Гаплогруппа C (Y-ДНК)
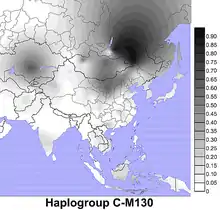
Гаплогруппа C (RPS4Y=M130) — гаплогруппа ДНК Y-хромосомы, распространённая в Восточной и Центральной Азии, в Северной Америке и в Австралии, также отчасти в Европе, Леванте и Японии.
| Гаплогруппа C | |
|---|---|
 | |
| Тип | Y-ДНК |
| Время появления | 60 тысяч лет назад |
| Место появления | Южная Азия |
| Предковая группа | CF |
| Сестринские группы | F |
| Субклады | C1, C2 |
| Мутации-маркеры | M130/Page51/RPS4Y711 |
Эта гаплогруппа имеет две основных субклады: С1 (F3393 / Z1426; ранее CxC3, то есть старый C1, старый С2, старый С4, старый C5 и старый C6) и С2 (M217).
Это одна из очень древних гаплогрупп, С F D E, распространившихся из вторичного Ближневосточного очага этногенеза. В период с 88000 по 68500 проживали потомки СТ, в период 68500 по 65200 проживали потомки DE CF и только с этого времени начинается бурное ветвление С F D E, причём Е начали распространяться 52300 лет назад из Эфиопского нагорья, D и С где то в Центральной Азии и только F стартовали с Ближнего Востока 48800 лет назад.
Определяется полиморфизмами уникальных событий M130/RPS4Y711, P184, P255, P260 — все они являются мутациями SNP. Является родственной гаплогруппе F, которая также является потомком более древней гаплогруппы CF. В отличие от других гаплогрупп, возникших в тот же период (включая потомков), все подклассы гаплогруппы CF являются неафриканскими, а гаплогруппа C интересна тем, что продвинулась на восток особенно далеко.
Распространение
Распространена у тунгусоязычных народов, нивхов, монголов, бурят, калмыков, хазарейцев и казахов[1], высокой концентрации достигает подгруппа С2 (ранее обозначалась как С3)[2][3]. C2 (M217) встречается до 50% у казахов-уйсунов Старшего жуза, 66% у казахов-кереев Среднего жуза, C2c1a1a1 у 86% казахов-коныратов Среднего жуза, C2b1a2-M48 (ранее С3с) до 77% у казахов-алимулы, до 69% у казахов-байулы Младшего жуза[1] и у киргизов (племена монолдор и жору)[4].
Теории миграции
_2017.png.webp)
Определяющая гаплогруппу C мутация M130 произошла, скорее всего, в период 88000-68500 лет назад когда ОНП-мутации M168 из Y-хромосомной гаплогруппы BT выделилась Y-хромосомная гаплогруппа 'CT, а из последней — посредством мутации P143 гаплогруппа CF. По данным компании YFull Y-хромосомная гаплогруппа C-M130 образовалась 65,9 тыс. л. н. Последний общий предок современных носителей гаплогруппы C-M130 жил 48,4 тыс. л. н. C1-F3393 образовалась 48 800 л. н. Последний общий предок современных носителей гаплогруппы C1-F3393 жил 47 200 л. н.
.PNG.webp)
C2-M217 образовалась 48 800 л. н. Последний общий предок современных носителей гаплогруппы C2-M217 (ранее C3) жил 34 000 лет назад[6].
Гаплогруппа С претерпела первоначальное быстрое расширение вскоре после 54 тыс л. н. и к 50 тыс. л. н. уже имела восемь субветвей. Линии C1-F3393 найдены сегодня от Европы до Восточной, Юго-Восточной Южной, Азии и в Океании, в то время как более распространённые линии C2-M217 в настоящее время встречаются в Восточной и Южной Азии, а в Центральной и Западной Азии[7] появились относительно поздно в период экспансии монголов.
Хотя сегодня наибольшая концентрация гаплогруппы C наблюдается среди коренного населения Монголии[8], Дальнего Востока России, Полинезии, аборигенов Австралии. Гаплогруппа С доминирует у западных бурят, эвенков и юкагиров, достигая частот 61,3 %, 69,2 % и 80 % соответственно[9].
Наибольшее разнообразие данной гаплогруппы обнаружено среди населения Индии, из чего некоторые исследователи делают предположение, что она либо произошла, либо существовала наиболее длительное время в своей истории на территории побережья Южной Азии. Субклад C8-CTS5573 был обнаружен у японцев, отобранных в Токио (JPT). Японский субклад C1a1-M8 вместе с европейской и непальской линией C1a2-V20 образуют одну ветвь C1a-CTS11043 (C1'8-CTS824). Это поддерживает гипотезу о том, что общий предок европейской и непальской ветви C1a2-V20 жил в палеолите в Восточной Азии (по данным компании YFull субклад C1a2-V20 образовался 46,7 тыс. л. н., последний общий предок современных носителей субклада C1a2-V20 жил 42,5 тысячи лет назад[10])[11].
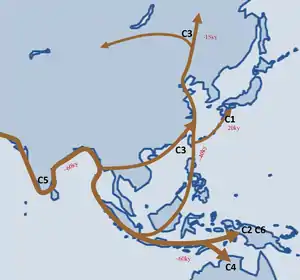
Согласно одной из гипотез, гаплогруппа C связана с теорией прибрежной миграцией первобытных людей через Южную Азию в Юго-Восточную Азию и Австралию, а затем на север вдоль по азиатскому побережью. Другая гипотеза предполагает внутренний континентальный маршрут миграции (Inland Route).
По мнению ряда исследователей, гаплогруппы C и D попали в Восточную Азию вместе, в рамках одной и той же популяции, которая впервые успешно колонизировала этот регион, однако в настоящее время распространение гаплогрупп C и D сильно различается. Различные субклады гаплогруппы C встречаются с высокой частотой среди аборигенов Австралии, полинезийцев, микронезийцев, монголов, западных бурят, калмыков, казахов и аборигенных народов Дальнего востока России, а также с умеренной частотой среди корейцев и маньчжуров. С другой стороны, гаплогруппа D встречается с высокой частотой только среди тибетцев, японцев (особенно айнов) и жителей Андаманских островов, однако не обнаружена ни в Индии, ни среди индейцев или аборигенов Океании.
По данным компании YFull гаплогруппа C1-F3393/Z1426 разделилась на ветви C1a-CTS11043 и C1b-F1370 47 300 лет назад[12]. Разделение гаплогрупп C1-F3393/Z1426 и С2-M217 на субклады происходило в Сибири до Последнего ледникового максимума.
По данным генетиков линии C1b-F1370 (ISOGG 2018) из Южной Азии, представленными субкладами C1b1a1-M356 из Индии и C1b1a2-AM00847/AMM008/B65 из Борнео (Калимантан), разошлись с линиями, представленными субкладами C1b2b-M347/P309 из Австралии и C1b2a-M38 из Новой Гвинеи, 54 тыс. л. н. (95%-й доверительный интервал: 47,8—61,4 тыс. л. н.). C1b2b аборигенов Австралии и C1b2a папуасов разошлись 50,1 тыс. л. н. (95%-й доверительный интервал: 44,3—56,9 тыс. л. н.)[13]. По данным компании YFull гаплогруппа C1b-F1370 разделилась на ветви C1b1-AM00694/K281 и C1b2-B477/Z31885 47 200 лет назад[14].
В период от 15,3 до 14,3 тыс. л. н. от гаплогруппы C2-M217 образовалось 8 субветвей. В Берингию мигрировали 3 субклада, а Америки достиг один субклад C2b1a1a-P39 (ISOGG 2018) или C2a1a1a-P39 (ISOGG 2019)[15][16]. C2a1a1-F3918 включает в себя субклад C2a1a1a-P39, который с высокой частотой обнаруживался в выборках некоторых коренных североамериканских популяций, и субклад C2a1a1b-FGC28881, который в настоящее время встречается с различной (но в целом довольно низкой) частотой по всей евразийской степи от провинций Хэйлунцзян и Цзянсу на востоке до турецкой провинции Гиресун, Подляского воеводства и Южно-Чешского края на западе.
Предполагается, что в Америку гаплогруппа C попала около 8—6 тысяч лет назад вместе с носителями языков на-дене и разошлась вдоль северо-западного побережья Северной Америки.
Палеогенетика
- C1 (C-F3393) определили у образцов ВВ7-240 и CC7-335 возрастом более 45 тыс. л. н. из пещеры Бачо Киро (Болгария)[17]
- C1b-F1370 определили у образца Костёнки 14 со стоянки Маркина Гора (костёнковские стоянки), жившего около 37 тыс. лет назад (верхний палеолит)[18][19] (субклад C1b1-K281>Z33130>Z33130*[20])
- C1a определили у образца GoyetQ116-1 (ориньякская культура) из пещеры Гойе в Бельгии (около 35 тыс. л. н.)
- C1a2 определили у четырёх образцов со стоянки Сунгирь (34,65—33,7 тыс. л. н.)[21]
- C1a2 определили у образца Pavlov 1 из Чехии (около 30 тыс. л. н.)[18]
- C1a2 определили у образца Vestonice 16 из Дольни-Вестонице (граветт Чехии, около 30 тыс. л. н.)[22]
- C1a2 определили у анатолийского охотника-собирателя ZBC из местечка Pınarbaşı в турецкой провинции Конья, жившего 13 642 — 13 073 лет до нашей эры[23].
- C2a1a1-F3918>C-F3918*[24] (ISOGG 2019) определили у представителя селенгинской культуры UKY001 (14 054—13 771 л. н.) из Усть-Кяхты в Бурятии[25]
- C1a1a определили у представителя азильской культуры BAL003 (12 830—10 990 лет до настоящего времени) из Бальма де Гилания или Бальма де ла Гвинеу (Balma Guianya) в Пиренеях (испанская провинция Льейда)[26]
- Гаплогруппу C определили у анатолийского фермера ZMOJ из местечка Бонджуклу (Boncuklu), жившего 8300—7800 лет до нашей эры (Анатолия)[23]
- C2c-P53.1 определили у представителя мезолитической культуры Лепенски-Вир[27]
- C2b (ISOGG 2018) или C2a>C-L1373 (ISOGG 2019) определили у образца CP19 (Lapa01 (Burial 1)) из бразильского Лапа до Санто (10 160—9600 лет до настоящего времени)[28]
- C1a2 определили образцов 2779 F.265 и 5357 F.576 (7035—6680 лет до н. э.) из Чатал-Хююка (Турция)[29]
- C1a2-V20 определили у неолитического образца из местонахождения Barçın в Турции с датировкой 8500—8200 лет назад[30]
- C1a2b определена у образца LEPE48 (8012—7867 лет до настоящего времени) из Лепенски-Вира в Сербии и у представителя культуры линейно-ленточной керамики (LBK) Dil16 (7235—6998 лет до н. в.) из Dillingen-Steinheim в Германии[31]
- Гаплогруппу C определили у образца La368 культуры Хоа-Бинь (около 7888 лет назад) из Лаоса[32]
- У темнокожих и голубоглазых мезолитических братьев из пещеры Ла Бранья-Аринтеро (La Braña 1, La Braña 2), найденных в провинции Леон (северо-запад Испании) определена Y-хромосомная гаплогруппа C1a2 (ранее обозначалась как C6)[33][34]
- Гаплогруппу C определили неолитического обитателя Малык-Преславеца (Болгария)[22]
- C2b-L1373 (ISOGG 2018) или C2a2-M217>MPB373/L1373 (ISOGG 2019) определён у образца NEO239 (7545 лет до настоящего времени) из пещеры Чёртовы Ворота в Приморье[35]
- C1a2 определили у представителя культуры кардиальной керамики из хорватской пещеры Земуника (Zemunica) близ Биско, жившего около 7,5 тыс. лет назад[22]
- C1a2 определили у раннеолитического (~5460—5220 лет до н. э.) образца UZZ61 из Сицилии[36]
- C1a2 (ранее обозначалась как C6) определили у представителя культуры линейно-ленточной керамики из венгерского местонахождения Apc-Berekalja I, жившего примерно 4950—5300 лет назад, и у представителя восточной культуры линейно-ленточной керамики (Alföld Linear Pottery) из венгерского местонахождения Kompolt-Kigyósér, жившего примерно 4990—5210 лет назад[37]
- C2b-F1396 определили у представителей культуры Бойсмана (~5000 лет до н. э.) в Приморском края[38][39]
- Гаплогруппу C определили у представителя культуры неолита/халколита Хуншань (Китай)[40][41]
- C2b1a1b-F3830 (ISOGG 2015) определили у образца TL1а (Жужаньский каганат)[42]
- C2b1b/F845 и C2a1a1b1a/F3830 определили у 6 кочевников с Монгольского плато, живших в период с позднего неолита до династии Юань (~3500—700 л. н.)[43]
- C1b1a2b определили у образца AMA001 (2258 ± 30 л. н.) с острова Моротай (Aru Manara, провинция Северное Малуку, Индонезия)[44]
- C2a2-M217>MPB373/L1373>F1756 (ISOGG 2019) определили у образца bla001 (1344—1270 л. н., VII век) со стоянки Октябрьское (Oktyabr'skoe (burial 2)) в Амурской области[45]
- C2a1a1b1b-Y10442/etc (xZ31688,Z31693) определили у раннего аварского военного лидера FGDper4 (Early Avar military leader, 620—660 гг.)[46]
- C2b1a3a1a-Y4580 (с отрицательной Y4541) определили у члена армии хана Джучи (образец DA28), похороненного в золотоордынском погребении XIII—XIV века в горах Улытау (Казахстан)[47]
Известные представители гаплогруппы C
См. также
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 GHIJK | |||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
Примечания
- Генофонд казахов. Web (22 января 2017).
- Copyright 2015 by ISOGG. ISOGG 2015 Y-DNA Haplogroup C. www.isogg.org. Дата обращения: 30 сентября 2015.
- В. Н. Харьков, К. В. Хамина, О. Ф. Медведева, К. В. Симонова, Е. Р. Ерёмина, В. А. Степанов. ГЕНОФОНД БУРЯТ: КЛИНАЛЬНАЯ ИЗМЕНЧИВОСТЬ И ТЕРРИТОРИАЛЬНАЯ ПОДРАЗДЕЛЕННОСТЬ ПО МАРКЕРАМ Y-ХРОМОСОМЫ // ГЕНЕТИКА, 2014, том 50, № 2, стр. 203–213.
- Жаксылык Сабитов, Батыр Даулет. Потомки Адигине и Тагая среди киргизов // The Russian Journal of Genetic Genealogy. — 2013. — Т. 5, № 1. — С. 48-51. — ISSN 1920-2997.
- Wang, CC; Li, H. Inferring human history in East Asia from Y chromosomes (англ.) // Investig Genet : journal. — 2013. — Vol. 4. — P. 11. — doi:10.1186/2041-2223-4-11. — PMID 23731529.
- C YTree v5.02 at 11 february 2017
- Pille Hallast, Anastasia Agdzhoyan, Oleg Balanovsky, Yali Xue, Chris Tyler-Smith. Early replacement of West Eurasian male Y chromosomes from the east, 2019
- Juhasz et al. New clustering methods for population comparison on paternal lineages. Molecular genetics and genomics 290 (2) : 767-784, (2014)
- Tatiana M. Karafet et al. Siberian genetic diversity reveals complex origins of the Samoyedic‐speaking populations, 08 November 2018
- C-V20 YTree
- Monika Karmin et al. A recent bottleneck of Y chromosome diversity coincides with a global change in culture
- C-F3393 YTree
- Anders Bergström et al. Deep roots for aboriginal australian Y chromosomes, 2016 (mutation rate of 0,76×10−9 per site per year)
- C-F1370 YTree
- Lan-Hai Wei et al. Paternal origin of Paleo-Indians in Siberia: insights from Y-chromosome sequences
- По пути в Америку остановка в Берингии была не слишком долгой
- Mateja Hajdinjak et al. Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry, 07 April 2021
- Qiaomei Fu et al. The genetic history of Ice Age Europe, 2016
- Dienekes’ anthropology blog: Genome of Kostenki-14, an Upper Paleolithic European (Seguin-Orlando, Korneliussen, Sikora, et al. 2014). dienekes.blogspot.ru. Дата обращения: 30 сентября 2015.
- C-Z33130 YTree
- Sikora M. et al. Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers, Science 10.1126/science.aao1807 (2017).
- Iain Mathieson et al. The genomic history of Southeastern Europe, 2017
- Michal Feldman et al. Late Pleistocene human genome suggests a local origin for the first farmers of central Anatolia, 19 March 2019
- C-F3918 YTree
- He Yu et al. Paleolithic to Bronze Age Siberians Reveal Connections with First Americans and across Eurasia, May 20, 2020
- Vanessa Villalba-Mouco et al. Survival of Late Pleistocene Hunter-Gatherer Ancestry in the Iberian Peninsula, March 14, 2019
- Zuzana Hofmanova. Palaeogenomic and biostatistical analysis of ancient DNA data from Mesolithic and Neolithic skeletal remains, 2017
- Cosimo Posth et al. Reconstructing the Deep population history of Central and South America, 2018
- Reyhan Yaka et al. Variable kinship patterns in Neolithic Anatolia revealed by ancient genomes, April 14, 2021
- Mathieson I. et al. (2015), Eight thousand years of natural selection in Europe
- The mixed genetic origin of the first farmers of Europe (2020)
- Hugh McColl et al. Ancient genomics reveals four prehistoric migration waves into Southeast Asia, 2018
- Olalde I., Mallick S., Patterson N. et al. The genomic history of the Iberian Peninsula over the past 8000 years // Science. 2019 Mar 15
- Brown-skinned, blue-eyed, Y-haplogroup C-bearing European hunter-gatherer from Spain (Olalde et al. 2014)
- Martin Sikora et al.The population history of northeastern Siberia since the Pleistocene
- Marieke Sophia van de Loosdrecht et al. Genomic and dietary transitions during the Mesolithic and Early Neolithic in Sicily, 2020
- Genome flux and stasis in a five millennium transect of European prehistory
- Chuan-Chao Wang et al. Genomic Insights into the Formation of Human Populations in East Asia, 2021
- Chuan-Chao Wang et al. The Genomic Formation of Human Populations in East Asia, 2020
- Y Chromosome analysis of prehistoric human populations in the West Liao River Valley, Northeast China
- Geographic locations and the Y-chromosome haplogroup distribution of prehistoric populations in this study
- Jiawei Li et al. The genome of an ancient Rouran individual reveals an important paternal lineage in the Donghu population, 2018
- Jiawei Li, Dawei Cai, Ye Zhang, Hong Zhu, Hui Zhou. Ancient DNA reveals two paternal lineages C2a1a1b1a/F3830 and C2b1b/F845 in past nomadic peoples distributed on the Mongolian Plateau, 14 May 2020
- Sandra Oliveira et al. Ancient genomes from the last three millennia support multiple human dispersals into Wallacea, 2021
- Gülşah Merve Kılınç et al. Human population dynamics and Yersinia pestis in ancient northeast Asia, 06 Jan 2021
- Zoltan Maroti et al. Whole genome analysis sheds light on the genetic origin of Huns, Avars and conquering Hungarians, 2021
- Peter de Barros Damgaard et al. 137 ancient human genomes from across the Eurasian steppes, 2018
- Число отцов-основателей современных жителей Азии пополнилось
- Елена Клещенко Всемирная история в четырёх буквах // «Химия и жизнь» № 4, 2015
- Relating Clans Ao and Aisin Gioro from northeast China by whole Y-chromosome sequencing
- Соломин А. В. Князья Гантимуровы.- М., 2016.- 2-е изд.- Стр. 12
Ссылки
- ISOGG 2016 Y-DNA Haplogroup C
- [www.familytreedna.com/public/C3 C3-M217, FTDNA]
- [www.familytreedna.com/(0gyohdakp43sweeab4z0bt55)/public/C/C3%20haplogroup/index.aspx?fixed_columns=on C & C3 Y-Haplogroup project at FTDNA]
- Spread of Haplogroup C, from National Geographic
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
