Гаплогруппа N (мтДНК)
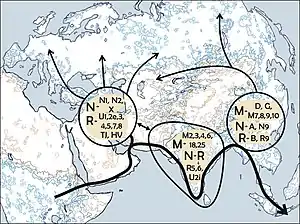
В популяционной генетике человека гаплогруппой N называют одну из гаплогрупп, выявленных при анализе последовательности митохондриальной ДНК (mtDNA). Это необыкновенно широко распространённая гаплогруппа, носители её субклад живут на нескольких континентах[6], поэтому её называют макро-гаплогруппой. В свою очередь, макрогруппа N сама представляет собой одну из ветвей гаплогруппы L3, от которой она отделилась в западной части Азии 50—80 тыс. лет назад. TMRCA для базальной неафриканской гаплогруппы N составляет около 51 тыс. лет (95% доверительный интервал: 55,1—46,9 тыс. лет)[7]. Предковая гаплогруппа L3, в свою очередь, происходит от потомков гипотетической митохондриальной Евы из Африки. Стивен Оппенгеймер предложил для макрогруппы N название клан Насрин, а Брайн Сайкс — Наоми.
| Гаплогруппа N | |
|---|---|
 | |
| Тип | мтДНК |
| Время появления | 65 тыс. лет назад |
| Место появления | западная Азия[1][2]или восточная Африка[3][4][5] |
| Предковая группа | Гаплогруппа L3 |
| Субклады | N1’5, N2, N9, N12, N13, N14, N21, N22, A, R, S, X |
| Мутации-маркеры | 8701, 9540, 10398, 10873, 15301 |
От гаплогруппы N происходят почти все гаплогруппы Европы, Океании, а также носителей индейских и многих азиатских языков. Она образовалась в западной части Азии приблизительно в то же время, когда и другая широко распространенная макрогруппа М. Обе в небольшом количестве встречаются в районе Африканского Рога, куда были привнесены в результате обратной миграции их носителей около 30 тыс. лет назад. Субгаплогруппа N9b существует на относительно высокой частоте в популяциях Юго-Восточной Сибири: удэгейцы — 30,4%, ульчи — 6,9% (Стариковская и др., 2005).
От макрогруппы N происходят другие широко распространенные макрогруппы: A, N1, R, I, S, W, X, Y. Из них:
- Субклада N* была обнаружена у представителей культуры кардиальной керамики[8].
- гаплогруппа А обнаружена в Центральной и восточной Азии, а также среди американских индейцев[9];
- гаплогруппа R — предок ряда распространенных групп, B, F, JT, P, а также HV, К и U, широко представленных в Европе[10];
- гаплогруппа S обнаружена у австралийских аборигенов[11];
- гаплогруппа W распространена преимущественно на западе Евразии[12];
- гаплогруппа X1 обнаружена в Северной Африке, а гаплогруппа X2 — в Европе, Сибири и Америке[13];
- гаплогруппа Y распространена среди нивхов и айнов, а также, в небольшом количестве, среди корейцев, монголов, коряков, ительменов, китайцев, японцев, жителей Малайского архипелага и некоторых тюркских народов[14][15].
Палеогенетика
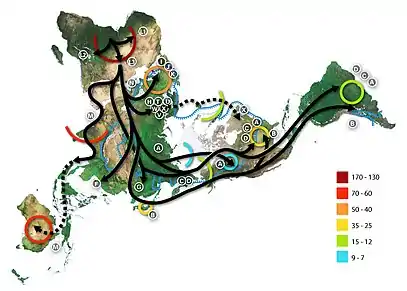
- Гаплогруппу N определили у женского черепа возрастом 47—45 тысяч лет из Конепрусских пещер на холме Золотой конь в Чехии[16].
- Гаплогруппа N определена у позднепалеолитических образцов ВВ7-240 и CC7-335 возрастом более 45 тыс. л. н. из пещеры Бачо Киро в Болгарии[17].
- Гаплогруппа N определена у образца Oase 1 из румынской пещеры Пештера-ку-Оасе (40 тыс. л. н.)[18] и у образца Oase 2[19].
- Гаплогруппа N1 определена у обитателя крымской стоянки Буран-Кая, жившего 36 тыс. лет назад. N1 BuranKaya3A несёт три из восьми мутаций, происходящих до редкой митохондриальной гаплогруппы N1b, наиболее концентрированной на Ближнем Востоке, но широко распространенной от Западной Евразии до Африки[20].
- Гаплогруппа N определена у монгольского черепа Салхит (Salkhit) возрастом 34 950—33 900 лет назад[21].
- Гаплогруппа N определена у обитателя грузинской пещеры Дзудзуана (Dzudzuana Cave), жившего 26 тыс. лет назад[22].
- N1b обнаружена у представителя натуфийской культуры[23].
- N1a1a1 обнаружена у двух анатолийских фермеров из местечка Бонджуклу (Boncuklu), живших 8300—7800 лет до нашей эры, и у двух фермеров из Тепеджик-Чифтлик (Tepecik-Çiftlik), живших примерно 6 тыс. лет назад[24].
- Субклада N1a1a1 была обнаружена у индивида Troc5 стоянки Эльс Трокс в Пиренеях, жившего примерно 7310—7206 лет назад[25].
- Гаплогруппу N определили у двух неолитических скотоводов из Такаркори (Ливия), живших в Зелёной Сахаре 7 тыс. лет назад[26].
- Субклада N1a в эпоху раннего неолита была широко распространена в Европе среди носителей культуры линейно-ленточной керамики[27], однако в дальнейшем вытеснена другими гаплогруппами.
- N1b1 определена у энеолитического (4500—3900/3800 гг. до н. э.) образца I1177 из израильской пещеры близ друзской деревни Пкиин (Peqi’in Cave)[28].
- N1b1a обнаружена у анатолийского образца Tep003 из Тепеджик-Чифтлик (примерно 6 тыс. лет назад)[24].
- Субклада N1b1 определена у потенциального майкопца (нет артефактов в захоронении)[29][30].
- Субклада N1 была обнаружена у представителей синташтинской культуры эпохи бронзы[31].
- Субклада N9a была обнаружена у обитателя археологического объекта эпохи бронзы, расположенного в Аладжосе (Вальядолид, Центральная Испания)[32].
- Субклада N1a1a1a обнаружена у представителя саргатской культуры[33].
- N9b1 определена представительницы культуры Дзёмон IK002 (2500 лет до настоящего времени, Центральная Япония)[34] и у представителей культуры Дзёмон F5 и F23, живших на острове Ребун примерно 3500—3800 лет назад[35], у образца I6341 (Burial 5, JOM_137, 1500—1000 лет до н. э.) из Funadomari, у образцов I13887 (1063 Burial 7, 2191—1982 лет до н. э.) и I13886 (1062 Burial 6, 2136—1959 лет до н. э.) из Rokutsu Shell Mound. У образца I13883 (1050 Burial 2, 984—835 лет до н. э., Rokutsu Shell Mound) определена митохондриальная гаплогруппа N9b2a[36].
- Гаплогруппа N определена у представителя кобанской культуры из могильника Заюково-3, расположенного близ села Заюково в Баксанском районе Кабардино-Балкарской республики (VIII—VII века до н. э.)[37].
- N8 определена у образца из висячих гробов из северного Таиланда, N9a3 — из Юньнани[38].
- N9a обнаружена образца из района Красноярска (400—100 лет до н. э.)[39]
- N5 определили у мумии 1-го тыс. до н. э. — начала нашей эры из коллекции Государственного музея изобразительных искусств имени А. С. Пушкина № 4 (1,1а 5302 ГМИИ)[40][41]
- N9a9 обнаружена у двух детей, найденных в окрестностях крепости Капидава в Добрудже (880—990 года)[39]
- N1b1a обнаружена у жительницы Пскова X—XI веков (образец VK158)[42].
Примечания
- Richards et al. (2006), A Model for the Dispersal of Modern Humans out of Africa (недоступная ссылка), Nucleic Acids and Molecular Biology, 10.1007/3-540-31789-9
- Chandrasekar et al. (2007), YAP insertion signature in South Asia, Ann Hum Biol. 2007 Sep-Oct;34(5):582-6.
- Watson. Mitochondrial Footprints of Human Expansions in Africa (англ.) : journal. — 1997.
- Kivisild et al. The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations (англ.) : journal. — 2003.
- Kivisild et al. Genetic Evidence of Modern Human Dispersals in South Asia // The Evolution and History of Human Populations in South Asia (англ.). — 2007.
- Haplogroups I & N.
- Cosimo Posth et al. Pleistocene Mitochondrial Genomes Suggest a Single Major Dispersal of Non-Africans and a Late Glacial Population Turnover in Europe, 2016
- A common genetic origin for early farmers from Mediterranean Cardial and Central European LBK cultures
- Haplogroup A
- Haplogroup R*
- Haplogroup S
- Haplogroup W
- Haplogroup X
- Haplogroup Y
- Miroslava Derenko, Boris Malyarchuk, Tomasz Grzybowski, Galina Denisova, Irina Dambueva, Maria Perkova, Choduraa Dorzhu, Faina Luzina, Hong Kyu Lee, Tomas Vanecek, Richard Villems, Ilia Zakharov. Phylogeographic Analysis of Mitochondrial DNA in Northern Asian Populations. // American Journal of Human Genetics, 2007 November; 81(5): 1025—1041.
- Kay Prüfer et al. A genome sequence from a modern human skull over 45,000 years old from Zlatý kůň in Czechia, 07 April 2021
- Jean-Jacques Hublin et al. Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria, 11 May 2020
- An early modern human from Romania with a recent Neanderthal ancestor
- Siska, Veronika (2019). “Chapter 2: Palaeolithic Oase genome implies diversification and extinction events across Eurasia” (PDF). Human population history and its interplay with natural selection. University of Cambridge (Thesis). DOI:10.17863/CAM.31536. (Creative Commons Attribution 4.0 International (CC BY 4.0))
- Andrew Bennett, Sandrine Prat, Stephane Pean, Laurent Crepin, Alexandr Yanevich, Simon Puaud, Thierry Grange, Eva-Maria Geigl. The origin of the Gravettians: genomic evidence from a 36,000-year-old Eastern European, 2019
- Thibaut Devièse et al. Compound-specific radiocarbon dating and mitochondrial DNA analysis of the Pleistocene hominin from Salkhit Mongolia, 30 January 2019
- Iosif Lazaridis et al. Paleolithic DNA from the Caucasus reveals core of West Eurasian ancestry, 2018
- Iosif Lazaridis et al. The genetic structure of the world's first farmers, 2016.
- Gülşah Merve Kılınç et al. The Demographic Development of the First Farmers in Anatolia, OCTOBER 10, 2016
- Haak, W. et al. (2015), Massive migration from the steppe is a source for Indo-European languages in Europe
- Stefania Vai et al. Ancestral mitochondrial N lineage from the Neolithic ‘green’ Sahara, 05 March 2019
- Haak (2005) and others, цитата в: Dienekes (2005) in External links, People below
- Éadaoin Harney et al. Ancient DNA from Chalcolithic Israel reveals the role of population mixture in cultural transformation, 2018
- В поисках корней майкопской культуры (комментарий Недолужко)
- Sokolov A. S. et al. Six complete mitochondrial genomes from Early Bronze Age humans in the North Caucasus
- Morten E. Allentoft et al. «Population genomics of Bronze Age Eurasia»
- Sara Palomo-Díez et al. An unexpected case in the prehistory of the Iberian Peninsula: Biogeographical origin analysis through mitochondrial DNA, September 16, 2017
- Пилипенко А. С. и др. Палеогенетическое исследование родства погребенных из курганов саргатской культуры в Барабинской лесостепи (Западная Сибирь)
- Takashi Gakuhari et al. Jomon genome sheds light on East Asian population history, 2019
- Late Jomon male and female genome sequences from the Funadomari site in Hokkaido, Japan
- Chuan-Chao Wang et al. Genomic Insights into the Formation of Human Populations in East Asia (Supplementary Tables // Table 1. Newly reported ancient individuals), 2021
- Eugenia Boulygina et al. Mitochondrial and Y-chromosome diversity of the prehistoric Koban culture of the North Caucasus, 2020
- Xiaoming Zhang et al. A Matrilineal Genetic Perspectiveof Hanging Coffin Customin Southern China and Northern Thailand, April 24, 20
- Ioana Rusu, Alessandra Modi, Stefania Vai, Elena Pilli, Cristina Mircea, Claudia Radu, Claudia Urduzia, Zeno Karl Pinter, Vitalie Bodolica, Catalin Dobrinescu, Montserrat Hervella, Octavian Popescu, Martina Lari, David Caramelli, Beatrice Kelemen. Maternal DNA lineages at the gate of Europe in the 10th century AD // PloS ONE. 2018. V. 13. № 3. e0193578
- Е. Б. Яцишина, Е. С. Булыгина, С. В. Васильев, Р. М. Галеев, Н. В. Слободова, С. В. Цыганкова, Ф. С. Шарко. Палеогенетическое изучение древних мумий в Курчатовском институте, 2020
- E. B. Yatsishina, E. S. Bulygina, S. V. Vasilyev, R. M. Galeev, N. V. Slobodova, S. V. Tsygankova, F. S. Sharko. Paleogenetic Study of Ancient Mummies at the Kurchatov Institute, 22 March 2021
- Ashot Margaryan et al. Population genomics of the Viking world, 2019
См. также
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| CZ | D | E | G | Q | R | O | A | S | X | Y | N1 | N2 | ||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | UK | I | N1a | W | ||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры IWX | ||||||||||||||||||||||
Ссылки
- N YFull MTree 1.02.00 (under construction)
- Spread of Haplogroup N, from National Geographic
- PhyloTree.org - mtDNA tree Build 17 (18 Feb 2016): subtree N
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||