Гаплогруппа M (мтДНК)
В популяционной генетике человека гаплогруппой M называют одну из гаплогрупп, выявленных при анализе последовательности митохондриальной ДНК (mtDNA). Эта гаплогруппа широко распространена в Азии[4], особенно в Индии[5]. Тем не менее, макрогруппа М сама представляет собой одну из ветвей митохондриальной гаплогруппы L3, от которой она отделилась 60 — 75 тыс. лет назад[2][1]. TMRCA для базальной неафриканской гаплогруппы M составляет около 49 тыс. лет (95% доверительный интервал: 54,8—43,6 тыс. лет)[6]. Предковая гаплогруппа L3 в свою очередь происходит от потомков гипотетической митохондриальной Евы из Африки.
| Гаплогруппа M | |
|---|---|
 | |
| Тип | мтДНК |
| Время появления | 60 тыс. лет назад |
| Место появления | Азия[1][2] или Африка[3] |
| Предковая группа | Гаплогруппа L3 |
| Субклады | M1, M*, CZ, Q, E, G, D |
| Мутации-маркеры | 263, 489, 10400, 14783, 15043 |
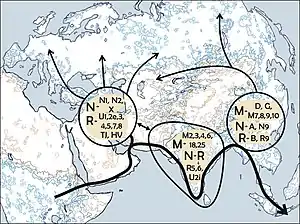
Наличие линии гаплогруппы M в Эфиопии, названной M1, привело к предположению, что гаплогруппа M могла возникнуть в Восточной Африке примерно 60 тыс. л. н., а затем попасть в Азию[7]. Однако, в 2006 году А. Оливьери сообщил[8], что примерно 45—40 тыс. л. н. преобладающие ныне североафриканские гаплогруппы M1 и U6 возникли в Юго-Западной Азии и вместе двинулись в Африку[9].
TMRCA для гаплогруппы M71d составляет около 22,21 тыс. лет (95% доверительный интервал: 14,4—31,52 тыс. лет). Гаплогруппа M71d выявлена в Таиланде, Лаосе и Мьянме и не была обнаружена в современных популяциях Южного Китая[10].
Стивен Оппенгеймер предложил для макрогруппы M название клан Манью. Со ссылкой на исследование эстонских генетиков Томаса Кивисилда и Рихарда Виллемса он писал, что носители субклады M1 переправились через Красное море и прошли на территорию нынешней Эфиопии во времена последнего ледникового периода[11].
Макрогруппа M является предком нескольких гаплогрупп. Важнейшие из них:
- гаплогруппа CZ
- гаплогруппа C — распространены у народов севера Азии, американских индейцев и латиноамериканцев[12][13];
- гаплогруппа Z — распространена в Корее, северном Китае и Центральной Азии. Однако с наибольшей частотой встречается среди некоторых народов России и среди саамов на севере Скандинавии;
- гаплогруппа D — встречается в Средней Азии, на Дальнем Востоке и среди американских индейцев[14];
- гаплогруппа E — встречается в юго-восточной Азии;
- гаплогруппа G — распространена на севере, востоке и в центре Азии[15]. С наибольшей частотой встречается на северо-востоке Сибири;
- гаплогруппа М1 — найдена в западной Евразии и на Африканском Роге[16][1][17]. Возникла в Леванте 40 тыс. лет назад[18]; В Африке и на Ближнем Востоке совместно распространены субклады M1b и M1a, на Кавказе распространена только субклада M1a[19]. Носители митохондриальной гаплогруппы M1, согласно исследованию эстонских генетиков Томаса Кивисилда и Рихарда Виллемса, переправились через Красное море и перешли из Аравии на территорию Африканского рога и Эфиопского нагорья во время последнего ледникового максимума[11][20].
- линии М2 — М6, а также М18 и М25 встречаются только в Индии[21][22][23][24][25][26];
- М7 — М10 и М21 — широко распространены в восточной и юго-восточной Евразии[27][28][29][30][31];
- M11 — встречается только среди китайцев[32];
- M12 — небольшая группа в Японии[32];
- В 2009 году два независимых изданий сообщили об обнаружении у австронезийского населения Мадагаскара субклада M23[33][34];
- гаплогруппы M27 — M29 и Q — локализуются в Меланезии и среди австралийских аборигенов[35][36];
- гаплогруппы M31, M31a1 и M32 — на Андаманских островах[37][38];
- гаплогруппы M33, M33a, M34, M34a, M35, M37a и М39 — M41 — в южной и юго-восточной Азии[39][24][40][41][42][43];
Палеогенетика
- Гаплогруппа M определена у позднепалеолитических образцов F6-620 и AA7-738 из пещеры Бачо Киро в Болгарии[44]
- Гаплогруппу M определили у позднепалеолитических обитателей бельгийской пещеры Гойе (Goyet), живших ок. 34 тыс. л. н. и у позднепалеолитического обитателя из Ла-Рошет (La Rochette) во Франции, жившего 28 тыс. лет назад[45]
- Гаплогруппу M определили у образца Ostuni1 возрастом ок. 27 тыс. л. н. (граветт Италии)[46]
- Гаплогруппу M определили у палелитического жителя острова Окинава Минато 1, жившего 20 тыс. лет назад (стоянка Минатогава)[47]
- M7a определили у образцов SRH 242 (20 000 лет до настоящего времени), SRH 292 бедренная кость (16 000 — 9000 лет до н. в.) и SRH 188 (4574 — 4452 лет до н. в.) из японской пещеры Сирахо-Саонэтабару на острове Исигаки[48]
- M1b определили у образца TAF014 (14,48 тыс. л. н.) иберо-мавританской культуры из марокканского Тафоральта[49][50][51]
- M71d определили у двух неолитических образцов в Южном Китае, датируемых возрастом более 11 тыс. лет. Один образец происходит из лунлиньской пещеры Лаомакао (Laomocao Cave) в Гуанси–Чжуанском автономном районе, второй — из Циншуйюань-Дадуна (Qingshuiyuan Dadong, QSYDD) в провинции Гуйчжоу[10]
- M71+151 определили у образца Longlin_1 (10 686—10 439 лет до настоящего времени) из Китая (Гуанси-Чжуанский автономный район)[52]
- Гаплогруппа M определена у образца DevilsGate2 (5726—5622 лет до н. э.) из пещеры Чёртовы Ворота в Приморье[53]
- M80’D определили у образца Higa020 (начальный Дзёмон, Higashimyou shell midden)[47]
- M7a1a определили у образцов Higa006 (начальный Дзёмон, Higashimyou shell midden, 7,934—7,792 тыс. л. н.), Todo5 (ранний Дзёмон, Todoroki shell midden, 6,210—6,094 тыс. л. н.) и MB-TB27 (поздний Дзёмон, Mabuni hantabaru)[47]
- M7a определили у образца Kaso6 (средний Дзёмон, Kasori shell midden)[47]
- M5 определили у образца La368 культуры Хоа-Бинь (ок. 7888 л. н.) из Лаоса[54]
- Гаплогруппу M (субклад M1[55]) определили у молодой представительницы донеолитической культуры тоалеан (7,3—7,2 тыс. лет до настоящего времени) из известняковой пещеры Леанг Паннинге на острове Сулавеси в Индонезии[56]
- Гаплогруппу M определили у представителя майкопской культуры[57]
- M7c1b и M7b1a определены у представителей культуры Донгшон бронзового века Вьетнама, M21b1a — у образца Ma91 (ок. 4319 л. н.) из Малайзии, M13c — у образца Ma912 (2447 л. н.) из неолита Малайзии, M20 — у образца Vt833 (ок. 4171 л. н.) из неолита Вьетнама[54]
- M1a1b1 обнаружена у образца I4246 (культура колоколовидных кубков) из Камино-де-лас-Йесерас (муниципалитет Сан-Фернандо-де-Энарес) в центральной Иберии (2473—2030 лет до н. э.)[58]
- M1 определена у образца irk078 из Забайкалья (3700 лет до настоящего времени)[59]
- Гаплогруппу M определили у представителя межовской культуры из Каповой пещеры[60]
- M1a1 определили у двух высокостатусных египетских мумий (Nakht-Ankh и Khnum-Nakht) 12-й династии (1985—1773 года до н. э.) эпохи Среднего царства, найденных в Дейр-Рифа (en:Rifeh) в 1907 году сэром Уильямом Флиндерсом Петри и Эрнестом Маккеем[61]
- M1a1, M1a1i, M1a1e и M1a2a определены у мумий из Абусира[62][63]
- M7b1a1 определили у образца BalongKD10 (1568—1409 л. н.) и — у образца BalongKD06 (1500 л. н.) из Китая (Гуанси-Чжуанский автономный район). M7b1a1a определили у образца BalongKD08 (1500 л. н.), М10 — у образца CenxunKP13 (1511 — 1310 л. н.), M7b1a1+(16192) — у образца CenxunKP05 (1467 — 1307 л. н.), M7b1a1a3 — у образцов YiyangKP17(1467—1307 л. н.) и CenxunKP07 (1366—1293 л. н.)[52]
- M65a определили у образца I6197 (1200—800 лет до н. э.), M30c1 определили у образца I12149 (1000—800 лет до н. э.), M30g определили у образца I12134 (1000—800 лет до н. э.), M4a определили у образца I12136 (1000—800 лет до н. э.), M5a определили у образца I13222 (1000—800 лет до н. э.) из долины Сват (Пакистан), M30b у образца I6549 (167—46 лет до н. э., Butkara_IA, долина Сват), M3a2 определили у образца I12450 (1000—800 лет до н. э., Butkara_II), M35b у образца I5397 (968—833 лет до н. э., Katelai_IA, долина Сват)[64], M3a2 определена у образца I12450 (824—792 calBCE, Butkara_IA)[65]
- M1a1 определена у кенийского образца I8809 (Kisima Farm, A5/Porcupine Cave, Pastoral_Neolithic (PN cluster), 3030—2860 л. н.), M1a1f определена у кенийского образца I8820 (Kisima Farm, A5/Porcupine Cave, Pastoral_Neolithic (PN cluster), 2840—2740 л. н.), M1a1b (скорее всего) определена у кенийского образца I8923 Rigo Cave (GrJh3), Pastoral_Neolithic/Elmenteitan (PN cluster), 2690—2350 л. н.), M1a1b определена у кенийского образца I8830 (Naivasha Burial Site, Pastoral_Neolithic (PN cluster), 2360—2210 л. н.)[66]
- M7a1a4 определили у образца Yayoi_1 (Период Яёй, Япония)[67]
- M10a1a1a выявлена у образца scy332 (248 — 391 гг.) из могильника близ молдавского села Глиное[68]
- M6 выявлена у образца Szeg076.422B из аварского могильника VI—VII веков в Сегвар-Оромдюль (Szegvár-Oromdülő) в Венгрии[69]
- M7b1a1a1 определили у образца имперского периода Кофун JpIw33 (1295—1355 л. н.)[67]
- M1a1b1d определили у образца PLEper200, а M1b2* — у образца MHper26 из Карпатского бассейна (X—XI вв.)[70]
- M7c1a1a1 определили у представительницы чияликской культуры (XI—XIV века)[71]
- M7c2 определена у вьетнамского образца Vt719, похороненного 229±69 лет назад[54]
Примечания
- Revathi Rajkumar et al., Phylogeny and antiquity of M macrohaplogroup inferred from complete mt DNA sequence of Indian specific lineages, BMC Evolutionary Biology 2005, 5:26 doi:10.1186/1471-2148-5-26
- Gonzalez et al., Mitochondrial lineage M1 traces an early human backflow to Africa, BMC Genomics 2007, 8:223 doi:10.1186/1471-2164-8-223
- Semino et al. (2000), The Genetic Legacy of Paleolithic Homo sapiens in Extant Europeans: A Y Chromosome Perspective, Science. 2000 Nov 10;290(5494):1155-9.
- Ghezzi et al. (2005), Mitochondrial DNA haplogroup K is associated with a lower risk of Parkinson’s disease in Italians, European Journal of Human Genetics (2005) 13, 748—752.
- Edwin et al. (2002), Mitochondrial DNA diversity among five tribal populations of southern India, CURRENT SCIENCE, VOL. 83, NO. 2, 25 JULY 2002.
- Cosimo Posth et al. Pleistocene Mitochondrial Genomes Suggest a Single Major Dispersal of Non-Africans and a Late Glacial Population Turnover in Europe, 2016
- Quintana-Murci L., Semino O., Bandelt H-J., Passarino G., McElreavey K. et al. (1999) Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern // Africa Nat Genet 23: 437–441
- Olivieri A., Achilli A., Pala M., Battaglia V., Fornarino S.. et al. (2006) The mtDNA Legacy of the Levantine Early Upper Paleolithic in Africa. Science 314: 1767
- Adimoolam Chandrasekar et al. Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor, October 20
- Fan Bai, Xinglong Zhang, Xueping Ji, Peng Cao, Xiaotian Feng, Ruowei Yang, Minsheng Peng, Shuwen Pei, Qiaomei Fu. Paleolithic genetic link between Southern China and Mainland Southeast Asia revealed by ancient mitochondrial genomes, 11 July 2020
- Stephen Oppenheimer. Out of Eden. 2004, Constable and Robinson ISBN 1-84119-894-3 UK title of The Real Eve.
- Haplogroup C
- Haplogroup C1
- D. Comas et al., Admixture, migrations, and dispersals in Central Asia: evidence from maternal DNA lineages. EJHG, 2004
- Haplogroup G
- Haplogroup M1
- A.D. Holden et al., MtDNA variation in North, East, and Central African populations gives clues to a possible back-migration from the Middle East Архивная копия от 3 марта 2016 на Wayback Machine, Program of the Seventy-Fourth Annual Meeting of the American Association of Physical Anthropologists (2005)
- Александр Марков. Эволюция человека: обезьяны, кости и гены. 2011
- Erwan Pennarun et al. Divorcing the Late Upper Palaeolithic demographic histories of mtDNA haplogroups M1 and U6 in Africa
- Kivisild Т. et al. (1999) Возможное влияние западноазиатских материнских генетических линий на жителей Восточной Африки в эпоху после Последнего ледникового максимума. Cold Springs Harbour Symposium on Human Origins & Dicease.
- Haplogroup M2
- Haplogroup M3
- Haplogroup M4
- K. Thangaraj et al, In situ origin of deep rooting lineages of mitochondrial Macrohaplogroup 'M' in India. BMC Genomics, 2006
- Haplogroup M5
- Haplogroup M6
- Haplogroup M7
- Haplogroup M8
- Haplogroup M9
- Haplogroup M10
- Haplogroup M21
- Haplogroups M11 & 12
- Dubut ; V; Cartault, F; Payet, C; Thionville, MD; Murail, P. et al. Complete mitochondrial sequences for haplogroups M23 and M46: insights into the Asian ancestry of the Malagasy population (англ.) // Human Biology : journal. — 2009. — Vol. 81, no. 4. — P. 495—500. — doi:10.3378/027.081.0407. — PMID 20067372.
- Ricaut, F. X.; Razafindrazaka, H; Cox, MP; Dugoujon, JM; Guitard, E; Sambo, C; Mormina, M; Mirazon-Lahr, M; Ludes, B; Crubézy, Eric et al. A new deep branch of eurasian mtDNA macrohaplogroup M reveals additional complexity regarding the settlement of Madagascar (англ.) // BMC Genomics : journal. — 2009. — Vol. 10. — P. 605. — doi:10.1186/1471-2164-10-605. — PMID 20003445.
- Haplogroup M27-29
- Haplogroup Q
- Haplogroup M31
- Haplogroup M32
- Haplogroup M33
- Haplogroup M34
- Haplogroup M35
- Haplogroup M39
- Haplogroup M40
- Jean-Jacques Hublin et al. Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria, 11 May 2020
- Cosimo Posth et al. Pleistocene Mitochondrial Genomes Suggest a Single Major Dispersal of Non-Africans and a Late Glacial Population Turnover in Europe, 2016
- Qiaomei Fu et al. The genetic history of Ice Age Europe, 2016
- Fuzuki Mizuno et al. Population dynamics in the Japanese Archipelago since the Pleistocene revealed by the complete mitochondrial genome sequences // Scientific Reports, 2021
- Ken-ichi Shinoda, Noboru Adach. Ancient DNA Analysis of Palaeolithic Ryukyu Islanders // This text is taken from New Perspectives in Southeast Asian and Pacific Prehistory, edited by Philip J. Piper, Hirofumi Matsumura and David Bulbeck, published 2017 by ANU Press, The Australian National University, Canberra, Australia
- M. van de Loosdrecht el al. Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations, Science (2018)
- Ancient Epigenomics
- Supplementary Materials for. Table S16. Y-haplogroup assignment for six Taforalt males. All individuals could be assigned to haplogroup E1b1b, and five of them more specifically to E1b1b1a1 (M-78).
- Tianyi Wang et al. Human population history at the crossroads of East and Southeast Asia since 11,000 years ago // Cell, June 24, 2021
- Veronika Siska et al. Genome-wide data from two early Neolithic East Asian individuals dating to 7700 years ago, 2017
- Hugh McColl et al. Ancient Genomics Reveals Four Prehistoric Migration Waves into Southeast Asia, 2018
- SI Table 17 // Supplementary information for: Genome of a middle Holocene hunter-gatherer from Wallacea
- Selina Carlhoff et al. Genome of a middle Holocene hunter-gatherer from Wallacea, 25 August 2021
- Sokolov A. S. et al. Six complete mitochondrial genomes from Early Bronze Age humans in the North Caucasus
- Iñigo Olalde et al. The genomic history of the Iberian Peninsula over the past 8000 years, 2019
- Gülşah Merve Kılınç et al. Human population dynamics and Yersinia pestis in ancient northeast Asia, 2021
- Morten E. Allentoft et al. «Population genomics of Bronze Age Eurasia», 2015
- Konstantina Drosou et al. The kinship of two 12th Dynasty mummies revealed by ancient DNA sequencing, 2018
- Verena J. Schuenemann et al. Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods, 30 May 2017
- Jean-Philippe Gourdine, S.O.Y Keita, Jean-Luc Gourdine, Alain Anselin. Ancient Egyptian Genomes from northern Egypt: Further discussion
- Vagheesh M Narasimhan et al. The Genomic Formation of South and Central Asia, March 31, 2018
- Vagheesh M. Narasimhan et al. The formation of human populations in South and Central Asia, 06 Sep 2019
- Mary E. Prendergast et al. Ancient DNA reveals a multistep spread of the first herders into sub-Saharan Africa (Table S7. (separate file) mtDNA haplogroups), 2019
- Niall P. Cooke et al. Ancient genomics reveals tripartite origins of Japanese populations // Science Advances • 17 Sep 2021 • Vol 7, Issue 38
- Maja Krzewińska et al. Ancient genomes suggest the eastern Pontic-Caspian steppe as the source of western Iron Age nomads, 2018
- Aranka Csősz et al. Maternal Genetic Ancestry and Legacy of 10th Century AD Hungarians, 16 September 2016
- Kitti Maár et al. Maternal Lineages from 10–11th Century Commoner Cemeteries of the Carpathian Basin, March 2021
- Bea Szeifert et al. Tracing genetic connections of ancient Hungarians to the 6—14th century populations of the Volga-Ural region, February 08, 2022
См. также
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| CZ | D | E | G | Q | R | O | A | S | X | Y | N1 | N2 | ||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | UK | I | N1a | W | ||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры IWX | ||||||||||||||||||||||
Ссылки
- M YFull MTree 1.01.15226 (under construction)
- PhyloTree.org - mtDNA tree Build 17 (18 Feb 2016): subtree M
- Spread of Haplogroup M, from National Geographic
- Tree of M haplogroup as for 2006
- Haplogroup M (mtDNA) interest group on Facebook
- China DNA interest group on Facebook
- [www.familytreedna.com/public/india The India DNA] geographical project at Family Tree DNA
- [www.familytreedna.com/public/china The China DNA] geographical project at Family Tree DNA
- Another tree emphasizing the Andamanese and Nicobarese populations in comparison with other peoples with high M presence
- K.Tharanghaj et al. In situ origin of deep rooting lineages of mitochondrial Macrohaplogroup M in India (PDF document)
{kind=link}
{kind=link}
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||