Заселение человеком Нового Света: Опыт комплексного исследования. СПб: Нестор-история, 2015.
- Volodko N. V., Starikovskaya E. B., Mazunin I. O. et al., 2008. Mitochondrial diversity in Arctic Siberians, with particular reference to the evolutionary history of Beringia and Pleistocenic peopling of the Americas // American Journal of Human Genetics. Vol. 82. № 5. P. 1084–1100.
- Natalia V. Volodko, Elena B. Starikovskaya, Ilya O. Mazunin et al., «Mitochondrial Genome Diversity in Arctic Siberians, with Particular Reference to the Evolutionary History of Beringia and Pleistocenic Peopling of the Americas», The American Journal of Human Genetics 82, 1084—1100, May 2008. DOI 10.1016/j.ajhg.2008.03.019.
- D. Comas et al. Admixture, migrations, and dispersals in Central Asia: evidence from maternal DNA lineages. European Journal of Human Genetics, 2004
- Guanglin He et al. Genomic insights into the differentiated population admixture structure and demographic history of North East Asians, July 20, 2021 (PDF)
- Late Pleistocene Human Skeleton and mtDNA Link Paleoamericans and Modern Native Americans
- Мексиканская американка из Сибири (недоступная ссылка). Дата обращения: 16 мая 2014. Архивировано 19 мая 2014 года.
- Rasmussen M. et al. The genome of a Late Pleistocene human from a Clovis burial site in western Montana, Nature. 2014. V. 506. P. 225—229.
- J. Víctor Moreno-Mayar et al. Early human dispersals within the Americas, 2018
- Roca-Rada Xavier et al. Ancient mitochondrial genomes from the Argentinian Pampas inform the peopling of the Southern Cone of South America // ISBA9 (PDF
- Xavier Roca-Rada et al. Ancient mitochondrial genomes from the Argentinian Pampas inform the earlypeopling of the Southern Cone of South America, 14 May 2021 (Table 1. Calibrated dates and mitochondrial haplogroups of the 18 Early to Late Holocene individuals from the Argentinian Pampas)
- Supporting Information. Lindo et al. 10.1073/pnas.1620410114
- Kemp Brian M. et al. (2007). Genetic Analysis of Early Holocene Skeletal Remains From Alaska and its Implications for the Settlement of the Americas (недоступная ссылка)
- Cosimo Posth et al. Reconstructing the Deep Population History of Central and South America, 2018
- Xue-feng Sun. Ancient DNA and multimethod dating confirm the late arrival of anatomically modern humans in southern China, February 23, 2021
- María Martinón-Torres et al. On the misidentification and unreliable context of the new “human teeth” from Fuyan Cave (China), May 24, 2021
- Darren Curnoe et al. Reply to Martinón-Torres et al. and Higham and Douka: Refusal to acknowledge dating complexities of Fuyan Cave strengthens our case, June 1, 2021
- Veronika Siska et al. Genome-wide data from two early Neolithic East Asian individuals dating to 7700 years ago, 2017
- Chuan-Chao Wang et al. The Genomic Formation of Human Populations in East Asia, 2020
- Андрей Коньков. Расшифровка древней ДНК рассказала о происхождении южноамериканских индейцев, 20.04.2016
- Bastien Llamas et al. Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas, 2016 // Supplementary Materials. Table S2. List of ancient samples, associated metadata, and sequencing results (separate Excel document
- Chao Ning et al. Ancient genome analyses shed light on kinship organization and mating practice of Late Neolithic society in China, 2021
- Martin Sikora et al. The population history of northeastern Siberia since the Pleistocene, 05 June 2019 (bioRxiv, 2018)
- Pavel Flegontov et al. Paleo-Eskimo genetic legacy across North America, 2017
- Rasmussen, M. et al., «Ancient human gonome sequence of an extinct Palaeo-Eskimo». Nature 463: 757—762.
- Amory S, Crubézy E, Keyser C, Alekseev AN, Ludes B (October 2006). «Early influence of the steppe tribes in the peopling of Siberia». Human Biology 78 (5): 531-49.
- Fuzuki Mizuno et al. Population dynamics in the Japanese Archipelago since the Pleistocene revealed by the complete mitochondrial genome sequences // Scientific Reports, 2021
- A western Eurasian male is found in 2000-year-old elite Xiongnu cemetery in Northeast Mongolia
- Первые результаты работы Лаборатории популяционной генетики
- Казахстанский ДНК-проект Архивная копия от 26 ноября 2016 на Wayback Machine
- Niall P. Cooke et al. Ancient genomics reveals tripartite origins of Japanese populations // Science Advances • 17 Sep 2021 • Vol 7, Issue 38
- Пилипенко А. С. Уникальное захоронение воина гунно-сарматского времени в Западно-Сибирской лесостепи: результаты палеогенетического анализа Архивная копия от 30 декабря 2018 на Wayback Machine // Археология, этнография и антропология Евразии. Том 46 № 4 2018
- Veronika Csaky et al. Early Medieval Genetic Data from Ural Region Evaluated in the Light of Archaeological Evidence of Ancient Hungarians, 2020
- Justin Tackney et al. Molecular analysis of an ancient Thule population at Nuvuk, Point Barrow, Alaska, 09 January 2019
Ссылки
Общие сведения
Гаплогруппа D
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
