Гаплогруппа A (Y-ДНК)
В человеческой генетике гаплогруппа A — это Y-хромосомная гаплогруппа. Это гаплогруппа всех существующих сегодня людей, от неё происходят все другие. Как полагают, гаплогруппа A была у Y-хромосомного Адама. Она является самой старой из гаплогрупп Y-ДНК. У Sapiens sapiens буквой А обозначают все гаплогруппы, которые ответвились между появлением Y-хромосомного Адама (Y-MRCA, по оценкам, ок. 250 тыс. л. н.) и мутациями определяющими гаплогруппу BT (оценивается примерно в 88 тыс. л. н. (TMRCA)).
| Гаплогруппа A | |
|---|---|
 | |
| Тип | Y-ДНК |
| Время появления | 75000—120000 |
| Место появления | Африка |
| Предковая группа | Адам |
| Субклады | A00, A0, A1, A2, A3 |
| Мутации-маркеры | M91 |
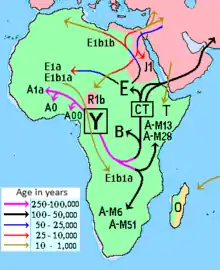
Наиболее базальными субкладами гаплогруппы А по возрасту дивергенции являются: «А00», «А0», «А1» (также «A1a-Т») и «А2-Т». Гаплогруппа BT, предковая для всех неафриканских гаплогрупп, является субкладой гаплогруппы «А2-Т» (в нотации 2011 года).
Происхождение
Предполагается, что в далёком прошлом человеческого рода гаплотип А, вид гаплогруппы А, возник как один из двух известных рядов поколений Y-хромосомного Адама, который является предком по мужской линии всех живущих мужчин. Большинством исследователей считается, что этот предок жил в Африке до того как люди расселились из Африки более 60 000 лет назад. По многим оценкам этот предок жил около 75 000 лет назад. Некоторые полагают, что это приблизительное время, когда люди покинули Африку[1]. По оценке 2014 года возникновение линии гаплогруппы A00 датируется примерно 208300 годом до н. э. (95 % 260 200—163 900 гг. до н. э.)[2].


Предположения о раннем распространении
Согласно многим предположениям в отношении происхождения Y-хромосомной гаплогруппы А, оно связано с генетической основой в виде южноафриканских охотников и собирателей или происходящих от них людей, говорящих на койсанских щёлкающих языках[3]. Предполагается, либо что эти люди возникли в Южной Африке, либо что они возникли к северу и востоку от южноафриканских пустынь (Намиб), и распространились на юг. При изучении митохондриальных геномов в группе койсанов Бехар (Behar) и др. в 2008 году обнаружили, что древние геномы койсанов ограничивались митохондриальными гаплогруппами L0d и L0k, по их оценке ранней митохондриальной ветви L0k, возникших приблизительно 144 000 лет назад, что составляет около 3/4 времени появления ближайшего митохондриального родственника (митохондриальной Евы)[4]. Оценки возраста митохондриальной Евы сильно различаются, но самые новые, основанные на последнем возможном исходе анатомически современных людей из Африки, свидетельствуют о времени приблизительно 108 000 лет назад[5]. Из этого следует, что ответвление относится ко времени приблизительно 80 000 лет назад, но такой срок предполагает очень небольшое население в Африке и противоречит определённому согласно исследованиям Х-хромосом и аутосом наименьшему размеру популяции. Более предпочтительные оценки, относящиеся к очищающему отбору, относят ответвление митохондриальной линии L0k к периоду между 120 000 и 160 000 лет назад[6][7].
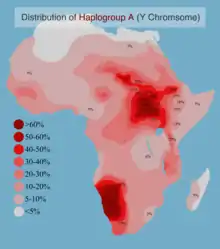
| Африка | ||
| . | Исследуемая популяция (народ) | Частота (в %) |
| [8] | Бушмены (Намибия, Тсумкве) | 66 |
| [8] | Нама (Намибия) | 64 |
| [9] | Динка (Судан) | 62 |
| [9] | Шиллук (Судан) | 53 |
| [9] | Нуба (Судан) | 46 |
| [10][11] | Эфиопские евреи | 41 |
| [12] | койсаны | 44 |
| [8][10] | Кунг/Секеле | ~40 |
| [9] | Маба (Судан) | 35 |
| [9] | Нуэр (Судан) | 33 |
| [9] | Фор (Судан) | 31 |
| [8] | Масаи (Кения) | 27 |
| [13] | Масалит (Судан) | 19 |
| [8][13] | Амхара (Эфиопия) | ~16 |
| [12] | Эфиопы | 14 |
| [14] | Банту (Кения) | 14 |
| [8] | Мандара (Камерун) | 14 |
| [9] | Хауса (Судан) | 13 |
| [10] | Кхое (Южная Африка) | 12 |
| [10] | Фульбе (Камерун) | 12 |
| [8] | Дамара (Намибия) | 11 |
| [13] | Оромо (Эфиопия) | 10 |
| [8] | Носители Южно-семитского (Эфиопия) | 10 |
| [14] | Арабы (Египет) | 3 |
Y-хромосомные гаплогруппы A и B могли распространиться во время этого немногочисленного распространения популяции людей. Из других исследований следует, что оцениваемый размер воспроизводившейся популяции мужчин составляет около половины от размера популяции женщин, поэтому возраст ближайшего мужского родственника должен быть меньше возраста митохондриального родственника. Таким образом, удалённость точек ветвления линий A/BT в Y-хромосомной гаплогруппе А не обязательно должна быть такой же большой, как от митохондриальной линии L0k до основной точки L0/L1.
Гипотезы смежных популяций
Археологические свидетельства предполагают, что в доисторическое время популяции, являвшиеся предшественниками койсанов, могли обитать на территориях, ограниченных с севера Эфиопией и Суданом. Расчленённое распространение гаплогруппы А может свидетельствовать о том, что она когда-то была распространена среди охотников и собирателей в Африке, но многие её субклады могли быть замещены потомками с гаплогруппой Е, которые пришли с распространением сельского хозяйства во время миграции банту[8]. Очаги гаплогрупп A и B могли сохраниться в отдельных популяциях, таких как эфиопская, нило-суданская и пигмейская, которые были изолированы от переселяющихся земледельцев банту.
Дополнительные подтверждения древней связи между койсанами и жителями Северной Африки относятся к митохондриальным исследованиям ДНК. К жителям Эфиопии и койсанам относятся древнейшие клады как митохондриальной филогенетики людей, так и Y-хромосомной. Клады митохондриальной гаплогруппы L0 были обнаружены среди койсанов и жителей Восточной Африки (например, танзанийцев, кенийцев и эфиопов), но являются редкими или отсутствуют в других популяциях Африки[15].
Распространение
Гаплогруппа A1b1a1a-M6 распространена среди койсанов, таких как бушмены, и можно предполагать, что это их наследственная гаплогруппа. К примеру, Knight и др. (2003) сообщают о 12—44 % гаплогруппы A1b1a1a-M6 среди различных племён койсанов. Удивительно, что эта гаплогруппа не была обнаружена у народа хадза из Танзании, хотя их традиционно рассматривали, как древний остаток койсанцев из-за присутствия щёлкающих согласных в их языке.
В 2001 году гаплогруппу A обнаружили у 10,3 % исследуемых оромо и 14,6 % исследуемых амхарцев в Эфиопии[16]. Часто (41 %) встречается у эфиопских евреев (Cruciani и др. 2002), у народа бантус в Кении (14 %, Luis и др. 2004), ираку в Танзании (17 %, Knight и др. 2003), и фульбе в Камеруне (12 %, Cruciani и др. 2002). Наибольшая распространённость гаплогруппы A в Восточной Африке, однако, была обнаружена среди суданского населения: 42,5 % (Underhill и др. 2000).
Y-хромосомный Адам
По результатам сравнения Y-хромосомы неандертальца из пещеры Эль-Сидрон и африканца с Y-хромосомной гаплогруппой A00, время появления Y-хромосомного Адама оценили в 275 тыс. лет назад (95 % доверительный интервал: 304—245 тыс. лет назад)[17].
A00
Все обнаруженные носители гаплогруппы A00 живут в Африке (Западный Камерун) в составе племён Nkongho-Mbo (en:Mbo people (Cameroon)) и Bangwa/Nweh (fr:Bangoua (peuple)) или являются афроамериканцами. Mendez и др. объявили об открытии ранее неизвестной гаплогруппы в 2013 году и предложили обозначение А00. Предполагаемый возраст образования новой гаплогруппы они тогда оценили в 270 тыс. л. н., что было старше текущих оценок возраста общего предка анатомически современных людей по мужской линии.
Впервые эта ранее неизвестная гаплогруппа была обнаружена в 2012 году в Y-хромосоме афроамериканца из Южной Каролины, который представил его ДНК для коммерческого генеалогического анализа (так как афроамериканца звали Альберт Перри (Albert Perry), то гаплотип также известен как «Perry’s Y»). Позже нашли гаплогруппу A00 в генетических данных одиннадцати мужчин камерунского племени Nkongho-Mbo, а затем ещё у восьми в 2015 году. Дальнейшие исследования в 2015 году показали, что самая высокая концентрация А00 выявлена в племени Bangwa/Nweh и что они находятся в отдельной подгруппе по отношению к носителям A00 из племени Nkongho-Mbo. Один человек с A00 не относится ни к одной из подгрупп[18].
Y-хромосомная гаплогруппа A00 обнаружена у одного образца 2/SEII из пещеры Шум Лака в Камеруне[19]. 2/SEII и современных носителей A00 жилл ок. 31 тыс. лет до настоящего времени (95% доверительный интервал: 37000—25000 лет до н. в.)[20].
A0-T
Гаплогруппа A0-T сформировалась 235,9 тыс. л. н., последний общий предок современных носителей A1b1 жил 161,3 тыс. л. н. (по данным компании YFull)[21].
A1
Подгруппа A1 очень редка. Сформировалась 161,3 тыс. л. н., время жизни общего предка современных известных носителей (TMRCA) A1 — 133,4 тыс. лет назад (дата определена по снипам компанией YFull[24]).
A1a
В 2007 году семь человек из Йоркшира в Англии, обладатели редкой йоркширской фамилии, оказались носителями субклады A1a. Это означало, что у них был общий предок по отцовской линии с XVIII века, но никаких подробностей об африканской родословной не было известно. В дополнение к семи йоркширцам, известно о 25 живущих сейчас носителях подгруппы A1, имеющих западноафриканскую родословную[25], и о нескольких выходцах из Европы (финнах шведского происхождения, норвежцах, немцах, голландцах, бельгийцах).
A1b
Гаплогруппа A1b-P108(xA1b1b2a) определена у кенийского образца I8758 (Naishi Rockshelter, Pastoral_Neolithic (other), 2700—2370 лет до настоящего времени)[26].
A1b1
Гаплогруппа A1b1 сформировалась 130,7 тыс. л. н., последний общий предок современных носителей A1b1 жил 126 тыс. л. н. (по данным комапнии YFull)[27].
Пик распространения приходится на Эфиопию и нилотские народы Восточной Африки субклады A1b1b-М32, найдена также у арабов Персидского залива и Ливана, армян, в Казахстане, на Русской равнине и Британских островах. За пределы Восточной Африки субклад A1b1b-М32 распространился как минорная ветвь в миграциях народов-носителей субклады E1b1b1-М35, со стороны Африканского Рога, в том числе и путём работорговли. Гаплогруппа A1b1b-M32 сформировалась 126 тыс. л. н., последний общий предок современных носителей A1b1b-M32 жил 55,1 тыс. л. н. (по данным компании YFull)[28].
Субклад A1b1b2a (снип М51) определён у 28,4 % бушменов Южной Африки, A1b1a1a1b (P28) — у 8,7 %, A1b1a1a1a (M114) — у 4,4 %, A1b1a1a (М14) — у 2,7 %[29].
Подгруппа A1b1b2b-M13/PF1374 (ранее A3b2), найденная в Восточной Африке (в Эфиопии и среди нилотских народов Уганды, Кении и Танзании), отличается от найденных у койсанов, и только отдалённо связана с ними (фактически только одна из многих подгрупп в пределах гаплогруппы A1b1). Также субклад A1b1b2b-M13 обнаружена у шотландцев и американцев шотландского происхождения, у арабов Персидского залива и севернее, вплоть до Чехии, России, Украины и Казахстана. Разделение гаплогруппы A1b1b2-L427 на субклады A1b1b2a-M51 и A1b1b2b-М13 произошло 45,2 тыс. лет назад[30].
Гаплогруппа A1b1b2-L427 определена у кенийского образца I8804 (Keringet Cave (GrJg4), Pastoral_Neolithic (other), 2700—2360 л. н.), A1b1b2b-M13 определена у кенийского образца I8919 (Naivasha Burial Site, Pastoral_Neolithic (other), 2340—2160 л. н.)[26].
BT
Примечания
- Основываясь на археологических определения возраста из Восточной Индии, определениях возраста анатомически современных людей в Лиучжяне[уточнить] и определениях возраста людей очень сходных с анатомически современными людьми в Леванте это время оценивается около 93 000 лет назад.
- The ‘extremely ancient’ chromosome that isn’t: a forensic bioinformatic investigation of Albert Perry’s X-degenerate portion of the Y chromosome
- Гондер (Gonder) и др. в 2007 году предположили, что митохондриальная Ева появилась в Танзанийском регионе. Однако, позднее Тишкофф (Tishkoff) расширил область её происхождения до региона границы Намибии и Анголы, ближе к Атлантическому побережью
- Behar, DM; Villems, R; Villems; Soodyall, H; Soodyall; Blue-Smith, J; Blue-smith; Pereira, L; Pereira; Metspalu, E; Metspalu; Scozzari, R; Scozzari; Makkan, H; Makkan; Tzur, S; Tzur; D. The dawn of human matrilineal diversity (англ.) // American Journal of Human Genetics : journal. — 2008. — May (vol. 82, no. 5). — P. 1130—1140. — doi:10.1016/j.ajhg.2008.04.002. — PMID 18439549.
- Endicott, P; Ho, SY; Metspalu, M & Stringer, C (2009), Evaluating the mitochondrial timescale of human evolution, Trends Ecol. Evol. (Amst.) Т. 24 (9): 515–21, PMID 19682765, DOI 10.1016/j.tree.2009.04.006
- Soares, P; Ermini, L; Ermini; Thomson, N; Thomson; Mormina, M; Mormina; Rito, T; Rito; Röhl, A; Röhl; Salas, A; Salas; Oppenheimer, S; Oppenheimer; Macaulay, V; MacAulay; M. B. Correcting for purifying selection: an improved human mitochondrial molecular clock (англ.) // American Journal of Human Genetics : journal. — 2009. — June (vol. 84, no. 6). — P. 740—759. — doi:10.1016/j.ajhg.2009.05.001. — PMID 19500773.
- Gonder, MK; Mortensen, HM; Mortensen; Reed, FA; Reed; de Sousa, A; De Sousa; Tishkoff, SA; Tishkoff. Whole-mtDNA genome sequence analysis of ancient African lineages (англ.) // Mol. Biol. Evol : journal. — 2007. — December (vol. 24, no. 3). — P. 757—768. — doi:10.1093/molbev/msl209. — PMID 17194802.
- Elizabeth T Wood, Daryn A Stover, Christopher Ehret et al., "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes, " European Journal of Human Genetics (2005) 13, 867—876. (cf. Appendix A: Y Chromosome Haplotype Frequencies)
- 28/53 (Dinka, Nuer, and Shilluk), Hassan H. Y., Underhill P. A., Cavalli-Sforza L. L., Ibrahim M. E. Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history (англ.) // Am. J. Phys. Anthropol. : journal. — 2008. — November (vol. 137, no. 3). — P. 316—323. — doi:10.1002/ajpa.20876. — PMID 18618658. Архивировано 4 марта 2009 года. Архивированная копия (недоступная ссылка). Дата обращения: 21 января 2010. Архивировано 4 марта 2009 года.
- Fulvio Cruciani, Piero Santolamazza, Peidong Shen et al., "A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes," American Journal of Human Genetics 70:1197–1214, 2002
- Peidong Shen, Tal Lavi, Toomas Kivisild et al., "Reconstruction of Patrilineages and Matrilineages of Samaritans and Other Israeli Populations From Y-Chromosome and Mitochondrial DNA Sequence Variation," Human Mutation 24:248-260 (2004).
- Underhill P. A., Shen P., Lin A. A., et al. Y chromosome sequence variation and the history of human populations (англ.) // Nat. Genet. : journal. — 2000. — November (vol. 26, no. 3). — P. 358—361. — doi:10.1038/81685. — PMID 11062480.
- Semino O., Santachiara-Benerecetti A. S., Falaschi F., Cavalli-Sforza L. L., Underhill P. A. Ethiopians and Khoisan share the deepest clades of the human Y-chromosome phylogeny (англ.) // Am. J. Hum. Genet. : journal. — 2002. — January (vol. 70, no. 1). — P. 265—268. — doi:10.1086/338306. — PMID 11719903.
- J. R. Luis, D. J. Rowold, M. Regueiro et al., "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations," American Journal of Human Genetics 74:532–544, 2004.
- Gonder et al. Whole-mtDNA Genome Sequence Analysis of Ancient African Lineages (англ.) : journal. — 2007.
- Ornella Semino, A. Silvana Santachiara-Benerecetti, Francesco Falaschi, L. Luca Cavalli-Sforza, Peter A. Underhill. Ethiopians and Khoisan Share the Deepest Clades of the Human Y-Chromosome Phylogeny. 2001 (недоступная ссылка)
- Fernando L. Mendez et al. The American Journal of Human Genetics: The Divergence of Neandertal and Modern Human Y Chromosomes, 2016.
- Which of Cameroon’s peoples have members of haplogroup A00? // How far we’ve come. Dec 01, 2015
- Lipson Mark et al. Ancient Human DNA from Shum Laka (Cameroon) in the Context of African Population History // SAA 2019
- Ancient West African foragers in the context of African population history (PDF), 2020
- A0-T YTree
- A0 YTree
- Y-DNA Haplogroup A and its Subclades
- A1 YTree v4.09
- en:Turi E. King, en:Emma J. Parkin, en:Geoff Swinfield, en:Fulvio Cruciani, en:Rosaria Scozzari, en:Alexandra Rosa, en:Si-Keun Lim, en:Yali Xue, en:Chris Tyler-Smith & en:Mark A. Jobling. Africans in Yorkshire? The deepest-rooting clade of the Y phylogeny within an English genealogy (англ.) // en:European Journal of Human Genetics : journal. — 2007. — March (vol. 15, no. 3). — P. 288—283. — doi:10.1038/sj.ejhg.5201771. — PMID 17245408. News article: Yorkshire clan linked to Africa, en:BBC News (1 февраля 2000). Дата обращения 27 января 2007.
- Mary E. Prendergast et al. Ancient DNA reveals a multistep spread of the first herders into sub-Saharan Africa (Table S6. (separate file) Y chromosome haplogroups and derived SNPs), 2019
- A1b1 YTree
- A-M32 YTree
- Naidoo T. et al. Development of a single base extension method to resolve Y chromosome haplogroups in sub-Saharan African populations Investig Genet. 2010.
- A-M13 YTree
Ссылки
- Y-DNA Haplogroup Tree 2017 (Root (Y-Adam) // A00 / A0-T)
- Y-DNA Haplogroup A and its Subclades — 2017
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 GHIJK | |||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
