Гаплогруппа N (Y-ДНК)
Гаплогруппа N — гаплогруппа Y-хромосомы человека, отмеченная мутацией в ОНП маркере M231.
- О других значениях N1 значения см. N-1
| Гаплогруппа N | |
|---|---|
.PNG.webp) | |
| Тип | Y-ДНК |
| Время появления | разделение с O - 37 000 лет назад, разнообразие на YFull - 22 000 лет назад[1] |
| Место появления | Юньнань |
| Предковая группа | Гаплогруппа NO |
| Мутации-маркеры | M231 |

Определение
Гаплотип N Y-хромосомы родительской линии определяется мутацией в ОНП маркере M231. Мутация b2/b3 в области AZFc Y-хромосомы также типична для N-гаплогруппы. Однако эта мутация возникала независимо четыре раза, и поэтому ее не следует рассматривать как уникальную характеристику линии гаплогруппы[2].
Происхождение
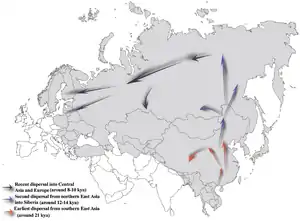
Гаплогруппа N происходит от гаплогруппы NO. Она считается относительно молодой и распространилась на север Евразии со времен последнего ледникового периода. Мужчины, несущие маркер, по-видимому, двинулись на север из-за потепления климата в течение голоцена. Отсутствие N-гаплогруппы у американских индейцев говорит о том, что распространение ее носителей в северо-восточную Азию произошло после появления Берингова пролива. Считается, что гаплогруппа зародилась в Юго-Восточной Азии 15–20 тысяч лет назад, а затем распространилась против часовой стрелки из Монголии и северного Китая в северо-восточную Европу. Субклад N1a1 (N1c) появился примерно 12-8 тысяч лет назад.
Распространение
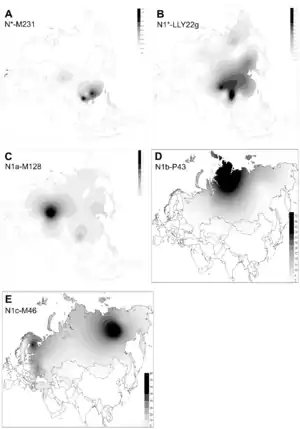
Эта гаплогруппа встречается в Центральной, Северной Европе и повсеместно в европейской и азиатской частях России. Преобладающими носителями данной гаплогруппы являются — финны Финляндии (68 %), балты (46 %)[3], якуты (94 %)[4], нганасаны (92%), энцы (78%), ненцы (74%), удмурты (56 %)[5], манси (57%), буряты России (48 %, у восточных бурят — 78 %)[6], коми (35,1 %). В Казахстане N1a1a-M178 с высокой частотой встречается у родов уак — 64 %[7], сиргели — 65,6 %[8], жалайыры (клан сырманак) — 46 %[7].
Гаплогруппа N является потомком субклады NO1-M214 гаплогруппы NO (F549/M2335/S22380/V4208)[9]. Появилась ок. 36 800 лет назад, последний общий предок современных носителей гаплогруппы N жил 22 100 лет назад[10]. Хотя современное распространение группы N максимально вокруг Балтийского моря и в северной Азии, нахождение древнейших её субклад N* в горах южного Китая (провинция Юньнань) может говорить о появлении мутации M231 именно в том регионе. Гаплогруппа N* (N x M128, P43, TA1) спорадически встречается в Сибири (в основном у коренного населения Южной Сибири). Максимальная частота этой гаплогруппы наблюдается в популяциях Юго-Восточной Азии. По-видимому, линия N* обозначает маршрут миграции палеолитических мигрантов из Юго-Восточной Азии на север, где в свою очередь произошли мутации, определяющие её производные, нижестоящие линии N2 и N1[11].
Подавляющее большинство современных представителей этой группы относятся к ветви N1, наиболее вероятное место появления которой — регион Алтая, Саяны, Прибайкалья, южной Сибири, Монголии или северного Китая, а время — между 15 000 и 10 000 лет назад. По данным эстонских популяционных генетиков субклад N1a2-F1008/L666 разошёлся с ветвью N1a1-M231 около 14 000 лет назад.
Ветвь N1a2b-P43 ранее обозначалась N1c2b и N1b. Субклад N1a2b2a2-VL67 из ветви N1a2b-P43 распространён на Алтае, а также рассеян во всем ареале тюркских народов, вплоть до Турции и Азербайджана. Субклад N1a2b2-Y3195 характерен для коми, удмуртов и в меньшей степени — для татар и марийцев. Там же и южнее распространён субклад N1a1-Y9022. Ареал субклада N1a1a1a1a-L1026 смещён к западу и у коренных народов Урала встречается очень редко. Субклад N1a2a-M128 (ранее N1a) распространён с низкой частотой среди маньчжуров, японцев, сибо, маньчжурских эвенков, корейцев, северных хань, буи, восточных казахов и некоторых других тюркских народов Средней Азии. У казахов очень молода (возможно, менее 1000 лет). Возраст N1a2a-M128 — менее 9000 лет.
Считается, что впервые в Европу она была принесена через Евразию большой миграцией лесных сибирских народов на запад, последняя фаза которой — расселение по Уралу и далее с него по Поволжью и северо-восточной Европе (Русский Север, Финляндия, Прибалтика) — ассоциируется с распространением в этом регионе уральских языков. В Поволжье преобладает субклад N1а-Y9022, разошедшийся с европейским субкладом N1a1a1a1a-L1026 около 7500 л. н. На втором месте в Поволжье находится субклад N1a2b-P43 (ранее N1b). Субклад N1a1a1a1a1a1a1-L1025 сформировался уже в юго-восточной части Балтики около 3000 лет назад и сегодня в значительной степени маркирует именно балтские миграции. N1b-F2930 распространена в Китае, на Тибете, в Японии, Вьетнаме, Индии (телугу), Белоруссии. N2-P189.2 распространена в Сербии, Хорватии, Боснии и Герцеговине, Словакии[12].
Семейное дерево
Семейное древо гаплогруппы N на основе знаний 2015–2016 годов[13].
- NO-M214
- N-M231 Гаплогруппа N, "Niilo"
- N1a-P189.2/N5-B482/-
- -/N1'4-B481/-
- -/N4-F2930/N2-F2930
- -/N4a-CTS39/N2b-F2569 [ISOGG N1b-L732 is a subclade]
- -/N4b-B486/N2a-F830
- N1c-L729/N1'2'3-L729/N1-F1206
- N1c1-M46/N3-Tat/N1b-M46 [formerly N3-Tat/M46 or N1c-Tat/M46] 14000-6000 лет[14]
- N1c1a-M178/-/- 10000-8000 лет
- -/-/N1b1a-F3331
- -/N3c-B496/-
- -/N3a'b-B508/-
- -/N3b-B187/-
- N1c1a1-L708/N3a-L708/N1b1a1-F4325
- -/N3a1-B211/-
- -/N3a2'6-M2110/-
- -/N3a2-M2118/-
- N1c1a1a-L1026/N3a3'6-CTS6967/-
- -/N3a3-CTS10760-
- N1c1a1a1-VL29/N3a3a'b-VL29/-
- -/N3a3c-F4134/-
- N1c1a1a2-Z1936/N3a4-Z1936/-
- -/N3a5-B197/-
- -/N3a5a-F4205/N1b1a1a-F3271
- -/N3a5b-B202/-
- -/N3a6-B479/-
- -/N3a3-CTS10760-
- -/-/N1b1a-F3331
- N1c1a-M178/-/- 10000-8000 лет
- N1c2-F1008/-/N1a-F3163
- -/-N1a1-F1154
- N1c2a-M128/N1-M128/N1a1a1-M128 [formerly N1-M128 or N1a-M128]
- N1c2b-P43/N2a-P43/N1a2-P43 [formerly N2-P43 or N1b-P43] 8000-6000 лет назад
- -/N2a1-B523/-
- -/N2a2-B520/-
- -/-N1a1-F1154
- N1c1-M46/N3-Tat/N1b-M46 [formerly N3-Tat/M46 or N1c-Tat/M46] 14000-6000 лет[14]
- -/N4-F2930/N2-F2930
- N-M231 Гаплогруппа N, "Niilo"
Палеогенетика
- Гаплогруппа N1-M231(xN1a-M128, xN1c-Tat) была основной у культуры Хуншань (6500—5000 лет назад) и более поздних культур этого же региона, а также 3000 лет назад появляется N1c-Tat[15][16].
- N1a1a1a1a-L392/L1026 определён у образца kra001 (2295—2140 лет до н. э.), из Красноярского края (LN/EBA), самый ранний зарегистрированный образец гаплогруппы N1a1a1a1a-L392, базальный к N1a1a1a1a1a-VL29 и N1a1a1a1a2-Z1936[17].
- N1b2-Z4762>F2905>CTS12473* определили у представителя культуры Луншань PLTM311 (2201—2024 лет до н. э., Пинлянтай, Хуайян, Хэнань, Китай)[18].
- Субклад N1a1a1a1a-L392 или N3a3′5 у образца OLS10 (Kunda, Lääne-Viru) из железного века Эстонии (EstIA) датируется 770—430 гг. до нашей эры, у двух других образцов EstIA определён субклад N3a3a[19].
- N1a обнаружен у обитателя свайной постройки № 1 со стоянки Сертея II (жижицкая археологическая культура позднего неолита, 4300 лет назад) и у представителя культуры псковских длинных курганов из кургана с трупосожжением в возможном кривичском[20] захоронении могильника «Девичьи горы» у озера Сенница (1200 лет назад)[21].
- N1a1a1a1a1-Y4341-Y4339∗ обнаружен у образца 84001 (cemetery 1, Nunnan) из Сигтуны (X—XII века)[22][23].
- Два из трёх протестированных представителей пазырыкской культуры имели субклад N1a2b—P43.
- N1b1 определена у неолитчика M54a из местонахождения Houtaomuga на реке Сунгари в провинции Гирин[24].
- N1b1:F4201 (~N-F2905) обнаружен у образца XJS1309_M4 из Сяоцзиншаня (Xiaojingshan) и у образца BS из Бошаня (Boshan)[25].
- N1a1-M46>L1026 (по NevGen) и N1a2-CTS6380 (по NGS) или N1a2-CTS6380>N1a2b2a2-VL67 (по NevGen) определили у двух мужчин из Рубленого города в Ярославле (массовое захоронение № 76, 1238 год)[26].
Известные представители гаплогруппы N
- Рюриковичи (потомки двух или трёх сыновей Ярослава Мудрого; потомки Владимира Мономаха — Мономаховичи) (N-Y4341-Y4339-Y10931)[27].
- Князья Мышецкие (N-Y4341)
- Гедиминовичи (N-L1025-Y13978*)[28]
- Литовские князья Гедройцы (N-L1025-L1027*)
- Татарские князья Кугушевы (N-L1025)[29].
- Значительная часть татарских Каринских князей (Деветьяровы, Арслановы, Хиляловы) (N-P43-Y67014)[29][30].
- Династия Бакинских ханов (N-P43)[31].
- Лаупер, Андрей Анатольевич (N1a2b-P43-Y3185?)[30][32].
- Павел Флоренский (M178, скорее всего Z1928(xZ1933))[33].
- Покачевы — коренное население Покачевского нефтяного месторождения (N1a2b-P43?-Y3185?-VL97?)[34].
- Валдис Пельш (N-L1025).
- зять Яакко Илкка (N-P43-Y3195-Y3185-VL97-FGC38830-Z35051-L665-BY1546)[34].
- Бунины (N-Y9022)[35].
- Генрик Сенкевич[36].
- Олег Кашин
См. также
Примечания
- NO YTree
- Y-DNA Haplogroup N and its Subclades - 2013 isogg.org
- Частоты гаплогрупп у русских и балтов
- Распространение гаплогруппы N в Европу из Сибири
- Гаплогруппа N и путь из Сибири
- В. Н. Харьков , К. В. Хамина , О. Ф. Медведева , К. В. Симонова , Е. Р. Еремина , В. А. Степанов. Генофонд бурят: клинальная изменчивость и территориальная подразделенность по маркерам Y-хромосомы // Генетика. — 2014. — Т. 50, № 2. — С. 203—213.
- Жабагин М. К. Анализ связи полиморфизма Y-хромосомы и родоплеменной структуры в казахской популяции. — 2017.
- Molecular Genetic Analysis of Population Structure of the Great Zhuz Kazakh Tribal Union Based on Y-Chromosome Polymorphism | SpringerLink
- G. David Poznik et al., 2016, «Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences» Nature Genetics, no. 48, pp. 593—599.
- N YTree
- Stepanov V. A. et al. «Origin of Sakha: Analysis of Y-chromosome Haplotypes» / Molecular Biology, 2008. — Volume 42, No 2. — P. 226—237.
- Описание субклад гаплогруппы N на сайте ISOGG
- ISOGG (2016)/Ilumae et al. (2016)/Kang Hu et al. (2015)
- Архивированная копия (недоступная ссылка). Дата обращения: 12 декабря 2020. Архивировано 3 ноября 2016 года.
- Y Chromosome analysis of prehistoric human populations in the West Liao River Valley, Northeast China
- Geographic locations and the Y-chromosome haplogroup distribution of prehistoric populations in this study
- Gülşah Merve Kılınç et al. Human population dynamics and Yersinia pestis in ancient northeast Asia, 06 Jan 2021
- Chao Ning et al. Ancient genome analyses shed light on kinship organization and mating practice of Late Neolithic society in China, 2021
- Lehti Saag et al. The Arrival of Siberian Ancestry Connecting the Eastern Baltic to Uralic Speakers further East, 2019
- Мустафин Х. Х., Альборова И. Э., Семенов А. С, Вишневский В. И. Первые результаты определения Y-хромосомной гаплогруппы для Средневекового захоронения XVI—XVII вв. в Радонеже (Московская область) // Русин. 2017. № 1 (47). DOI: 10.17223/18572685/47/9
- Е. М. Чекунова, Н. В. Ярцева, М. К. Чекунов, А. Н. Мазуркевич. «Первые результаты генотипирования коренных жителей и человеческих костных останков из археологических памятников Верхнего Подвинья». С. 287—294. Таблица на с. 294. // Археология озёрных поселений IV—II тыс. до н. э.: хронология культур и природно-климатические ритмы. — СПб.: ООО «Периферия», 2014.
- N-Y4339 YTree
- Genomic and Strontium Isotope Variation Reveal Immigration Patterns in a Viking Age Town, 2018
- Evolution of Steppe, Neolithic, and Siberian ancestry in Eurasia (ISBA 8, 19th Sep)
- Melinda A. Yang et al. Ancient DNA indicates human population shifts and admixture in northern and southern China, 14 May 2020
- Мустафин Х. Х., Энговатова А. В., Альборова И. Э., Тарасова А. А. Палеогенетическая экспертиза останков из одного массового захоронения 1238 года в Ярославле // Археология Подмосковья. Материалы научного семинара. Выпуск 18. М.: Институт археологии РАН, 2022. С. 111
- Происхождение Рюриковичей с точки зрения ДНК-генеалогии
- Происхождение Гедиминовичей с точки зрения ДНК-генеалогии
- Tatarstan Y-DNA SNP
- familytreedna.com/public/N%20Russia%20%20DNA%20Project?iframe=ycolorized
- YSearch (недоступная ссылка)
- familytreedna.com/public/Finland/default.aspx?section=yresults
- https://web.archive.org/web/20190203035912/http://pereformat.ru/wp-content/uploads/2019/01/florenskiy-dna.pdf
- familytreedna.com/public/N%20Russia%20%20DNA%20Project?iframe=yresults
- ДНК-тесты замечательных людей: Иван Бунин (первые сюрпризы) | Переформат.ру
- Wędzony Septentrion (польск.). Newsweek.pl. Дата обращения: 18 мая 2020.
Ссылки
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 GHIJK | |||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
