Гаплогруппа R (мтДНК)
Гаплогруппа R — гаплогруппа митохондриальной ДНК человека.
| Гаплогруппа R | |
|---|---|
_%2526_subclades.PNG.webp) | |
| Тип | мтДНК |
| Время появления | 55 тыс. лет назад |
| Место появления | Юго-западная Азия |
| Предковая группа | N |
| Сестринские группы | A, S, X, Y, N1, N2 |
| Субклады | B, F, R0, pre-JT, P, UK |
| Мутации-маркеры | 12705, 16223 |
Происхождение
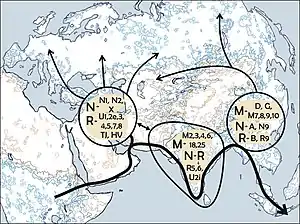
Гаплогруппа R является потомком макрогруппы N и по древности, широте распространения и количеству групп-потомков сама может быть названа макрогруппой. Вероятно, возникла на Ближнем Востоке вскоре (всего через несколько тысяч лет) после заселения его людьми (около 55 тыс. л. н.). В дальнейшем из этого региона была осуществлена экспансия, так что носители группы R (зачастую вместе с другими субкладами группы N) распространились: на юг в Аравию и северо-восточную Африку, на восток по Иранскому нагорью, далее в Центральную Азию до Алтая и на Индостан, на север в Анатолию, на Кавказ и далее в восточную Европу.
Субклады R5, R6, R7, R8, R30, R31 являются автохтонными гаплогруппами в Индии. В наборе данных по Центральной Азии (Comas et al. 2004) из 232 линий только 2 принадлежат к R5[1].
К потомкам этой группы относятся B, UK (и её потомки U и K), F, R0 (и её потомки HV, H, и V), а также pre-JT (и её потомки JT, J и T).
Палеогенетика
Митохондриальная гаплогруппа R* была обнаружена у палеосибиряка из Усть-Ишима (Усть-ишимский человек), жившего 45 тыс. лет назад[2], но в 2016 году команда Позника классифицировала у усть-ишимца митохондриальную гаплогруппу U*[3].
Гаплогруппа R определена у позднепалеолитического образца CC7-2289 (ок. 44 тыс. л. н.) из пещеры Бачо Киро в Болгарии[4]
К гаплогруппе R* относится митохондриальный геном молочного зуба из пещеры Фумане (it:Grotta di Fumane), найденный в слое возрастом 41 110 — 38 500 лет (протоориньяк)[5].
У останков девушки Пальиччи 23 (en:Paglicci 23) из итальянской пещеры Пальиччи возрастом около 28 тыс. лет была обнаружена кембриджская эталонная последовательность (en:Cambridge Reference Sequence) HVR1, указывающая на то, что индивидуум имел либо митохондриальную гаплогруппу R, либо митохондриальную гаплогруппу H[6]. Анализ ДНК кроманьонца Paglicci-12, жившего около 24 тыс. лет назад, показал, что он принадлежал к митохондриальной гаплогруппе R[7].
У обитательницы Афонтовой горы AG-3, жившей 16 930—16 490 лет назад, была обнаружена субклада R1b.
R1 определена у образца yak025 (14 865—14 590 лет до н. э.) из пещеры Хайыргас (Khaiyrgas Cave, дюктайская культура)[8]. У образца Khaiyrgas-1 из Якутии (16,9 тыс. л. н.) определена R1b[9].
R9c1b определили у образца Qihe3 (11 747—11 356 лет до настоящего времени) из провинции Фуцзянь в Китае[10].
R2-a* определили у образца AH2 (8205—7756 лет до н. э.) из Тепе Абдул-Хосейн (Tepe Abdul Hosein) в Иране (шахрестан Нурабад)[11].
R11[12] определили у образца NE35 (AR8.9K, ARpost9K, 9131—8770 лет до настоящего времени) из Амурского региона (Китай)[13].
R3 обнаружена у представителя неолита Венгрии, жившего ок. 7500 лет назад[14].
R1b определена у мезолитического обитателя Южного Оленьего острова[15].
R1a определили у энеолитических образцов I2055 (Unakozovskaya, 6533 л. н.) и I2056 (Unakozovskaya, 6477,5 л. н.) с Северного Кавказа (Унакозовская пещера на ручье Мешоко)[16].
R1b1 определена у представительницы эстонской культуры боевых топоров SOP002 (2864—2495 лет до н. э.) из могильника Сопе в уезде Ида-Вирумаа[17].
R1a определили у представителя новосвободненского этапа майкопской культуры I6268 (5564 л. н.) из урочища Клады в Адыгее[16].
R1a1a определили у образца MK5009.A0101 с Северного Кавказа (Marinskaya 5, 4710 л. н.)[16].
R1b1 определена у образца GLZ003 из Глазково (Glazkovskoe predmestie), датирующегося возрастом 4519—4417 л. н. (ранний бронзовый век Байкала)[18].
R1a определили у представителей дарквети-мешоковской культуры (накольчатой жемчужной керамики) эпохи позднего энеолита[19].
R1b определена у образца эпохи бронзы Anosovo-1 из Забайкалья (ок. 4 тыс. л. н.)[9].
R1b определили у представителя посткатакомбной лолинской культуры NV3001 (Nevinnomiskiy 3, 3970,5 л. н.) в Калмыкии (Лола)[16].
R1b1 определили у образца BIY002.A0101 (III—II века до н. э., саргатский горизонт) из урочища Горная Бития на реке Ишим, расположенного в 8 км к юго-западу от деревни Нижняя Ильинка (Омская область)[20].
R1b1 определили у таримской мумии L5213 (2000—1800 л. н.)[21].
R11[12] обнаружена у неклассифицированного образца KHI001 (средне-поздний бронзовый век) из Восточно-Евразийской степи с Алтая[22].
R1a определили у образца ALA135 (1876—1636 лет до н. э.) из Телль-Атчана (Алалах, Турция)[23].
R1b1 определили у представителей унетицкой культуры из Чехии I14191 (Czech_EBA_Unetice, 3850 л. н.)[24].
R1b1 обнаружена у образца I3860 (Oy-Dzhaylau III, 3626 л. н.) из среднего-позднего бронзового века Казахстана (Oy_Dzhaylau_MLBA_o)[25].
R9b1a3 определили у образца LayiKD01 (1532—1403 л. н.), R9b1b — у образца QinchangKD14 (1545—1407 л. н.) из Китая (Гуанси-Чжуанский автономный район)[10].
R1a1a обнаружена в образце из могильника Канагех (Kanagegh, Армения) возрастом 1500—1300 лет до нашей эры[26].
R1b1 обнаружена у обитателя Неркин-Геташена (Nerqin Getashen, Армения), жившего 1400—1200 лет до нашей эры[26].
У образцов из района Хафт-Тапе периода среднего Элама определены митохондриальные гаплогруппы R2 и R5[27].
R6 определена у представителя кобанской культуры из могильника Заюково-3, расположенного близ села Заюково в Баксанском районе Кабардино-Балкарской республики (VIII—VI века до н. э.)[28].
R9 определена у образца из висячих гробов из Юньнани[29].
R0a1 определена у образцов с христианского кладбища R (∼650—1000 гг.) на острове Кулубнарти в Северном Судане[30].
R11a обнаружена у позднесредневекового образца DEC01/SHR001 из Монголии[22].
R1a1a обнаружена у норвежского образца VK114 из Тронхейма (XII—XIII век)[31].
R9b1b определили у образца HuatuyanNL18 (500 л. н.), R+16189 — у образца HuatuyanNL19 (455—294 л. н.) из Китая (Гуанси-Чжуанский автономный район)[10].
Примечания
- Malliya gounder Palanichamy et al. Phylogeny of Mitochondrial DNA Macrohaplogroup N in India, Based on Complete Sequencing: Implications for the Peopling of South Asia, December 2004
- SUPPLEMENTARY INFORMATION 8. MITOCHONDRIAL GENOME ANALYSIS
- Posnik G. D. et al. (2016) Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences, Nature Genetics, 48, 593–599 (Poznik supp. fig. 15).
- Jean-Jacques Hublin et al. Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria, 11 May 2020
- Палеогенетические данные подтвердили, что создателями протоориньякской культуры были люди современного типа, 27.04.2015
- A 28,000 Years Old Cro-Magnon mtDNA Sequence Differs from All Potentially Contaminating Modern Sequences
- Ancient DNA
- Gülşah Merve Kılınç et al. Investigating Holocene human population history in North Asia using ancient mitogenomes, 12 June 2018 // Dataset 1 / Table S4
- Gülşah Merve Kılınç et al. Human population dynamics and Yersinia pestis in ancient northeast Asia, 2021
- Tianyi Wang et al. Human population history at the crossroads of East and Southeast Asia since 11,000 years ago // Cell, June 24, 2021
- Broushaki F. et al. (2016 July). “Early Neolithic genomes from the eastern Fertile Crescent”. Science. 353 (6298). DOI:10.1126/science.aaf7943. PMC 5113750. PMID 27417496. Проверьте дату в
|date=(справка на английском) - R11 MTree
- Xiaowei Mao et al. The deep population history of northern East Asia from the Late Pleistocene to the Holocene, May 27, 2021
- Fu. Q. et al. The genetic history of Ice Age Europe, Nature, published online 2 May 2016.
- Alissa Mittnik et al. The Genetic History of Northern Europe, 03.03.2017
- Chuan-Chao Wang et al. The genetic prehistory of the Greater Caucasus, May 16, 2018
- Lehti Saag, Sergey V. Vasilyev, Svetlana V. Oshibkina et al. Genetic ancestry changes in Stone to Bronze Age transition in the East European plain (Table 1), July 03, 2020 (bioRxiv)
- He Yu et al. Paleolithic to Bronze Age Siberians Reveal Connections with First Americans and across Eurasia, May 20, 2020
- Chuan-Chao Wang et al. The genetic prehistory of the Greater Caucasus, May 16, 2018
- Guido Alberto Gnecchi-Ruscone et al. Ancient genomic time transect from the Central Asian Steppe unravels the history of the Scythians // Science Advances. Vol. 7, Issue 13, 26 Mar 2021
- Extended Data Table 1 A summary of the Bronze Age Xinjiang individuals reported in this study. From: Fan Zhang et al. The genomic origins of the Bronze Age Tarim Basin mummies // Nature, 2021
- Choongwon Jeong et al. A dynamic 6,000-year genetic history of Eurasia's Eastern Steppe, 2020
- Tara Ingman et al. Human mobility at Tell Atchana (Alalakh), Hatay, Turkey during the 2nd millennium BC: Integration of isotopic and genomic evidence // Plos One, June 30, 2021
- Nick Patterson et al. Large-scale migration into Britain during the Middle to Late Bronze Age // Nature, 22 December 2021
- Vagheesh M. Narasimhan et al. The formation of human populations in South and Central Asia (англ.) // Science. — 2019-09-06. — Vol. 365, iss. 6457. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.aat7487. (The Genomic Formation of South and Central Asia, bioRxiv)
- Margaryan, Derenko et al. Eight Millennia of Matrilineal Genetic Continuity in the South Caucasus, Current Biology 27, 1–6 July 10, 2017, DOI: 10.1016/j.cub.2017.05.087
- تعیین توالی DNA میتوکندری (mtDNA)بقایای اسکلتی دورهی ایلام میانی محوطهی باستانی هفتتپه
- Eugenia Boulygina et al. Mitochondrial and Y-chromosome diversity of the prehistoric Koban culture of the North Caucasus, 2020
- Xiaoming Zhang et al. A Matrilineal Genetic Perspectiveof Hanging Coffin Customin Southern China and Northern Thailand, April 24, 20
- Kendra A. Sirak et al. Social stratification without genetic differentiation at the site of Kulubnarti in Christian Period Nubia, February 17, 2021 (Supplementary Figure 5, 6)
- Ashot Margaryan et al. Population genomics of the Viking world, 2019
См. также
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| CZ | D | E | G | Q | R | O | A | S | X | Y | N1 | N2 | ||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | UK | I | N1a | W | ||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры IWX | ||||||||||||||||||||||
Ссылки
Общие сведения
- Ian Logan’s Mitochondrial DNA Site
- PhyloTree.org - mtDNA tree Build 17 (18 Feb 2016): subtree R
- R YFull MTree 1.02.00 (under construction)
Гаплогруппа R
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
