Гаплогруппа L (Y-ДНК)
Гаплогруппа L (M20) — Y-хромосомная гаплогруппа. Определяется SNP (снипами) M11, M20, M61 и M185. Является вторичным потомком гаплогруппы K и первичной ветвью гаплогруппы LT(K1), гаплогруппа L в настоящее время имеет альтернативное филогенетическое имя K1a и является параллельной и наиболее родственной Y-хромосомной гаплогруппе T (K1b).
| Гаплогруппа L-M20 | |
|---|---|
 | |
| Тип | Y-ДНК |
| Время появления |
25-30 тыс. лет назад 45500-39700 по yfull.com |
| Место появления | Южная Азия, Западная Азия, Памир |
| Время до БОП | 24000 лет назад |
| Предковая группа | K > LT |
| Сестринские группы | T |
| Субклады |
L1a1-M27,M76 (L1,L1a до 2015 года) L1a2-M357,L1307 (L3, L1c до 2015 года), Гаплогруппа L3 (Y-ДНК) L1a3-M2533* носитель ветви выявлен в 2020-ом году L1b-M317 (L2 до 2015 года) L2-L595 (открыт в 2014 году) |
Особенности
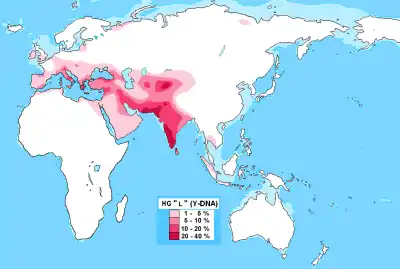
Одна из самых редких. Мировая концентрация гаплогруппы приходится на Южную Азию, но составляет лишь 11,2 % от всего населения региона[1] (281 из 2504 исследованных людей). В южной Азии отсутствует максимальное разнообразие субкладов (ветвей) L, оно приходится на Западную Азию и Кавказ, достигая пика в полосе, которая охватывает западный край Ирана, северный Ирак и северную Сирию.[2][3][4][5]
Частота L не превышает 10-20 % практически ни у одного народа мира с населением более 1 млн человек, что делает это мужское потомство одним из самых редких на земле[6][1]. Исключения: 85,2 миллионный народ джат из северо-западной Индии (36.8 %)[7] и двухмиллионная языковая община саураштра с юга Индии (26.1 %)[8]. В последнем случае данные могут быть искажены, так как исследовалось всего 46 человек.
На крайнюю редкость L указывают мужские генетические древа компаний Yfull и FTDNA (лидер по количеству Y-ДНК тестов). Количество выявленного разнообразия ветвей L составляет 339 вариантов у FTDNA и 349 вариантов у Yfull, это около 1 % от количества ветвей, выявленных у всех остальных гаплогрупп (данные на 30 июня 2021[9][10][11]).
Одна из самых «горных». Вне южной Индии и южного Пакистана пики концентраций L приходятся на популяции совершенно разных горных регионов планеты:
- Долины, соседствующие с Гималаями и Тибетом. Тянутся цепью от Памира Таджикистана на севере до Ладакха Индии на юге. Преобладает субклад L1a2 с наличием L1a1.
- Cеверный Кавказ. Чечня. L1a2 с небольшим наличием L1b.
- Черноморский регион Турции, вдоль которого пролегают Понтийские горы, достигая пика 3428 метров на восточной окраине. Субклад L1b.
- Два самых горных региона Италии — Тироль на севере и Л’Акуила на юге (исторический Самний). Субклад L1b.
Часть этносов, проживающих в данных регионах, сближаются гипотетической дене/сино-кавказской языковой семьёй, предложенной A. Старостиным в 1980-х годах, ещё до начала генетических исследований человечества.
Наибольшая частота L-M20 приходится на Южную Азию, охватывая соседние Памирские горы Центральной Азии. Пики концентраций L обнаруживаются у популяций Пакистана, Индии и Таджикистана, проживающих в высокогорных долинах в близости от Гималаев и Тибета:
- Брокпа\Минаро — 62,7 % L1a2\L3[12], население 48000 человек. Субэтнос народа шина. Проживают в Ладакхе — на самом высоком плато Индии, населяемом тибетоязычными и индоевропеоязычными людьми. Генетическое исследование вышло в свет в конце 2019 года.
- Джаты — 36,8 % L[7], население 82,5 млн человек. Занимают обширные области северной Индии и Пакистана.
- Калаша\Калаши — 25 % L[13], население 4000 человек. Крайний север Пакистана.
- Бурушо\Буриши — 16 % L[13], население 87000 человек. Крайний север Пакистана.
- Шугнанцы Памира — 16 % L[14], население 90000-115000 человек. Территория Таджикистана к северу от Пакистана.
- Ишкашимцы Памира — 12 % L[14], население 1500 человек. Территория Таджикистана к северу от Пакистана.
- Высокие концентрации L также отмечаются в Южной Индии (17-19 % от региона в целом),[15] у пуштунов северного Афганистана (25 %)[16] и в Пакистане (11,6 % в целом[13], 28 % на западе страны, включая Белуджистан[1]). Встречается в Таджикистане, Узбекистане, Турции, а также более низкими частотами в Иране и на всём Ближнем Востоке. В течение тысячелетий этот мужской ген присутствовал очень низкими частотами на Кавказе и в Европе.
Открытия
Гаплогруппа L и три её субклада были обнаружены в 2006 году учёным S. Sengupta[17]
В 2014 году был обнаружен редкий субклад — L2 (L-L595) который встречается только в Европе и западной Азии и отстоит от субкладов L1 генетической дистанцией в 23000 лет до общего предка[18][19].
В 2020 году у этнического курда из северной Сирии был выявлен редкий промежуточный субклад L1a3 (L-M2533*), входящий в ветвь L1a (L-M2481), но не имеющий родство с ветвями L1a1 (L-M27) и L1a2 (L-M357, L-L1307) ближе, чем на 17000 лет до общего предка.[20]
Филогенетическое древо с возрастами субкладов, и географией их высоких концентраций
За основу взяты древа ISOGG (2018) и Yfull (v7.06.00).[21][22] Приведена распространенная в научных работах номенклатура геномутаций.
L M20, M11, M61, M185 (гаплогруппа зародилась 42600 л. назад и распалась 23000 л. наз. на два субклада L1 и L2)
• L1 M22 (зародился 24000 лет наз. и распался 18200 лет наз. на два субклада — L1a и L1b)
• • L1a M2481 (зародился 18200 лет наз. и распался 17000 лет наз. на L1a1, L1a2 и L1a3)
• • • L1a1 (L1, L1a до 2015) M27, M76
Юг Индии в абсолютном большинстве связан с дравидоязычными народами. Исключение — 82.5 миллионый народ джат северной Индии, 36,8 % мужчин этого народа являются носителями L, однако точный процент L1a1 неизвестен.[7]
• • • L1a2 (L3, L1c до 2015) M357, L1307 (Возраст до общего предка -
Пакистан: пики на юге и в высокогорных долинах на крайнем севере.
Памир Таджикистана.
Индия: Очень высокая концентрация 62,7 % на крайнем севере в Ладакхе, у этнической группы брокпа\минаро, которая составляет около половины от народа шина. В целом L1a2 составляет около 1 % на всю страну, рассеян по всей южной Индии крайне низкими частотами.[12]
Чечня (Северный Кавказ, Россия). Уже более 1000 человек числятся в таблице чеченского национального ДНК проекта «Сhechen-noahcho dna project», созданного на базе американской лаборатории FTDNA. Его данные показывают, что от 7 до 14 % чеченских мужчин являются носителями L1a2, однако никто это число официально не уточнял. ДНК-исследования кавказцев, проведённые О. Балановским в 1998—2009, годах происходили в Чечне во времена боевых действий и не охватили все регионы республики.
• • • L1a3 (M2533*)
Северная Сирия, Турция. Крайне редок, на 2021 год известно лишь два его носителя. Выявлен у курда из северной Сирии в 2020-ом году.
• • L1b (L2 до 2015) M317, L655
Северная Турция и восточная часть Альп.
• L2-L595 обнаружен в 2014-ом году
Европа и западная Азия. Крайне редок. Не имеет родства с другими ветвями ближе 23000 лет до общего предка.
Происхождение и общая география субкладов
L-M2645*/M20/PF5570* сформировалась около 42,6 тыс. лет назад, последний общий предок современных носителей гаплогруппы L жил 24,2 тыс. лет назад[23]. L-M20 происходит от Y-хромосомной гаплогруппы LT[24][25], которая происходит от гаплогруппы K-M9[26][25]. По словам доктора Спенсера Уэлса гаплогруппа L зародилась в сложной системе Памирских гор на территории нынешнего Таджикистана, и мигрировала на территорию Пакистана и Индии 30 000 лет назад[27][28][29]. Тем не менее, большинство других иcследований указывают на западноазиатское происхождение гаплогруппы L и связывают её экспансию в долину Инда с неолитическими фермерами[30][31][32][8][33][34]. На сей день абсолютное большинство исследованных останков неолитических фермеров показывают субклад G2a2-L30 гаплогруппы G (в прошлом G2a3), только очень малая часть относится к потомству гаплогруппы J2[35][36][37][38][39]. Палео-ДНК центральной и южной Азии эпохи неолита всё ещё не исследована, однако научная работа Singh 2016, посвящённая изучению современного распространения J2 в южной Азии, отмечает, что география её распространения в мире сходится c расположением агрокультурных центров[40].
Учёные McElreavy и Quintana-Murci, описывая Индскую (Хараппскую) цивилизацию, утверждают, что «[т]олько Y-хромосомная гаплогруппа L-M20 имеет высокую среднюю частоту — 14 % в Пакистане и поэтому отличается от всех других гаплогрупп по своему частотному распределению. L-M20 также найдена, хотя и более низкими частотами, в соседних странах, таких как Индия, Таджикистан, Узбекистан и Россия. Как распределение частот, так и предполагаемое время экспансии (~ 7000 лет назад) этой генетической линии позволяют предположить, что её распространение в долине Инда может быть связано с экспансией местных фермерских групп в период неолита».[41]
Обширное исследование Sengupta 2006, в котором была изучена Y-ДНК 728 индийцев и 176 пакистанцев, выявило три субклада гаплогруппы L: L1-M76,M27 (ныне L1a1), L2-M317 (ныне L1b) и L3-M357 (ныне L1a2, L-1307 по yfull.com).[17] Почти все индийские носители гаплогруппы L показали субклад L1a1, причем L1a2, отстоящий от L1a1 генетической дистанцией в 17000 лет[18], оказался очень редким (0,4 % 3/728).[17][42] В Пакистане же субклад L1a2 составил 86 % от всей L и достиг средней частоты 6,8 % в целом.[43] L1a1 выявлена с частотой 7,5 % в Индии и 5,1 % в Пакистане, пик её разнообразия приходится на Махараштру — прибрежную западную часть Индии. Субклад L1b был найден только в Пакистане — 1,14 % (2 из 176).
Имеющиеся результаты исследований Y-ДНК мировых популяций, множество из которых приведены в данной статье, показывают редкость распространения субклада L1b-M317 в южной и центральной Азии, в то же время данный субклад имеет большинство носителей L-M20 в западной Азии и Европе. У субкладов L1a1 и L1a2 всё предстоит наоборот, их частота резко падает вне территорий южной Азии и исторической Бактрии. Из ненаучных данных это подтверждает и ДНК-проект «Haplogroup L» на базе коммерческой лаборатории FTDNA (около 700000 Y-хромосомных ДНК-тестов[44]), состоящий из 795 носителей L-M20 со всего мира. (7.06.2019)
Высокая частота субклада L1a2-M357 в Европе и на Кавказе обнаруживается только у нахоязычных народов Кавказа, в большей части у чеченцев — более 10 % (110 из 922 человек на 26.06.2019 г.) в проекте «Chechen-Noahcho DNA project», созданном на базе американской лаборатории FTDNA. Исследование Balanovsky 2011, в котором участвовало 1525 мужчин из 14 популяций Кавказа, выявило L1a2 только у чеченцев и ингушей.[45]
Палеогенетика
- L1a1-M27 определили у представителей халколита Армении из пещеры Арени (около 6 тыс. л. н.)[39][46]. Субклад имели все трое индивидов мужского пола из пещеры Арени. У образца I1632 субклад L1a1-M27>Y31961* определён компанией Yfull[47]. В пещере были обнаружены самые древние кожаная обувь (5500 л. н.)[48][49][50], юбка[51], винодельня[52]
- Cубклад L2-L595 определён у троих представителей поздней майкопской культуры. Образец SIJ002.A0101 из могильника Синюха в Адыгее, образцы MK5001, MK5004 — могильник Марьинская 5[53] у границ Ставропольского края и КБР. Возраст останков 5141—5173 лет назад[54]
- L2-L595 найден у образца ALA084 из турецкого Алалаха (2006—1777 лет до н. э. или 3556±25 лет до настоящего времени)[55]
- L1a определили в БМАК (Бактрийско-Маргианский археологический комплекс) на крайнем юге Узбекистана. Образец i4285 из Саппали-тепе имеет возраст 1873—1661 лет до н. э. с калибровкой. Возраст i5604 из могильника Бустан — 1880—1697 лет до н. э.[46]
- L1a обнаружен у пяти образцов найденных в Пакистане, возраста от 1300 до 800 лет до нашей эры[46]
- L-M20 определили у филистимлянина найденных в Ашкелоне (Израиль)[56]
- L1a1-Y31961* определили у гунна из Тянь-Шаня (на территории Киргизстана), образец DA85 (ERS2374343) имеет возраст 1781 л. н. без калибровки[57]. Ветвь определена компанией Yfull[47]
- L1a2 (L-M357) определили у представителя отрарской культуры KNT005 из могильника Коныр-Тобе (256—381 гг.) на юге Казахстана[58]
- Исследование ДНК скелета гуннского периода из Музея естественной истории (г. Будапешт), датированного средней третью V века, показало, что он имел Y-хромосомную гаплогруппу L[59]
- L2-L595>L-BY106184* определили у образца VK355 с острова Эланд (лен Кальмар, Швеция). Останки датируются 847 годом (± 65 лет)[60]. Субклад L-Y31961* определён компанией Yfull[61]
- L1a1b[62] определили у средневекового образца VK538 (XI—XIII века) с кладбища в 3,5 км от города Троя на юге Италии в Апулии, которое использовалось во времена завоевания Италии норманнами[60]
Распространение
Южная Азия
Обширное исследование (Mahal 2018) по южной Азии в целом, состоящее из 2504 образцов мужской ДНК из Индии, Бангладеш, и Пакистана — выявило 11,2 % L-M20 (281 из 2504)[1].
Индия
Встречается чаще среди дравидийских каст (около 17-19 %), и реже среди индоарийских (около 5-6 %).[17] Может достигать 68 % в некоторых племенах и кастах Карнатаки,[63] 38 % в некоторых кастах Гуджарата,[63] 48 % в некоторых кастах Тамилнада, и 12 % по общей частоте в Пенджабе.[17][63][42] Более ранние исследования (например, Wells 2001) сообщают об очень высокой частоте (приближающейся к 80 %) гаплогруппы L-M20 в Тамилнаде, по-видимому, из-за экстраполяции данных полученных из выборки 84 калларов — тамило-говорящей высокой касты правителей Тамилнада, среди которых 40 (приблизительно 48 %) показали мутацию M20 определяющую гаплогруппу L. Присутствие гаплогруппы L-M20 редко встречается среди племенных групп (около 5,6-7 %).[8][17][32]
68 % L-M20 было обнаружено у племени Корова из Карнатаки, 38 % в касте Барвад (Bharwad) из округа Джунагадх в Гуджарате, 21 % в касте Чаран из округа Джунагадх в Гуджарате, 17 % в племени Каре Воккал (Kare Vokkal) из Уттара Каннада в Карнатаке. Также встречается с низкой частотой в других популяциях областей Джунагадх и Уттара Каннада. L-M20 имеет большая часть мужчин (36,8 %) среди Джатов в Северной Индии, и встречается у 16,33 % Гуджаров в Джамму и Кашмире.[28][64] Обнаружена у 18,6 % Конканских (Читпаванских) брахманов[42] и 15 % маратхов Махараштры. L-M20 также обнаружена у 32,35 % Воккалигов и у 17,82 % Лингаятов Карнатаки.[65] L-M20 обнаружена на уровне 20,7 % среди амбалакараров, 16,7 % среди Айенгаров, и 17,2 % среди Айеров (брахманская каста) Тамилнада.[17] L-M11 встречается частотой 8-16 % среди индийских евреев.[66] 2 % L-M11 обнаружено у Сидди — народа Африканского происхождения.[63] В целом гаплогруппа L-M20 в настоящее время присутствует в индийской популяции с общей частотой около 7-15 %.[67][8][17][32]
Пакистан
Наибольшая концентрация L-M20 в Пакистане тянется вдоль реки Инд, где 3300-1300 лет до н. э. процветала Индская (Хараппская) цивилизация. Наибольшая частота и разнообразие L1a2-M357 обнаружены в Белуджистане — 28 %,[30] другая точка концентрации L1a2-M357 находится в горных изолированных долинах на севере страны, там он обнаружен у 16,5 % Буришей и 25 % Калашей.[13] У Пакистанских Пуштунов численность которых составляет около 32 млн чел.[68] процент M357 составил около 7 % (Firasat 2007),[13] исследование Lee 2014 (270 человек) показало результат — 5,9 %[69] В среднем M357 имеет распространение 11,6 % по всему Пакистану.[13]
Афганистан
Пуштуны являются самым многочисленным народом Афганистана (около 14 млн человек)[70]. Исследование (Lacau 2012) ДНК 190 мужчин показало, что гаплогруппа L-M20 с общей частотой 9,5 % является второй по численности мужской линией среди них, субклад L1a2-M357 составил 7,5 % из 9,5 %.[16] Исследование демонстрирует существенную разницу распределения L-M20 по обеим сторонам хребта Гиндукуш — L выявлена у 25 % северных афганских пуштунов и 4,8 % южных. В частности, L1a2-M357 составляет большинство как на севере (20,5 %), так и на юге (4,1 %). В более раннем исследовании, в котором участвовало меньшее количество образцов, сообщалось, что L1a2-M357 составляет 12,24 % от общего количества афганских пуштунов.[71]
Таджики — второй по численности народ в Афганистане (ок. 9-11 млн человек),[72][73] сконцентрированный в северных провинциях страны. Выявлено 6,34 % L1a (9 из 142). Из них 5 L1a2 и 4 L1a1. Провинции — Балх (6 чел.), Бадахшан (2 чел.), Саманган (1 чел.)[74]
У узбеков (ок. 4 млн чел[75]) выявлено 9,52 % L (12/126). Из них L1a1 — 6 чел, L1a2 — 5 чел. L1b — 1 чел. — северныe провинции Дзаузджан, Сари-Пуль, Балх (1 чел.)[74]
Хазарейцы 2,97 % L-M20 (3/101) — 2 человека в пров. Балх и 1 человек в пров. Бамиан.[74]
Западная Азия и Кавказ
| Популяции | Распространение | Источники |
|---|---|---|
| Турция | 57 % — деревня Афшар, 12 % (10/83) — Черноморский регион, 6,6 % (7/106) — Турки из Турции. Так-же — 4,2 % (1/523 L-M349 и 21/523 L-M11(xM27, M349)) | Cinnioğlu 2004, Gokcumen 2010 Karafet 2016 |
| Иран | Самое обширное исследование Grugni 2012 (938 мужчин,15 этносов, 14 провинций) показало результат — 5 % L-M20 (L1a1 — 1,8 % L1a2 — 1,5 % L1b — 1,5 % L* — 0,2 %)[76]
Результаты других исследований, часть которых вошла в Grugni 2012: 54,9 % (42/71) — у зороастрийских священников среди Парсов 22,2 % L1b и L1c в южном Иране (2/9) 8 % — 16 % L2-L595, L1a, L1b и L1c у Курдов Курдистана (2-4/25) 9,1 % L-M20 (7/77) у персов восточного Ирана 3,4 % L-M76 (4/117) и 2,6 % L-M317 (3/117), в целом 6,0 % (7/117) L-M20 в Южном Иране 3,0 % (1/33) L-M357 в северном Иране 4,2 % L1c-M357 Азербайджанцев в провинции восточный Азербайджан (1/21) 4,8 % L1a и L1b у Персов из Исфахана (2/42) |
Regueiro 2006, Grugni 2012, Cristofaro 2013[74], Malyarchuk 2013, Lopez 2017 |
| Сирия | 51.0 % (33/65) Сирийцев в Ракке, 31,0 % у восточных Сирийцев. В абсолютном большинстве L1b-M317 | El-Sibai 2009 |
| Саудовская Аравия | 15.6 % (4/32 из L-M76 и 1/32 of L-317) 1,91 % (2/157=1.27 % L-M76 и 1/157=0.64 % L-M357) | Karafet 2016 and AbuAmero 2009 |
| Курды | Турция
3,2 % в юго-восточной Турции (Flores 2005) |
Flores 2005 |
| Ирак | 3.1 % (2/64) L-M22 (Sanchez 2005)
Северный Ирак (Dogan 2017) Язиды — 10,09 % (11/109 все L1b-m317) Арабы — 3,70 % (4/108 L1a-m27 — 3 чел. L1b-m317 −1 чел) Курды — 2,73 % (3/110 L1a-M27 — 2 чел. L1c — 1 чел.) Туркмены 2,73 % (3/110 L1b-m317 — 2 L1a-m27 — 1) Ассирийцы — 1,09 % (1/92 L1a-m27) |
Sanchez 2005 Dogan 2017[2] |
| Оман | 1 % L-M11 | Luis 2004 |
| Катар | 2.8 % (2/72 L-M76) | Cadenas 2008 |
| Арабы ОАЭ | 3.0 % (4/164 L-M76 и 1/164 L-M357) | Cadenas 2008 |
| Армяне | от 1,63 % (12/734) до 4,3 % (2/47) | Weale 2001 and Wells 2001 |
| Лазы | 41.7 % (15/36) L1b-M317 | Balanovsky 2017 |
| Грузины | 20 % (2/10) в Гали, 14,3 % (2/14) в Чохатаури, 12,5 % (2/16) в Мартвили, 11,8 % (2/17) в Абаше, 11,1 % (2/18) в Багдати, 10 % (1/10) в Гардабани, 9,1 % (1/11) в Адигени, 6,9 % (2/29) в Омало, 5,9 % (1/17) в Гурджаани, 5,9 % (1/17) в Лентехи. Также — 1,5 % (1/66) L-M357(xPK3) , 1,6 % (1/63) L-M11 | Battaglia 2008, Semino 2000 and Tarkhnishvili 2014 |
| Дагестан | 9.5 % L1b-M317 (4/42) у Аварцев, 8,3 % (2/24) L1a2-M357 у Татов, 11,76 % (2/17) L1b-M317 у горских Евреев 3,7 % (1/27) Чамалинцев, | Yunusbaev 2006, Caciagli 2009, Karafet 2016 |
| Чеченцы и Ингуши | Чеченцы
Из ненаучных данных: Более 10 % L1a2-M357 и около 1 % L1b-M317 (110 из 922 и 9 из 922 человек на 26.06. 2019 г.) в ДНК-проекте «Chechen-noahcho dna project» созданном на базе американской лаборатории FTDNA. 14 % (14/100) L1a2-M357 у чеченцев из Ауховского района Дагестана (Balanovsky 2011) Ингуши Из ненаучных данных: 7 % L1a2-M357 (20/277) в ДНК-проекте «Ingush DNA project» Все чеченцы и ингуши носители L1a2-M357 происходят от ветви L-Y6266 возрастом 3300 лет, которая является западно-Азиатской и отстоит от средне и южно Азиатских ветвей генетической дистанцией в 6000-7000 лет.[77] |
Chechen-Noahcho DNA project
Ingush DNA project Balanovsky 2011[45] |
В небольшой выборке, взятой у друзов Израиля, гаплогруппа L была обнаружена у 7 человек из 20 (35 %). С другой стороны, исследования, проведенные на более широких выборках, показали, что мутация L-M20 в среднем составляет 5 % среди друзов Израиля[78], 4 % среди друзов Ливана[79]. Кроме этого, она не была обнаружена вообще в выборке из 59 друзов Сирии.
Центральная Азия
| Популяции | Распространение | Источник |
|---|---|---|
| Казахи | 1 % (3/300) L1c-m357 — 2 чел. в Жамбыльской и Восточно-Казахстанской областях. L1b-m317 — 1 чел. в Акмолинской обл. | Zhabagin 2018 |
| Таджики | Таджикистан
7,74 % (13/168) Шугнан — 7\44 Ишкашим — 3\25 Душанбе — 2\16 Худжанд — 1\22 (Wells 2001). 22,5 % 9\40 — из них L1a2 — 4 (Malyarchuk 2013) Северный Афганистан 6,34 % L1a (9 из 142). Из них 5 L1a2 и 4 L1a1. Провинции — Балх (6 чел.), Бадахшан (2 чел.), Саманган (1 чел.) |
Malyarchuk 2013, Wells 2001[14], Cristofaro 2013[3] |
| Узбеки | Узбекистан
L-M20 3,0 % 11/366 (Wells 2001). L-M20 5,12 % (11/215) 5-Хорезм 4-Фергана 2-Ташкент, из них L1a2-M357 3 — 2-Фергана 1-Xорезм (Balanovska 2017) Северный Афганистан 9,52 % L (12/126). Из них L1a1 — 6 чел, L1a2 — 5 чел. L1b — 1 чел. — северныe провинции Балх, Дзаузджан, Сари-Пуль. |
Wells 2001, Cristofaro 2013, Zhabagin-Balanovska 2017[80] |
| Уйгуры | 16.7 % (1/6) L1c-M357 в Киргизстане | Cristofaro 2013 |
| Памирцы | 16 % (7/44) Шугнанцы , 12 % 3/25 Ишкашимцы, 0/30 Бартанги | Wells 2001 |
| Хазарейцы | 2.97 % L-M20 (3/101) — 2 человека в пров. Балх и 1 человек в пров. Бамиан | Cristofaro 2013 |
| Ягнобцы | 9.7 % (3/31) | Wells 2001 |
| Бухарские Арабы | 9.5 % (4/42) | Wells 2001 |
| Дунгане | 8.3 % (1/12) L1c в Киргизстане | Cristofaro 2013 |
| Уйгуры (Lopliks) | 7.8 % (5/64) L-M357 деревня Qarchugha, округ Lopnur, Синцзян | Liu 2018[81] |
| Каракалпаки | 4.5 % (2/44) | Wells 2001 |
| Уйгуры | 4.4 % (3/68) | Karafet 2001 and Hammer 2005[ |
| Tуркмены | Афганистан
4,1 % (3/74) L1a-M27 в Джаузджан |
Cristofaro 2013 |
| Челканы | 4.0 % (1/25) | Dulik 2012 and Dulik 2012 |
| Киргизы | 2.7 % (1/37) L1c сев. запад и 2,5 % (1/40) L1a в центре Киргизстана | Cristofaro 2013 |
| Казанские Татары | 2.6 % (1/38) | Wells 2001 |
| Хуэй | 1.9 % (1/54) | Karafet 2001 |
| Башкиры | 0.64 % (3/471) | Lobov 2009 |
Европа
| Популяция | Распространение | Источник |
|---|---|---|
| Валь-ди-Фасса (Fascia). Италия | 19.2 % L-M20 | Valentina Coia 2013[82] |
| Валь-ди-Нон (Nonstal). Италия | 10 % L-M20 | F. di Giacomo 2003[83] |
| Южный Тироль, Италия | 8.9 % Ладино-язычных из Валь Бадия 8,3 % Валь Бадия, 2,9 % долина Пустерталь, 2,2 % германоязычных из Валь Бадия, 2 % германоязычных в верхнем Финшгау, 1,9 % германоязычных в нижнем Финшгау и 1,7 % Итальяноязычных в Больцано | Pichler 2006[84] and Thomas 2008[85] |
| Самний, Италия | 10 % Аквиланцев L-M20 | Alessio Boattini 2013[86] |
| Виченца, Италия | 10 % Венецианцев L-M20 | Alessio Boattini 2013 |
| Восточный Тироль, Австрия | L-M20 1,9 % у Тирольцев в регионе Б (Isel, Lower Drau, Defereggen, Virgen, Kals valley) | H.Niederstätter 2012 |
| Северный Тироль, Австрия | L-M20 0,8 % Ройтте | D.Erhart 2012 |
| Архангельская область, Россия | 5.9 % Русских L1c-M357 (Нет результатов исследования в открытом виде, требуется уточнение информации) | Hongyang Xu 2014 |
| Португалия | 5.0 % в Коимбра | Beleza 2006 |
| Болгария | 3.9 % у Болгар | Karafet 2016 |
| Фландрия, Бельгия | L1a*: 3,17 % в Мехелен 2,4 % в Тюрнхаут and 1,3 % в Де-Кемпен. L1b*: 0,74 % на западе и востоке | Larmuseau 2010 and Larmuseau 2011 |
| Гипускоа, Испания | L1b 1,7 % | Young 2011 |
См. также
Примечания
- David G. Mahal, Ianis G. Matsoukas. The Geographic Origins of Ethnic Groups in the Indian Subcontinent: Exploring Ancient Footprints with Y-DNA Haplogroups (англ.) // Frontiers in Genetics. — 2018. — Т. 9. — ISSN 1664-8021. — doi:10.3389/fgene.2018.00004.
- Serkan Dogan, Cemal Gurkan, Mustafa Dogan, Hasan Emin Balkaya, Ramazan Tunc. A glimpse at the intricate mosaic of ethnicities from Mesopotamia: Paternal lineages of the Northern Iraqi Arabs, Kurds, Syriacs, Turkmens and Yazidis (англ.) // PLOS ONE. — 2017-03-11. — Vol. 12, iss. 11. — P. e0187408. — ISSN 1932-6203. — doi:10.1371/journal.pone.0187408.
- Julie Di Cristofaro, Erwan Pennarun, Stéphane Mazières, Natalie M. Myres, Alice A. Lin. Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge // PLoS ONE. — 2013-10-18. — Т. 8, вып. 10. — ISSN 1932-6203. — doi:10.1371/journal.pone.0076748.
- L YTree. www.yfull.com. Дата обращения: 1 июля 2021.
- Oleg Balanovsky, Khadizhat Dibirova, Anna Dybo, Oleg Mudrak, Svetlana Frolova. Parallel Evolution of Genes and Languages in the Caucasus Region // Molecular biology and evolution. — 2011-10. — Т. 28, вып. 10. — С. 2905–2920. — ISSN 0737-4038. — doi:10.1093/molbev/msr126.
- List of Y-chromosome haplogroups in populations of the world (англ.) // Wikipedia. — 2021-02-08.
- David G. Mahal, Ianis G. Matsoukas. Y-STR Haplogroup Diversity in the Jat Population Reveals Several Different Ancient Origins // Frontiers in Genetics. — 2017-09-20. — Т. 8. — ISSN 1664-8021. — doi:10.3389/fgene.2017.00121.
- Richard Cordaux, Robert Aunger, Gillian Bentley, Ivane Nasidze, S.M. Sirajuddin. Independent Origins of Indian Caste and Tribal Paternal Lineages (англ.) // Current Biology. — Cell Press, 2004-2. — Vol. 14, iss. 3. — P. 231—235. — doi:10.1016/j.cub.2004.01.024.
- www.familytreedna.com/public/y-dna-haplotree
- ftdna haplotree - Поиск в Google. www.google.com. Дата обращения: 25 июня 2021.
- YFull | Статистика YTree. www.yfull.com. Дата обращения: 29 июня 2021.
- Adhikarla Syama. Origin and identity of the Brokpa of Dah-Hanu, Himalayas – an NRY-HG L1a2 (M357) legacy (англ.) ?.
- Sadaf Firasat, Shagufta Khaliq, Aisha Mohyuddin, Myrto Papaioannou, Chris Tyler-Smith. Y-chromosomal evidence for a limited Greek contribution to the Pathan population of Pakistan (англ.) // European Journal of Human Genetics. — 2007-1. — Vol. 15, iss. 1. — P. 121—126. — ISSN 1476-5438 1018-4813, 1476-5438. — doi:10.1038/sj.ejhg.5201726.
- R. Spencer Wells, Nadira Yuldasheva, Ruslan Ruzibakiev, Peter A. Underhill, Irina Evseeva. The Eurasian Heartland: A continental perspective on Y-chromosome diversity // Proceedings of the National Academy of Sciences of the United States of America. — 2001-08-28. — Т. 98, вып. 18. — С. 10244–10249. — ISSN 0027-8424. — doi:10.1073/pnas.171305098.
- Sanghamitra Sengupta, Lev A. Zhivotovsky, Roy King, S. Q. Mehdi, Christopher A. Edmonds. Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists (англ.) // The American Journal of Human Genetics. — 2006-2. — Vol. 78, iss. 2. — P. 202—221. — doi:10.1086/499411.
- Harlette Lacau, Tenzin Gayden, Maria Regueiro, Shilpa Chennakrishnaiah, Areej Bukhari. Afghanistan from a Y-chromosome perspective (англ.) // European Journal of Human Genetics. — 2012-10. — Vol. 20, iss. 10. — P. 1063—1070. — ISSN 1476-5438 1018-4813, 1476-5438. — doi:10.1038/ejhg.2012.59.
- Sanghamitra Sengupta, Lev A. Zhivotovsky, Roy King, S.Q. Mehdi, Christopher A. Edmonds. Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists (англ.) // The American Journal of Human Genetics. — 2006-2. — Vol. 78, iss. 2. — P. 202—221. — doi:10.1086/499411.
- Copyright 2014 by ISOGG. ISOGG 2014 Y-DNA Haplogroup L (англ.). isogg.org. Дата обращения: 4 июля 2019.
- L YTree. www.yfull.com. Дата обращения: 4 июля 2019.
- L-M2481 YTree. www.yfull.com. Дата обращения: 27 июня 2021.
- ISOGG 2018 Y-DNA Haplogroup L. isogg.org. Дата обращения: 7 июля 2019.
- YTree. www.yfull.com. Дата обращения: 7 июля 2019.
- L YTree
- ISOGG 2018 Y-DNA Haplogroup Tree Trunk. isogg.org. Дата обращения: 3 июля 2019.
- J. Chiaroni, P. A. Underhill, L. L. Cavalli-Sforza. Y chromosome diversity, human expansion, drift, and cultural evolution (англ.) // Proceedings of the National Academy of Sciences. — National Academy of Sciences, 2009-12-01. — Vol. 106, iss. 48. — P. 20174—20179. — ISSN 1091-6490 0027-8424, 1091-6490. — doi:10.1073/pnas.0910803106.
- ISOGG 2018 Y-DNA Haplogroup K. isogg.org. Дата обращения: 3 июля 2019.
- Wells, Spencer, 1969-. Deep ancestry : inside the Genographic project. — Washington D. C.: National Geographic, 2007. — 247 pages с. — ISBN 9781426201189, 1426201184.
- David G. Mahal, Ianis G. Matsoukas. Y-STR Haplogroup Diversity in the Jat Population Reveals Several Different Ancient Origins // Frontiers in Genetics. — 2017-09-20. — Т. 8. — ISSN 1664-8021. — doi:10.3389/fgene.2017.00121.
- Spencer Wells. The Journey of Man. A Genetic Odyssey. — India: New Delhi: Penguin Books, 2003. — С. 167.
- Raheel Qamar, Qasim Ayub, Aisha Mohyuddin, Agnar Helgason, Kehkashan Mazhar. Y-Chromosomal DNA Variation in Pakistan (англ.) // The American Journal of Human Genetics. — 2002-5. — Vol. 70, iss. 5. — P. 1107—1124. — doi:10.1086/339929.
- Zhongming Zhao, Faisal Khan, Minal Borkar, Rene Herrera, Suraksha Agrawal. Presence of three different paternal lineages among North Indians: A study of 560 Y chromosomes (англ.) // Annals of Human Biology. — 2009-1. — Vol. 36, iss. 1. — P. 46—59. — ISSN 1464-5033 0301-4460, 1464-5033. — doi:10.1080/03014460802558522.
- Ismail Thanseem, Kumarasamy Thangaraj, Gyaneshwer Chaubey, Vijay Kumar Singh, Lakkakula VKS Bhaskar. Genetic affinities among the lower castes and tribal groups of India: inference from Y chromosome and mitochondrial DNA // BMC Genetics. — 2006-08-07. — Т. 7, вып. 1. — С. 42. — ISSN 1471-2156. — doi:10.1186/1471-2156-7-42.
- K. Mcelreavey, L. Quintana-Murci. A population genetics perspective of the Indus Valley through uniparentally-inherited markers (англ.) // Annals of Human Biology. — 2005-3. — Vol. 32, iss. 2. — P. 154—162. — ISSN 1464-5033 0301-4460, 1464-5033. — doi:10.1080/03014460500076223.
- Kumarasamy Thangaraj, B. Prathap Naidu, Federica Crivellaro, Rakesh Tamang, Shashank Upadhyay. The Influence of Natural Barriers in Shaping the Genetic Structure of Maharashtra Populations (англ.) // PLOS One / Richard Cordaux. — Public Library of Science, 2010-12-20. — Vol. 5, iss. 12. — P. e15283. — ISSN 1932-6203. — doi:10.1371/journal.pone.0015283.
- F. Di Giacomo, F. Luca, L. O. Popa, N. Akar, N. Anagnou. Y chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe (англ.) // Human Genetics. — 2004-10. — Vol. 115, iss. 5. — P. 357—371. — ISSN 1432-1203 0340-6717, 1432-1203. — doi:10.1007/s00439-004-1168-9.
- Zuzana Hofmanová, Susanne Kreutzer, Garrett Hellenthal, Christian Sell, Yoan Diekmann. Early farmers from across Europe directly descended from Neolithic Aegeans (англ.) // Proceedings of the National Academy of Sciences. — National Academy of Sciences, 2016-06-21. — Vol. 113, iss. 25. — P. 6886—6891. — ISSN 1091-6490 0027-8424, 1091-6490. — doi:10.1073/pnas.1523951113.
- Gülşah Merve Kılınç, Ayça Omrak, Füsun Özer, Torsten Günther, Ali Metin Büyükkarakaya. The Demographic Development of the First Farmers in Anatolia (англ.) // Current Biology. — Cell Press, 2016-10. — Vol. 26, iss. 19. — P. 2659—2666. — doi:10.1016/j.cub.2016.07.057.
- Iain Mathieson, Iosif Lazaridis, Nadin Rohland, Swapan Mallick, Nick Patterson. Eight thousand years of natural selection in Europe // bioRxiv. — 2015-10-10. — doi:10.1101/016477.
- Iosif Lazaridis et al. Genomic insights into the origin of farming in the ancient Near East, 2016 (The genetic structure of the world’s first farmers, bioRxiv)
- Dissecting the influence of Neolithic demic diffusion on Indian Y-chromosome pool through J2-M172 haplogroup. Singh 2016 https://www.nature.com/articles/srep19157#discussion
- K. McElreavy and L. Quintana-Murci (2005) http://ibrarian.net/navon/paper/A_population_genetics_perspective_of_the_Indus_Va.pdf?paperid=6978605 Архивная копия от 16 января 2017 на Wayback Machine
- T. Kivisild, S. Rootsi, M. Metspalu, S. Mastana, K. Kaldma. The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations (англ.) // The American Journal of Human Genetics. — 2003-2. — Vol. 72, iss. 2. — P. 313—332. — doi:10.1086/346068.
- Sengupta 2006 стр. 219 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1380230
- ftdna statistics - Поиск в Google. www.google.com. Дата обращения: 5 июля 2019.
- Oleg Balanovsky, Khadizhat Dibirova, Anna Dybo, Oleg Mudrak, Svetlana Frolova. Parallel Evolution of Genes and Languages in the Caucasus Region (англ.) // Molecular Biology and Evolution. — Oxford University Press, 2011-10-01. — Vol. 28, iss. 10. — P. 2905—2920. — ISSN 0737-4038 1537-1719, 0737-4038. — doi:10.1093/molbev/msr126.
- Vagheesh M Narasimhan, Nick J Patterson, Priya Moorjani, Iosif Lazaridis, Lipson Mark. The Genomic Formation of South and Central Asia // bioRxiv. — 2018-03-31. — doi:10.1101/292581.
- L-M27 YTree. www.yfull.com. Дата обращения: 4 июля 2019.
- Ron Pinhasi, Boris Gasparian, Gregory Areshian, Diana Zardaryan, Alexia Smith. First Direct Evidence of Chalcolithic Footwear from the Near Eastern Highlands (англ.) // PLOS One / Michael D. Petraglia. — Public Library of Science, 2010-06-09. — Vol. 5, iss. 6. — P. e10984. — ISSN 1932-6203. — doi:10.1371/journal.pone.0010984.
- Вячеслав Локацкий. Люди медного века щеголяли в кожаных ботинках. Правда.Ру (15 июня 2010). Дата обращения: 4 июля 2019.
- Armenian cave yields what may be world's oldest leather shoe - CNN.com (англ.). www.cnn.com. Дата обращения: 4 июля 2019.
- 5,900-year-old women’s skirt discovered in Armenian cave (англ.). news.am. Дата обращения: 4 июля 2019.
- Earliest Known Winery Found in Armenian Cave. National Geographic News (12 января 2011). Дата обращения: 4 июля 2019.
- Могильник Марьинская 5 — Россия как археологическое пространство. Дата обращения: 4 июля 2019.
- Chuan-Chao Wang, Sabine Reinhold, Alexey Kalmykov, Antje Wissgott, Guido Brandt. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions (англ.) // Nature Communications. — Nature Publishing Group, 2019-12. — Vol. 10, iss. 1. — ISSN 2041-1723. — doi:10.1038/s41467-018-08220-8.
- Eirini Skourtanioti et al. Genomic History of Neolithic to Bronze Age Anatolia, Northern Levant, and Southern Caucasus, 2020
- Michal Feldman, Daniel M. Master, Raffaela A. Bianco, Marta Burri, Philipp W. Stockhammer. Ancient DNA sheds light on the genetic origins of early Iron Age Philistines (англ.) // Science Advances. — 2019-7. — Vol. 5, iss. 7. — P. eaax0061. — ISSN 2375-2548. — doi:10.1126/sciadv.aax0061.
- Peter de Barros Damgaard, Nina Marchi, Simon Rasmussen, Michaël Peyrot, Gabriel Renaud. 137 ancient human genomes from across the Eurasian steppes (англ.) // Nature. — 2018-5. — Vol. 557, iss. 7705. — P. 369—374. — ISSN 1476-4687 0028-0836, 1476-4687. — doi:10.1038/s41586-018-0094-2.
- Guido Alberto Gnecchi-Ruscone et al. Ancient genomic time transect from the Central Asian Steppe unravels the history of the Scythians, 26 Mar 2021
- (нет достоверного источника, используется статья удалённая из интернета но доступная в архиве) Казахстанский ДНК-проект Архивная копия от 26 ноября 2016 на Wayback Machine
- Ashot Margaryan, Daniel John Lawson, Martin Sikora, Fernando Racimo, Simon Rasmussen. Population genomics of the Viking world // bioRxiv. — 2019-07-17. — doi:10.1101/703405.
- L-BY106184 YTree
- L-Z5926 YTree
- Anish M. Shah, Rakesh Tamang, Priya Moorjani, Deepa Selvi Rani, Periyasamy Govindaraj. Indian Siddis: African Descendants with Indian Admixture (англ.) // The American Journal of Human Genetics. — 2011-7. — Vol. 89, iss. 1. — P. 154—161. — doi:10.1016/j.ajhg.2011.05.030.
- Swarkar Sharma, Ekta Rai, Prithviraj Sharma, Mamata Jena, Shweta Singh. The Indian origin of paternal haplogroup R1a1* substantiates the autochthonous origin of Brahmins and the caste system (англ.) // Journal of Human Genetics. — 2009-1. — Vol. 54, iss. 1. — P. 47—55. — ISSN 1435-232X 1434-5161, 1435-232X. — doi:10.1038/jhg.2008.2.
- Shilpa Chennakrishnaiah, Shilpa Chennakrishnaiah, Deetta K. Mills. Analysis of Y-Chromosome Diversity in Lingayat and Vokkaliga Populations of Southern India. — 2011.
- Gyaneshwer Chaubey, Manvendra Singh, Niraj Rai, Mini Kariappa, Kamayani Singh. Genetic affinities of the Jewish populations of India (англ.) // Scientific Reports. — 2016-5. — Vol. 6, iss. 1. — ISSN 2045-2322. — doi:10.1038/srep19166.
- A. Basu. Ethnic India: A Genomic View, With Special Reference to Peopling and Structure (англ.) // Genome Research. — 2003-10-01. — Vol. 13, iss. 10. — P. 2277—2290. — ISSN 1088-9051. — doi:10.1101/gr.1413403.
- South Asia :: Pakistan — The World Factbook - Central Intelligence Agency (недоступная ссылка). www.cia.gov. Дата обращения: 6 июля 2019. Архивировано 24 мая 2020 года.
- Eun Young Lee, Kyoung-Jin Shin, Allah Rakha, Jeong Eun Sim, Myung Jin Park. Analysis of 22 Y chromosomal STR haplotypes and Y haplogroup distribution in Pathans of Pakistan (англ.) // Forensic Science International: Genetics. — 2014-7. — Vol. 11. — P. 111—116. — doi:10.1016/j.fsigen.2014.03.004.
- South Asia :: Afghanistan — The World Factbook - Central Intelligence Agency (недоступная ссылка). www.cia.gov. Дата обращения: 6 июля 2019. Архивировано 20 сентября 2017 года.
- Marc Haber, Daniel E. Platt, Maziar Ashrafian Bonab, Sonia C. Youhanna, David F. Soria-Hernanz. Afghanistan's Ethnic Groups Share a Y-Chromosomal Heritage Structured by Historical Events (англ.) // PLOS One / Manfred Kayser. — Public Library of Science, 2012-03-28. — Vol. 7, iss. 3. — P. e34288. — ISSN 1932-6203. — doi:10.1371/journal.pone.0034288.
- Atlas. — 4th ed. — London: Dorling Kindersley, 2010. — pages cm с. — ISBN 9781405350396, 1405350393.
- Joshua Project. Afghan, Tajik in Afghanistan (англ.). joshuaproject.net. Дата обращения: 6 июля 2019.
- Julie Di Cristofaro, Erwan Pennarun, Stéphane Mazières, Natalie M. Myres, Alice A. Lin. Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge (англ.) // PLOS One / Manfred Kayser. — Public Library of Science, 2013-10-18. — Vol. 8, iss. 10. — P. e76748. — ISSN 1932-6203. — doi:10.1371/journal.pone.0076748.
- South Asia :: Afghanistan — The World Factbook - Central Intelligence Agency. www.cia.gov. Дата обращения: 6 июля 2019.
- Viola Grugni, Vincenza Battaglia, Baharak Hooshiar Kashani, Silvia Parolo, Nadia Al-Zahery. Ancient Migratory Events in the Middle East: New Clues from the Y-Chromosome Variation of Modern Iranians (англ.) // PLOS One / Toomas Kivisild. — Public Library of Science, 2012-07-18. — Vol. 7, iss. 7. — P. e41252. — ISSN 1932-6203. — doi:10.1371/journal.pone.0041252.
- L-L1307 YTree. yfull.com. Дата обращения: 20 июля 2019.
- 12/222 Shlush et al. 2008
- 1/25 Shlush et al. 2008
- Maxat Zhabagin, Elena Balanovska, Zhaxylyk Sabitov, Marina Kuznetsova, Anastasiya Agdzhoyan. The Connection of the Genetic, Cultural and Geographic Landscapes of Transoxiana // Scientific Reports. — 2017-06-08. — Т. 7. — ISSN 2045-2322. — doi:10.1038/s41598-017-03176-z.
- Guanglin He, Zheng Wang, Mengge Wang, Tao Luo, Jing Liu. Forensic ancestry analysis in two Chinese minority populations using massively parallel sequencing of 165 ancestry-informative SNPs (англ.) // ELECTROPHORESIS. — 2018. — Vol. 39, iss. 21. — P. 2732–2742. — ISSN 1522-2683. — doi:10.1002/elps.201800019.
- Valentina Coia, Marco Capocasa, Paolo Anagnostou, Vincenzo Pascali, Francesca Scarnicci. Demographic Histories, Isolation and Social Factors as Determinants of the Genetic Structure of Alpine Linguistic Groups // PLoS ONE. — 2013-12-02. — Т. 8, вып. 12. — ISSN 1932-6203. — doi:10.1371/journal.pone.0081704.
- Clinal patterns of human Y chromosomal diversity in continental Italy and Greece are dominated by drift and founder effects (англ.) // Molecular Phylogenetics and Evolution. — 2003-09-01. — Vol. 28, iss. 3. — P. 387–395. — ISSN 1055-7903. — doi:10.1016/S1055-7903(03)00016-2.
- Irene Pichler, Jakob C. Mueller, Stefan A. Stefanov, Alessandro De Grandi, Claudia Beu Volpato. Genetic structure in contemporary South Tyrolean isolated populations revealed by analysis of Y-chromosome, mtDNA, and Alu polymorphisms. 2006 // Human Biology. — 2009-12. — Т. 81, вып. 5—6. — С. 875–898. — ISSN 1534-6617. — doi:10.3378/027.081.0629.
- Mark G. Thomas, Ian Barnes, Michael E. Weale, Abigail L. Jones, Peter Forster. New genetic evidence supports isolation and drift in the Ladin communities of the South Tyrolean Alps but not an ancient origin in the Middle East (англ.) // European Journal of Human Genetics. — 2008-01. — Vol. 16, iss. 1. — P. 124–134. — ISSN 1476-5438. — doi:10.1038/sj.ejhg.5201906.
- Alessio Boattini, Begoña Martinez-Cruz, Stefania Sarno, Christine Harmant, Antonella Useli. Uniparental Markers in Italy Reveal a Sex-Biased Genetic Structure and Different Historical Strata (англ.) // PLOS ONE. — 2013-05-29. — Vol. 8, iss. 5. — P. e65441. — ISSN 1932-6203. — doi:10.1371/journal.pone.0065441.
Литература
- A. Basu et al.: Ethnic India: A Genomic View, With Special Reference to Peopling and Structure. Genome research, 2003, http://www.genome.org/cgi/doi/10.1101/gr.1413403.
- R. Cordaux et al.: Independent Origins of Indian Caste and Tribal Paternal Lineages. Current Biology, 2004, Vol. 14, p. 231—235
- C. Cinnioğlu et al., "Excavating Y-chromosome haplotype strata in Anatolia, " Hum Genet (2004) 114 : 127—148, http://evolutsioon.ut.ee/publications/Cinnioglu2004.pdf
- R. Qamar et al.: Y-Chromosomal DNA Variation in Pakistan. American Journal of Human Genetics, 2002, p. 1107—1124
- M. Regueiro et al.: "Iran: Tricontinental Nexus for Y-Chromosome Driven Migration, " Human Heredity, 2006, vol. 61, pp. 132-43.
- S. Sahoo et al.: A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios. PNAS 2006, www.pnas.org/cgi/doi/10.1073/pnas.0507714103
- S. Sengupta et al.: Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists. American Journal of Human Genetics, 2006, p. 202—221
- I. Thamseem et al.: Genetic affinities among the lower castes and tribal groups of India: Inference from Y chromosome and mitochondrial DNA. BMC Genetics, 2006, http://www.biomedcentral.com/1471-2156/7/42
- Sadaf Firasat, Shagufta Khaliq, Aisha Mohyuddin, Myrto Papaioannou, Chris Tyler-Smith, Peter A Underhill and Qasim Ayub: Y-chromosomal evidence for a limited Greek contribution to the Pathan population of Pakistan. European Journal of Human Genetics (2007) Vol. 15, p. 121—126. http://www.nature.com/ejhg/journal/v15/n1/full/5201726a.html
- R. Spencer Wells, Nadira Yuldasheva, Ruslan Ruzibakiev, Peter A. Underhill, Irina Evseeva, Jason Blue-Smith, Li Jin, Bing Su, Ramasamy Pitchappan, Sadagopal Shanmugalakshmi, Karuppiah Balakrishnan, Mark Read, Nathaniel M. Pearson, Tatiana Zerjal, Matthew T. Webster, Irakli Zholoshvili, Elena Jamarjashvili, Spartak Gambarov, Behrouz Nikbin, Ashur Dostiev, Ogonazar Aknazarov, Pierre Zalloua, Igor Tsoy, Mikhail Kitaev, Mirsaid Mirrakhimov, Ashir Chariev, and Walter F. Bodmer: «The Eurasian Heartland: A continental perspective on Y-chromosome diversity.» Proceedings of the National Academy of Sciences of the United States of America v.98(18); Aug 28, 2001
Ссылки
- ISOGG 2016 Y-DNA Haplogroup L
- Spread of Haplogroup L, from National Geographic
- [www.familytreedna.com/public/india The India Genealogical Project]
- L YFull YTree v7.07.00
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 GHIJK | |||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
