Гаплогруппа I2 (Y-ДНК)
Гаплогруппа I2 — гаплогруппа Y-хромосомы. Наивысшей концентрации достигает у жителей Боснии-Герцеговины, Сербии и Хорватии.
| Гаплогруппа I2 | |
|---|---|
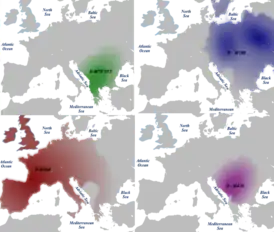 Распространение гаплогруппы I2-M438, а также гаплогрупп E-M78 (V13), R1a-M198, R1b-M269 в Европе Распространение гаплогруппы I2-M438, а также гаплогрупп E-M78 (V13), R1a-M198, R1b-M269 в Европе | |
| Тип | Y-ДНК |
| Время появления | 15 000 до н.э. |
| Место появления | центральная Европа |
| Предковая группа | I |
| Сестринские группы | I1 |
| Субклады | I2a, I2b |
| Мутации-маркеры | M438/P215/S31 |
До 2008 года была известна под названием I1b. Имеет две субклады — I-L460 и I-L1251.
Гаплогруппа I2 происходит от мутации гаплогруппы I, произошедшей у мужчины, жившего ок. 27 500 лет назад. Сама гаплогруппа возникла между 26 000 — 31 000 лет до нашей эры. Последний общий предок современных носителей гаплогруппы I2 жил 21 600 лет назад (даты определены по снипам компанией YFull[1]).
Встречается практически на всей территории современной Европы, максимальной частоты достигает в Динарском нагорье. Мужчины там имеют рекордный для мировой статистики средний рост — 185.6 см.[2][3]
Палеогенетика, происхождение
I2* (I-M438), если говорить о крупных гаплогруппах, значительно представленных в генофонде европейцев, является самой древней гаплогруппой Европейского континента. По всем признакам она также является автохтонной гаплогруппой Европы, возникшей именно в границах данного континента. Такой же автохтонно-европейской гаплогруппой является C1a2 — но данная ветвь очень слабо представлена в генофонде европейцев.
Изначально с Ближнего Востока в Европу пришли люди, нёсшие гаплогруппу IJ — 35 000 лет назад. В процессе миграций населения она постепенно превратилась в гаплогруппу I.
Гаплогруппа I2 (M438/P215/S31) возникла в период позднего палеолита, во время последнего ледникового максимума, между 26 500 — 19 000 лет назад. Определить место или регион в Европе, где произошла гаплогруппа — невозможно, поскольку в тот период все Homo sapiens были охотниками-собирателями — кочевниками, которые постоянно меняли места своих стоянок. Самый старый мужчина-носитель I2 — грот Бишон в Швейцарии, он был представителем азильской культуры, мтДНК у него была U5b1h. К 2016 году проанализировано 15 образцов мужской Y ДНК периода мезолита. Среди этих образцов, один был C1a2 (Испания), один — F (Германия), двое — I (Франция) и шесть образцов — I2 (Люксембург и Швеция), в том числе к I2a1 (P37.2), I2a1a1a (L672), I2a1b (M423) и I2c2 (PF3827), мтДНК образцов — U2e, U4, U5a1, U5a2 и U5b. Четыре образца из России принадлежали Y-хромосомные гаплогруппам — J, R1a1 и R1b1a. Уже на стадии мезолита заметно сильное разнообразие мужских и женских гаплогрупп, но на сегодняшний день C1a2, F, I, J — чрезвычайно мало представлены в генофонде европейцев. В этом своеобразном конкурентном соревновании гаплогруппа I2a1 вышла «победителем», но причины данной «победы» пока не объяснены научно.
Анализ образцов раннего неолита в Анатолии в работе Mathieson et al. (2015)[4] показал, что I2c-L596 существовала 8500 лет назад на территории современной Турции, но носители этой гаплогруппы в итоге были ассимилированы земледельцами периода неолита на пути из Плодородного полумесяца в Европу. В итоге в период неолита все гаплогруппы мезолита в Европе попали под массивную ассимиляцию многочисленными земледельцами неолита, пришедшими с Ближнего Востока. Но даже после этого I2a выделяется на общем фоне европейских гаплогрупп как очень распространённая (второе место — 16 % из 69 образцов периода неолита). Первое место — G2a — это линия, прямо происходящая от земледельцев, пришедших с территории Анатолии.
I2a распространились в старчево-кришской культуре (сущ. между 8000 — 6500 лет назад) и до сих пор являются основной мужской ДНК в Юго Восточной части Европы. В целом I2 активно продолжила развиваться на территории Европы в период медного и бронзового веков. Находки обнаруживаются в Испании — I2a2a-M223, I2a2, I2a2a1 (CTS616). На Юге России (Ямная культура I2a2a1 (CTS9183)), в Венгрии (культура Ватья I2a1), в Германии (Унетицкая культура I2a2b (L38) и I2c2), в Северной Италии (культура Ремеделлло I2a1a1)
Концентрация гаплогруппы снизилась, когда в Европу пришли индоевропейцы[5][6][7][8]. В настоящее время гапплогруппа I2 остаётся одной из наиболее распространённых в Европе, уступая Y-хромосомным гапплогруппам R1a и R1b.
На данный момент известно об определении гапплогруппы I2 у представителей большого количества различных археологических культур:
- Эпохи палеолита:
- гаплогруппа I2* была определён в останках из пещеры Франхти, Греция.
- субклад I2a найден у представителя азильской культуры из Бишонского грота на территории сегодняшней Швейцарии, датируемая древность этой находки — 13,5 тыс. лет[9][10].
- I2a2 (ISOGG, version 10.107) определён у позднего эпиграветтского образца San Teodoro 3 из сицилийской пещеры Сан-Теодоро, жившего после последнего ледникового максимума (LGM) 15 322—14 432 лет назад[11].
- Эпохи мезолита:
- Одна из субклад I2, I2a1a2 (ранее известная как I2a1b) определена в останках западноевропейского охотника-собирателя с мезолитической стоянки Хеффинген-Лошбур близ Хеффингена в Люксембурге (Человек из Лошбура)[12].
- I2a1a2 (ранее известный как I2a1b) определили у мезолитического охотника-собирателя, жившего около 7,5 тыс. лет назад на территории нынешней Швеции, возле нынешнего города Мутала[9][13].
- Гаплогруппа I2 обнаружена у представителей культуры Лепенски-Вир и у обитателя северо-западной Анатолии (Aktopraklık), живших ок. 8,4 тыс. лет назад[14].
- I2a2 (xI2a2a, I2a2b) или I2a1b*-Y10705>I2a1b2a-S2524>S2524*[15] (ISOGG 2018) определили у темнокожего и голубоглазого человека из Чеддара, I2a2 определили и у образца I6754 (Ogof_yr_ychen_1), датирующегося возрастом 7593—7204 лет до нашей эры[16].
- Эпохи неолита:
- В неолитической Британии гаплогруппа I2a2a1a1a2 определена у образца I6760 (BurnGround), датирующегося возрастом 3943—3711 лет до н. э. и у образца I6762 (UpperSwell). I2a2a1a1a определена у образца I3041 (Raschoille_1), I2a2a определена у образцов I3039 (Whitehawk_1), I3040 (Whitehawk_2), I2a2a1 определена у образца I6765 (CaveHa3_1), датирующегося возрастом 3654—3523 лет до нашей эры[16]
- I2a обнаружили у жившего примерно 4360—4490 лет назад представителя неолитической культуры Лендьель и у представителя культуры Кёрёш (Старчево-Кришская культура), жившего примерно 5650—5780 лет назад[17].
- Y-хромосомные гаплогруппы I2 и I2a были обнаружены у представителей энеолитической культуры Ремеделло[18].
- I2a1 обнаружен у представителя культуры ямочной керамики Ajvide 58 (2800—2000 BC)[19].
- У представителя мегалитической культуры из Испании (Ла Мина) была обнаружен субклад I2a2a1[13], у представителей мегалитической культуры из Франции (дольмен La Pierre Fritte) — I2a1[20].
- I2a2a1b2a-L801 определили у представителя культуры шаровидных амфор[21].
- I2c1a1 определён у представителей культуры Вартберг[22].
- Эпохи медного и бронзового веков:
- I2a2a определили у обитателя Атапуэрки (Испания), жившего в медном веке[23].
- I2a2a1b1b (L699, L703)>I2a2a1b1b1b-Y5669*[24] определили у представителя катакомбной культуры[25].
- I2 (I2a2a1b2b-L623/I2a2a-M223) определили у образца GLR002 (Necropoli di Gattolino, Италия), I2 (I2a1a1-M26) — у образца GLR004 (Necropoli di Gattolino)[26].
- I2a1a2b1a1-Y3120>I2a-CTS10228 определили у образца I18719 из пещеры Безданяча (Bezdanjača Cave) в Хорватии, который археологически датируется поздним бронзовым веком (3200 л. н., Croatia_MBA_LBA), но возможно является средневековым образцом[27][28].
- Средневековье:
- I2a1b1a-CTS9183, I2a1b1a1b1a1a1a-S391, I2a1b1a2b1-S390, I2a1b1a2b1a2a2-ZS20 определили у варваров VI века с кладбища лангобардов в Соладе (Венгрия)[29].
- I2a1a2b1a1a-Y3120/YP196>I2a1b2a1a1b-Y4460>Y3106>Y91535*[30] (Din) обнаружен у останков Глеба Святославича, внука Ярослава Мудрого (VK542, 950—850 л. н.)[31].
- I2a1b2a1a1a1a3-A16681>A16681*[32] определили у образца Sunghir 6 (730—850 л. н.) из средневекового захоронения на стоянке Сунгирь на окраине Владимира[33].
- I2a1b2a1a1a2b-Y16473>Y16473*[34] определили у образца KRA008 из Кракауэр-Берга (Krakauer Berg) в Восточной Германии, датирующегося возрастом примерно 599 л. н. (649—548 л. н.)[35].
Субклады
Примечание: на протяжении первого десятилетия XXI века классификация гаплогрупп неоднократно уточнялась и пересматривалась, поэтому в литературе и некоторых статьях википедии могут быть использованы устаревшие названия.
Приведённая ниже схема основана на исследовании ISOGG 2019[36], в котором обновлены данные прежней работы Т. М. Карафет и др. (Karafet et al. 2008).[37] Более ранние схемы опубликованы в ISOGG 2007[38] и ISOGG 2006[39].
I2 (L68, M438/P215/S31) (ранее I1b)
- I2a (CTS179)
- I2a1 (L460)
- I2a1a (P37.2)(ранее I2a, ещё ранее I1b1)
- I2a1a1 (CTS595)
- I2a1a1a (M26/L158) "Sardinian" (ранее I2a2, I1b1b)
- I2a1a1b (S21825) "Western"
- I2a1a2 (M423) (ранее I2a2, I2a1b)
- I2a1a2a (L161.1) "Isles"
- I2a1a2b (L621) "Dinaric"
- I2a1a1 (CTS595)
- I2a1b (M436) (ранее - I2a2)
- I2a1b1 (M223)
- I2a1b2 (FGC29562)
- I2a1a (P37.2)(ранее I2a, ещё ранее I1b1)
- I2a2 (L596)
- I2a1 (L460)
- I2b (L415)
I2a-M26/L158 "Sardinian"
Гаплогруппа I2a-M26/L158 "Sardinian" (старое название I2a1) составляет около 40 % всех мужских линий на Сардинии[40] (I2a1a1 ~ 39%[41]). Кроме того, она также обнаружена с низкой или умеренной частотой в Стране Басков и на Иберийском полуострове, а также составила 1,6 % (1/64) в выборке албанцев, живущих в Македонии[42] и 1,2 % (3/257) в выборке чехов.[43] Также данная гаплогруппа встречается на Британских островах.[44]
Предполагается, что ветвь M26 возникла около 18200 лет назад, а последний общий предок всех современных носителей данного варианта жил около 11700 лет назад.[44]
I2a-M423
I2a-M423 был типичен для населения Европы доиндоевропейского завоевания (см. раздел "Палеогенетика"). Согласно данным компании YFull субклад образовался за 18 200 лет до настоящего времени, расхождение субклады на ветви произошло около 13 400 лет назад. В настоящий момент состоит из следующих ветвей:
I2a-L161.1 "Isles"
Субклад I2a-L161.1 "Isles" встречается практически исключительно на Британских островах, в Ирландии, и по всей Великобритании, с очень низкой частотой - в континентальной Европе. Согласно данным компании YFull образовалась за 10 600 до настоящего времени, расхождение субклад произошло около 7900 лет назад.
I2a-L621
I2a1b2-L621/S392 возникла между 11 400 л. н. Последний общий предок современных носителей гаплогруппы I2a1b2-L621 жил 6 500 лет назад (даты определены по снипам компанией YFull)[45].
Субклад I2a-L621 встречается в основном в славянских странах. Максимальная частота наблюдается у динарских славян - словенцы, хорваты, боснийцы, сербы, черногорцы и македонцы. Так же высока частота в Болгарии, Румынии и Молдавии, на Западной Украине и в Белоруссии. В меньшей степени распространена в Албании, Греции, Словакии и в Польше. Незначительно представлена на юго-западе России. L621 в настоящее время встречается в пять - десять раз чаще, чем гаплогруппа G2a в Юго-Восточной Европе. Напротив, в период неолита G2a была в четыре раза более распространённой гаплогруппой. Подобная смена позиций объясняется индоевропейской миграцией и неразрывно связанными с ними славянскими миграциями населения. В частности, последним исторически зафиксированным рывком в этом распространении являются миграции славян между 6 - 9 веками нашей эры. Большая часть восточно-европейских славян это носители субклада L147.2 (он же CTS10228, CTS2180 или Y3111) "Dinaric" который возник около 5100 лет назад, незадолго до начала экспансии людей из ямной культуры. Но данный субклад имеет общего предка (TMRCA) около 3800 лет назад. И около 1900 лет назад I2a1b - L147.2 резко стал увеличивать свою численность, что совпадает по срокам с окончательным формированием славян. Когда готы, вандалы и бургундцы из Восточной Германии и в Польше вторглись в Римскую Империю, славяне смогли заполнить образовавшийся этнический вакуум и заселили эти территории. К 476 году, когда распалась Западная Римская империя, славяне расселились по Динарским Альпам и Балканам. К IX веку нашей эры славяне сформировали современные славяноязычные территории.
Субклад I2a-Disles (Dinaric Isles) - парафилический субклад, в который включают все субклады I2a-L621, кроме I2a-CTS10228 "Dinaric". Данный субклад с очень низкой частотой встречается на Британских островах и во Франции.
Субклад I2a1a2b1a1-CTS10228 (ранее I2a1a2b - Din, в ISOGG2017 - I2a1b2a1a) известен как «Динарская» гаплогруппа[46] (Dinaric). Также этот субклад определяется снипами CTS10228/V4023, CTS5966, CTS2180, CTS5996, FGC12075/YP194, FGC12076/YP217, FGC12077/YP193, FGC12078/Y3107, FGC12081/SK1237/V2129/Y3111, FGC12082/Y3774, FGC12084/Y3775, FGC12096/Y3121, L147.2, S12235, S15187/YP191, S17076/YP188, S17790/YP223, S19015 (ISOGG 2017). Субклад I2a1b2a1a-CTS10228 возник 5 100 л. н. Последний общий предок современных носителей гаплогруппы I2a1b2a1a-CTS10228 жил 3 400 лет назад (даты определены по снипам компанией YFull[27]).
I2a1b2a1a-CTS5966, CTS2180, CTS5996, CTS10228/V4023 распространён по всей юго-восточной Европе (Греция, Македония, Сербия, Хорватия, Словения, Босния и Герцеговина, Албания, Черногория, Чехия, Словакия, Польша, Болгария, Белоруссия, Россия, Украина, Венгрия, Австрия, Германия, Швейцария, Румыния, Италия, Молдавия, Литва, Латвия), а также найдена в Фенноскандии (Финляндия, Швеция, Норвегия) и в западной Анатолии. Среди «Динарской гаплогруппы» разделяют 2 кластера: разнообразный северный Динарский кластер (Dinaric North cluster - "Din N"); и южный Динарский кластер (Dinaric South cluster - "Din S") - крупнейшую Динарскую ветвь (около 40% всех представителей Din), которая характеризуется снипом I2a1b2a1a1a3-PH908. Наиболее высокая плотность CTS10228 — в Далмации (Хорватия) и Боснии-Герцеговине (>50 %)[47]. Расстояние до времени жизни общего предка субклады I2a-L147.2/CTS5966/CTS10228 — 2757±404 года[48].
I2a1b1
Распространение субгаплогруппы I2a1b1 хорошо коррелирует с распространением гаплогруппы I1, за исключением Фенноскандии, что позволяет предположить её наличие как минимум в одном из палеолитических убежищ, где также находилась гаплогруппа I1. Отсутствие корреляции между I1 и I2a1b1 в Фенноскандии может быть связано с тем, что на гаплогруппу I2a1b1 в наиболее ранних поселениях региона сильнее повлиял «эффект основателя» и генетический дрейф из-за её редкости, поскольку гаплогруппа I2a1b1 составляет менее 10 % от всего Y-хромосомного разнообразия населения на указанной территории за пределами Нижней Саксонии.
Распространение гаплогрупп I1 и I2a1b1 относительно хорошо коррелирует с историческими границами распространения носителей германских языков. Гаплогруппа I2a1b1 обнаружена более чем у 4 % населения лишь в Германии, Нидерландах, Бельгии, Дании, Англии (кроме Уэльса и Корнуолла), Шотландии, на южной оконечности Швеции и Норвегии, а также в провинциях Нормандия, Мэн, Анжу и Перш на северо-западе Франции, в Провансе на юго-востоке Франции, в исторических областях Италии — Тоскане, Умбрии и Лациуме; а также в Молдавии; в России — Рязанской области и в Мордовии. Представляется достаточно вероятным, что присутствие I1 и I2b1 в нынешних Франции, Англии и Италии, а также в восточной Европе связано уже с кельтской и германской экспансией, а в доиндоевропейское время эти гаплогруппы концентрировались лишь в северной Европе. Одна из субклад гаплогруппы I2a1b1, а именно I2a2a1a1-M284, обнаружен почти исключительно у населения Великобритании, что может говорить о его длительной истории существования на Британских островах. Интересно, что с небольшой частотой гаплогруппы I1 и I2b встречаются на территории исторических регионов Вифиния и Галатия в современной Турции, куда их могли принести кельты, мигрировавшие туда по приглашению Никомеда I Вифинского.
Гаплогруппа I2a1b1 также встречается примерно у 1 % населения Сардинии. Дивергенция субклада для мутации I2a2a-M223 произошла около 14,6±3,8 тысяч лет назад (Rootsi 2004). Возраст вариации YSTR для субклада I2a2a-M223 составляет 13,2±2,7 тысяч лет (Rootsi 2004) или 12,3±3,1 тысяч лет (Underhill 2007).
I2a1b2a
Гаплогруппа I2a1b2a - L38/S154 была обнаружена в скелетных останках, найденных в пещере Лихтенштейн — археологическом памятнике бронзового века в центральной Германии, где также были найдены артефакты культуры полей погребальных урн[49]. Из 19 мужских останков пещеры у 13 была обнаружена гаплогруппа I2a1b2a , у одного — R1b, и у двоих — R1a. Предположительно, пещера находилась в центре тогдашнего распространения гаплогруппы I2a1b2.
Примечания
- YTree v5.02 at 11 February 2017
- Pineau, JC; Delamarche, P; Bozinovic, S (2012-05-24). «Average height of adolescents in the Dinaric Alps. They are also reputed to have the tallest males in Europe. Study claims it is not complete as yet». Comptes Rendus Biologies. 328 (9): 841-46.
- Grasgruber, Pavel; Popović, Stevo; Bokuvka, Dominik; Davidović, Ivan; Hřebíčková, Sylva; Ingrová, Pavlína; Potpara, Predrag; Prce, Stipan; Stračárová, Nikola (2017-04-01). «The mountains of giants: an anthropometric survey of male youths in Bosnia and Herzegovina». Royal Society Open Science. http://rsos.royalsocietypublishing.org/content/4/4/161054
- Eppie R. Jones et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians (Nature Communications volume 6, Article number: 8912 (2015)).
- Mesolithic Western Eurasian DNA
- Eppie R. Jones et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians // Nature Communications, 16 November 2015
- There’s no such thing as a 'pure' European—or anyone else // Science | AAAS, 2017
- Ancient DNA reveals 'genetic continuity’ between Stone Age and modern populations in East Asia // University of Cambridge
- The genetic history of Ice Age Europe
- Jones E. R. et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians, 2015
- Gabriele Scorrano et al. Genomic ancestry, diet and microbiomes of Upper Palaeolithic hunter-gatherers from San Teodoro cave (Sicily, Italy), December 08, 2021
- «Ancient human genomes suggest three ancestral populations for present-day Europeans»
- Haak, W. et al. (2015), Massive migration from the steppe is a source for Indo-European languages in Europe
- Zuzana Hofmanova. Palaeogenomic and biostatistical analysis of ancient DNA data from Mesolithic and Neolithic skeletal remains, 2017
- I-S2524 YTree
- Selina Brace et al. Ancient genomes indicate population replacement in Early Neolithic Britain (Supplementary Data 1), 2019
- Genome flux and stasis in a five millennium transect of European prehistory
- Morten E. Allentoft et al. «Population genomics of Bronze Age Eurasia», 10 June 2015
- Skoglund P. et al. Genomic diversity and admixture differs for Stone-Age Scandinavian foragers and farmers // Science, 2014
- Lacan, M. et al. (2011), Ancient DNA suggests the leading role played by men in the Neolithic dissemination
- Hannes Schroeder et al. Unraveling ancestry, kinship, and violence in a Late Neolithic mass grave, March 25, 2019
- Alexander Immel et al. Genome-wide study of a Neolithic Wartberg grave community reveals distinct HLA variation and hunter-gatherer ancestry, 25 January 2021
- Ancient genomes link early farmers from Atapuerca in Spain to modern-day Basques
- I-Y5669 YTree
- Morten E. Allentoft et al. «Population genomics of Bronze Age Eurasia» // Nature, 2015
- Tina Saupe et al. Ancient genomes reveal structural shifts after the arrival of Steppe-related ancestry in the Italian Peninsula, May 10, 2021 (Table 1. Archaeological information, genome coverage, genetic sex, mtDNA, and Y chromosome haplogroups of the individuals of this study)
- I-CTS10228 YTree
- Nick Patterson et al. Large-scale migration into Britain during the Middle to Late Bronze Age // Nature, 22 December 2021
- Carlos Eduardo G. Amorim, Krishna R. Veeramah et al. Understanding 6th-century barbarian social organization and migration through paleogenomics, 2018
- YTree, id:VK542
- Ashot Margaryan et al. Population genomics of the Viking world, 2019 (bioRxiv, ResearchGate)
- YTree, I-A16681
- Sikora M. et al. Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers, Science 10.1126/science.aao1807 (2017).
- I-Y16473 YTree
- Cody Parker et al. A systematic investigation of human DNA preservation in medieval skeletons, 26 October 2020 (bioRxiv, May 22, 2020)
- 2019 Y-DNA Haplogroup I
- Tatiana M. Karafet et al., New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree, Genome Research, doi:10.1101/gr.7172008 (2008)
- ISOGG 2007
- ISOGG 2006
- アーカイブされたコピー. Дата обращения: 19 марта 2008. Архивировано 6 марта 2009 года.
- Charleston W. K. Chiang et al. Genomic history of the Sardinian population
- Vincenza Battaglia et al., "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe, " European Journal of Human Genetics advance online publication 24 December 2008; doi: 10.1038/ejhg.2008.249.
- F. Luca, F. Di Giacomo, T. Benincasa et al., "Y-Chromosomal Variation in the Czech Republic, " American Journal of Physical Anthropology 132:132-139 (2007).
- I-L158 YTree
- I-L621 YTree
- ТЕЗИСНЫЕ ВЫВОДЫ СТАТЬИ О ЭВОЛЮЦИИ «ДИНАРСКОЙ ПОДГРУППЫ» ГАПЛОГРУППЫ I2A, 2014
- Pericic et al., High-Resolution Phylogenetic Analysis of Southeastern Europe Traces Major Episodes of Paternal Gene Flow Among Slavic Populations
- Веренич В. Актуальные проблемы изучения так называемого динарского кластера гаплогруппы I2a
- Lichtenstein Cave Data Analysis Архивная копия от 6 марта 2009 на Wayback Machine | Ken Nordtvedt: «The Lichtenstein cave ydna haplotypes show three from the new S23+(xM223) I2b* (ISOGG 2008) tree»
Ссылки
- ISOGG 2016 Y-DNA Haplogroup I
- Гаплогруппа I2
- Phylogeography of Y-Chromosome Haplogroup I (Rootsi 2004)
- Frequency and Variance of I2 (called I1b)
- Map of I2a1 (called I1b)
- Map of I2a2 (called I1b2)
- Map of I2b (called I1c, also classed as I1b2a)
- [www.familytreedna.com/public/I2aHapGroup/ I2a project at FTDNA]
- [www.familytreedna.com/public/M223-Y-Clan/ I2b1 — M223 project at FTDNA]
- [www.familytreedna.com/public/I2b2/default.aspx I2b2 L38+ Project]
- Danish Demes Regional DNA Project: Y-DNA Haplogroup I2 (called I1b) (недоступная ссылка)
- British Isles DNA Project
{kind=link}
{kind=link}
{kind=link}
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 GHIJK | |||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | LT (K1) | K2 | ||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/NO/NO1 | K2b | ||||||||||||||||||||||||||||||
| N | O | K2b1 | P (K2b2)/P1 | ||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||