CREBBP
CREB-свя́зывающий бело́к (англ. (cyclic AMP response element binding protein) bind, CREBBP, CBP) — белок, который у человека кодируется геном CREBBP, локализованным на 16-й хромосоме[1][2]. CBP (как и его близкий гомолог, p300) функционирует как транскрипционный коактиватор, то есть посредством белок-белковых взаимодействий связывает различные белковые трансактиваторы транскрипции с основным транскрипционным комплексом[3]. Впервые описан исследователями в 1993 году[1].
Ген
Ген CREBBP у человека располагается на 16-ой хромосоме и занимает локус 16p13.3. Ген содержит 34 экзона и экспрессируется повсеместно[4], в том числе и на эмбриональном этапе развития. Ген расположен на минус-цепи[5]. Длина гена составляет около 190 килобаз, транскрипция идёт от центромеры до теломеры. Длина мРНК составляет 8,7 кб, из которых на кодирующую последовательность приходится 7,3 кб[6].
Гомологи CREBBP имеются у многих многоклеточных организмов, в том числе мух, червей и растений, однако отсутствуют у низших эукариот, например, дрожжей[7]. У данио-рерио паттерны экспрессии CREBBP и его метилирования играют роль в ходе развития организма[8]. У ящерицы североамериканского красногорлого анолиса экспрессия CBP различается у двух полов[9].
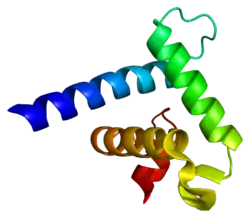
Структура
CREBBP представляет собой белок с молекулярной массой 265 кДа[10], состоящий из 2442 аминокислот. Локализуется в ядре[6]. По строению он очень схож с гомологичным[6] белком p300, вместе с которым CBP составляет семейство коактиваторов p300-CBP[7].
Большая часть функциональных доменов этих двух белков высоко консервативна. В числе этих доменов 4 известных трансактивационных домена (TAD):
- первый цистеин/гистидин-обогащённый участок (CH1), в который входит транскрипционный адаптерный цинковый палец 1 (англ. transcriptional adapter zinc finger 1, TAZ1);
- домен KIX, взаимодействующий с CREB;
- ещё один цистеин/гистидин-обогащённый участок (СН3), содержащий цинковый палец TAZ2 и цинковый палец типа ZZ;
- коактиваторный связывающий домен ядерных рецепторов (англ. nuclear receptor co-activator binding domain, NCBD), также известный как интерферон-связывающий домен (англ. interferon-binding domain, IBiD). Эти TAD обеспечивают различные белок-белковые взаимодействия CBP с ДНК-связывающими факторами транскрипции, ключевыми компонентами транскрипционной машинерии и прочими коактиваторами транскрипции (подробнее см. ниже).
Кроме того, и p300, и CBP также содержат домен с гистонацетилтрансферазной активностью (НАТ), который ацетилирует гистоны и другие белки. Рядом с ним находится бромодомен, который связывается с ацетилированными остатками лизина и может привлекать СВР к специфическим сайтам на хромосомах. Рядом с каталитическим участком располагается ещё один цистеин/гистидин-обогащённый участок (CH2), включающий бромодомен и PHD-палец, который связывается с коферментом А. Наличие этого мотива уникально для CBP, он отсутствует у других белков с гистонацетилтрансферазной активностью[7][11][12].

Действие на клеточном уровне
Ген CREBBP экспрессируется повсеместно и участвует в транскрипционной коактивации многих факторов транскрипции. Белок CREBBP впервые был описан как ядерный белок, который связывается белком CREB. Этот ген, как теперь известно, играет важную роль в эмбриональном развитии, контроле роста и поддержании гомеостаза[13]. Было показано, что мыши, у которых нокаутирован ген CREBBP или EP300, кодирующий белок р300, погибают на ранних этапах развития[14][15]. Кроме того, мыши, у которых имелся только один функциональный аллель каждого из генов CREBBP и EP300 вместо двух, также погибали в ходе эмбриогенеза. По-видимому, для развития эмбриона критическое значение имеет общее суммарное содержание белков CBP и p300, которое в обоих случаях составляло половину от нормального[14]. В отличие от организма в целом, отдельные клетки могут развиваться в отсутствии белков CBP или р300. Например, у мышей В- и Т-клетки, лишённые или СВР, или р300, развивались нормально в условиях in vivo, а лишённые обоих белков одновременно погибали[16][17].
CBP, как и p300, повышает экспрессию генов-мишеней при помощи следующих основных механизмов:
- релаксация хроматина в области промоторов при помощи ацетилирования гистонов;
- рекрутирование ключевых элементов транскрипционного комплекса, в том числе РНК-полимеразы II, к промоторам;
- функционирование в качестве опорного белка (белок скэффолда) для стабилизации взаимодействий других факторов транскрипции с транскрипционным комплексом[18].
CBP, как и p300, предпочитает ацетилировать N-концевые хвосты гистонов, а именно остатки К12 (лизин 12) и К15 гистона H2B, K14 и K18 гистона Н3 и К5 и К8 гистона Н4. Однако СВР ацетилирует не только гистоны, но и негистоновые белки, такие как различные факторы и коактиваторы транскрипции. Эти модификации могут изменять белок-белковые взаимодействия, взаимодействия белков с ДНК, а также ядерную локализацию белков. Другие важные партнёры СВР и р300 — белки репликации и репарации ДНК, в частности, PCNA, Flap-эндонуклеаза 1, ДНК-полимераза β, тимин-ДНК-гликозилаза. Кроме того, СВР и р300 могут взаимодействовать и с белками, не имеющими непосредственного отношения к хроматину, в частности, с циклином Е и циклин-зависимой киназой 2, тем самым участвуя в регуляции клеточного цикла. СВР и р300 участвуют в регуляции разрушения транскрипционного фактора p53. Было показано, что регион СН-1 СВР и р300 может проявлять полиубиктивинлигазную активность р53, тем самым непосредственно влияя на его деградацию. Наконец, СВР, как и р300, могут ацетилировать белки, связанные с ядерным транспортом, в частности, импортин-α1 и импортин-α7[7].
CBP используется в качестве белка скэффолда многими транскрипционными факторами, в числе которых c-jun, c-myb, MyoD, E2F1, YY1, а также члены суперсемейства рецепторов стероидных гормонов (полный список белков, с которыми взаимодействует CBP, см. ниже)[19].
Результаты последних (по состоянию на 2009 год) исследований показали, что СВР-опосредованное посттрансляционное N-гликозилирование изменяет конформацию белков, взаимодействующих с CBP, регулируя таким образом экспрессию генов, клеточный рост и дифференцировку[20].
Физиологические функции
CBP задействован в сигнальных путях G-белков и через них принимает участие в ответе клетки на связывание адреналина. Некоторые G-белки в активированном состоянии стимулируют аденилатциклазу, в результате чего уровень сАМР в клетке повышается. сАМР активирует протеинкиназу А (РКА), которая состоит из четырёх субъединиц: двух регуляторных и двух каталитических. Связывание сАМР с регуляторными субъединицами вызывает освобождение каталитических субъединиц, которые вследствие этого могут переместиться в ядро и взаимодействовать с транскрипционными факторами, таким образом влияя на экспрессию генов. Транскрипционный фактор CREB, связывающийся с последовательностью ДНК, называемой элементом отклика на сАМР (CRE), фосфорилируется РКА по остатку серина (Ser 133) в домене KID. Эта модификация стимулирует взаимодействие домена KID CREB с доменом KIX CBP или р300, в результате чего усиливается транскрипция генов, подконтрольных CREB, в том числе и тех, которые участвуют в глюконеогенезе. Этот сигнальный путь запускается при связывании адреналина с клеткой-мишенью[21].
Огромное количество экспериментальных данных свидетельствуют о ключевой роли ацетилирования гистонов в пластичности синапсов у млекопитающих (в том числе долговременной потенциации), памяти и поведенческой адаптации к окружающей среде. Первоначальные свидетельства связаны с наблюдением, касающимся больных синдромом Рубинштейна — Тейби: у них мутации, затрагивающие CBP, приводят к умственной отсталости. Кроме того, мыши, мутантные по CREBBP, имели нарушения долговременной памяти[22]. Дальнейшие эксперименты показали, что CBP важен не только для нейронной пластичности, но также выживания нейронов, связанного с их активностью[23], и нейрогенеза[24].
Нарушения со стороны иммунной системы, проявляющиеся у пациентов с синдромом Рубинштейна — Тейби, свидетельствуют о важной роли CBP в функционировании иммунной системы и воспалительных процессах. По-видимому, этот эффект обусловлен ненормально пониженной экспрессией таких важных для иммунитета белков, как CREB, NF-κB, c-jun, c-Fos, BCL2 и c-Myc, обусловленной отсутствием активности СВР[25]. В самом деле, регулируя активность транскрипционных факторов NF-κB и AP-1, которые активируют гены, связанные с хроническим и острым воспалением, СВР может играть важную роль в контроле воспаления на уровне транскрипции[26]. Установлено, что СВР задействован в экспрессии белка CD59, который участвует в защите клеток организма от атак со стороны системы комплемента[27].
СВР играет важную роль в функционировании женской репродуктивной системы. На мышах было показано, что он необходим для экспрессии лютеинизирующего гормона-бета и поддержании нормальной фертильности[28]. Кроме того, СВР необходим для экспрессии генов-мишеней лютеинизирующего гормона во время овуляции[29]. Он также участвует в клеточном ответе на связывание эстрогена[30].
CBP и p300 играют важную роль в функционировании фоторецепторов (палочек и колбочек): колбочко-палочковый гомеобоксовый белок (англ. cone-rod homeobox protein, CRX) привлекает их к промоторам необходимых генов, где они ацетилируют гистоны и способствуют их экспрессии. Интересно, что нокаут одного из генов CREBBP и EP300 не оказывал почти никакого эффекта на фоторецепторные клетки, а нокаут обоих генов сильно изменял их морфологию и функционирование. Такой эффект был связан со снижением ацетилирования гистонов Н3 и Н4, что является следствием нефункциональности CBP и p300[31].
Показана возможность участия СВР в адипогенезе[32].
Регуляция
Ацетилтрансферазная активность CBP положительно регулируется фосфорилированием МАР-киназой p42/p44, Cdk2 и протеинкиназой А. При этом фосфорилирование консервативного остатка серина в положении 89 (S89) протеинкиназой С-δ снижает ацетилтрансферазную активность СВР. Ацетилтрансферазная активность СВР может изменяться (как в положительную, так и в отрицательную сторону) при взаимодействии с другими белками. Например, она усиливается при взаимодействии с такими транскрипционными факторами, как C/EBP-α, NFE2 и HNF-1α и понижается при взаимодействии с транскрипционным фактором PU.1. При этом все перечисленные белки взаимодействуют с одним и тем же сайтом СВР — участком СН3. Механизмы того, как белки, взаимодействующие с СВР в одном и том же сайте, влияют на его активность противоположным образом, в настоящий момент не ясны[7].
В настоящее время разрабатываются искусственные ингибиторы CBP, некоторые из которых могут найти потенциальное применение в медицине (например, ICG-001 подавляет рост раковых клеток в поджелудочной железе[33]). Ингибиторы могут связываться, например, с бромодоменом CBP[34][35].
Клиническое значение
Синдром Рубинштейна — Тейби
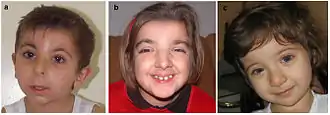
Мутации в гене CREBBP вызывают синдром Рубинштейна — Тейби (RTS)[36]. Однако RTS может развиваться и при мутациях гена EP300. Пациенты с RTS характеризуются множественными врождёнными пороками, отставании в умственном развитии и постнатальном развитии, микроцефалией, характерными лицевыми аномалиями, широкими, часто заострёнными пальцами рук и увеличенными пальцами ног. Частота встречаемости этого заболевания составляет 1 случай на 100 тысяч—125 тысяч новорождённых[37]. Как правило, пациенты с RTS имеют повышенный риск развития опухолей. У человека фенотипы, проявляющиеся при гаплонедостаточности по CREBBP и при мелких делециях и мутациях, приводящих к образованию укороченной формы белка, не различаются. У мышей гетерозиготная делеция или укорочение CBP приводит к фенотипу, напоминающему RTS. Список описанных мутаций CREBBP, приводящих к развитию RTS, постоянно пополняется[38][39]. В общем случае к RTS приводят те мутации CREBBP, которые подавляют его гистонацетилтрансферазную активность[7].
Острый миелоидный лейкоз
Хромосомные транслокации, затрагивающие CREBBP, связывают с острым миелоидным лейкозом (AML)[13][40]. К AML также могут приводить транслокации в EP300. Более того, транслокации, затрагивающие CREBBP и вызывающие AML, могут быть следствием противораковой химиотерапии (например, при лечении рака молочной железы)[41]. В случае этих сбалансированных транслокаций 5'-конец генов MLL, MOZ или MORF сшивается с 3'-концом гена CBP или p300, и наоборот. Это приводит к образованию химерных белков, например, MLL–CBP и CBP–MLL. Химерные белки, у которых CBP (p300) сливается с другим своим С-концом (MLL–CBP, MLL–p300, MOZ–CBP, MOZ–p300, MORF–CBP), по-видимому, играют ключевую роль в лейкемогенезе, а мРНК, кодирующие обратные белки (например, CBP–MLL), у пациентов с AML не обнаруживаются. В отличие от RTS, при котором CBP и p300 нефункциональны, в случае описанных выше транслокаций HAT-домены CBP и p300 остаются интактными в химерных белках. Однако CBP и p300 начинают работать неправильно (например, они работают не с теми последовательностями), поэтому в случае AML имеют место мутации приобретения, а не утраты функции[7].
Другие злокачественные заболевания
В отличие от p300, который является классическим супрессором опухолей, ситуация с CBP менее ясна. С одной стороны, предрасположенность к опухолеобразованию у пациентов с RTS, а также тот факт, что и CBP, и p300 являются мишенями вирусов, изменяющих ДНК (аденовирус, SV40, папилломавирус человека), говорит о том, что CBP тоже может функционировать как опухолевый супрессор. Однако в тех опухолевых клетках, в которых имелись мутации в CREBBP, мутированной оказывалась только одна аллель, другая же оставалась интактной и функциональной. Это противоречит представлению о CBP как о классическом опухолевом супрессоре[7]. Роль СВР была показана в таких видах карциномы, как рак толстой кишки[42], лёгких[43][44], поджелудочной железы[33] и других раковых заболеваниях.
Установлено, что CBP-зависимый сигнальный путь Wnt/β-катенина находится в постоянно активированном состоянии у раковых клеток, устойчивых к действию противоракового препарата доксорубицина. Нарушение взаимодействия CBP с β-катенином, останавливающее работу сигнального пути, фармакологическим путём может восстановить чувствительность раковых клеток к этому препарату и улучшить прогноз для пациента[45]. Кроме того, показано, что ингибитор комплекса CREB-CBP, нафтол-AS-TR-фосфата, может применяться для лечения рака лёгкого[46].
Нарушения со стороны нервной системы
Многие нейродегенеративные заболевания человека (хорея Хантингтона (HD), синдром Кеннеди и другие) связаны с удлинением области полиглутаминовых повторов в определённых белках (в частности, Htt в случае HD и андрогенового рецептора при синдроме Кеннеди), которое приводит к образованию нерастворимых агрегатов. Они могут связываться с полиглутаминовыми трактами нормальных белков, в частности, CBP, который имеет участок из 18 остатков глутамина (остатки 2199—2216) в своём Q-обогащённом С-конце. При связывании с такими агрегатами происходит инактивация СВР, которая, по-видимому, и обусловливает их токсичность: эктопическая сверхэкспрессия СВР уменьшала смертность клеток в культуре, вызванную образованием глутаминовых агрегатов. В ходе экспериментов с дрозофилами было показано, что развитие нейродегенеративного фенотипа подавлялось при обработке ингибиторами деацетилаз, что служит доказательством необходимости гистонацетилтрансферазной функции CBP для предотвращения развития нейродегенеративных заболеваний[7].
Было показано участие CBP (и р300) в апоптозе нейронов, который происходит при болезни Альцгеймера (AD). Предполагается, что при этом СВР разрушается каспазой 6, что снижает уровень ацетилирования гистонов. Интересно, что увеличение содержания СВР связано с ранним развитием AD. Было продемонстрировано, что опосредованное пресенилином-1 разрезание N-кадгерина приводило к образованию пептида N-Cad/CTF2, который в цитоплазме связывается с СВР и способствует его протеасомному разрушению. Уменьшение количества СВР приводит к подавлению транскрипции генов, активируемых CREB. Стоит отметить, что мутации пресенилина-1 наблюдаются при семейных формах AD, причём они могут приводить как утрате, так и приобретению функции CBP[7]. Кроме того, показано, что CBP функционирует как регулятор циркадных ритмов, и его разрушение, индуцированное бета-амилоидом, приводит к нарушению циркадных ритмов при AD[47]. Имеются данные, свидетельствующие о связи СВР с развитием бокового амиотрофического склероза. Предполагается, что препараты, компенсирующие утрату функции СВР, могут использоваться в лечении нейродегенеративных заболеваний[48].
Имеются данные, что нарушение гистонацетилтрансферазной активности CBP вызывает проблемы при формировании долговременной памяти[49].
CBP может быть вовлечён в развитие некоторых психиатрических нарушений. Исследование 2012 года показало, что однонуклеотидные полиморфизмы в гене CREBBP могут быть ассоциированы с развитием шизофрении[50], большого депрессивного расстройства и биполярного аффективного расстройства и влиять на ответ пациентов на лечение[51]. Более того, полиморфизмы в этом гене могут быть связаны с чувствительностью к наркотическим веществам, например, героину, а также алкоголю, и служить факторами развития привыкания к ним[52]. В прилежащем ядре СВР регулирует индуцированное кокаином ацетилирование гистонов и необходим для развития поведенческих реакций, связанных с употреблением кокаина[53].
Другие заболевания
CBP и p300 часто являются мишенями вирусов[19]. Например, показано, что коровый белок вируса гепатита B усиливает транскрипцию генов, подконтрольных CRE, действуя на путь CRE/CREB/CBP[54].
В 2012 году было показано участие CBP в развитии муковисцидоза[55].
Взаимодействия с другими белками
В таблице ниже перечислены ключевые белки, взаимодействующие с различными доменами CREBBP[12].
| Домен | Белки, которые с ним взаимодействуют | Группа |
|---|---|---|
| TAZ1 | TBP p53 FOXO3a HIF1A CITED2 STAT2[12] | Общий транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор |
| KIX (сайт c-Myb) | CREB c-Myb p53 FOXO3a[12] | Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор |
| KIX (сайт MLL) | MLL p53 FOXO3a HTLV-1HBZ C-jun E2A HTLV-1 Tax | Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Вирусный белок |
| KIX (два сайта) | p53 FOXO3a | Транскрипционный фактор Транскрипционный фактор |
| KIX (сайт связывания неизвестен) | BRCA1 SREBP | Транскрипционный фактор Транскрипционный фактор |
| Бромодомен | Гистон H4 p53 | Гистон Транскрипционный фактор |
| TAZ2 | E1A TFIIB PCAF GCN5 p53 FOXO3a STAT1 MEF2 | Белок аденовируса Общий транскрипционный фактор Коактиватор Коактиватор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор Транскрипционный фактор |
| NCBD (IBid) | p53 IRF3 ACTR[12] | Транскрипционный фактор Транскрипционный фактор Коактиватор |
| Мотивы LXXLL | Ретиноидный X-рецептор Эстрогеновый рецептор Андрогеновый рецептор[12] | Ядерный рецептор Ядерный рецептор Ядерный рецептор |
Примечания
- Chrivia J. C., Kwok R. P., Lamb N., Hagiwara M., Montminy M. R., Goodman R. H. Phosphorylated CREB binds specifically to the nuclear protein CBP. (англ.) // Nature. — 1993. — Vol. 365, no. 6449. — P. 855—859. — doi:10.1038/365855a0. — PMID 8413673.
- Wydner K. L., Bhattacharya S., Eckner R., Lawrence J. B., Livingston D. M. Localization of human CREB-binding protein gene (CREBBP) to 16p13.2-p13.3 by fluorescence in situ hybridization. (англ.) // Genomics. — 1995. — Vol. 30, no. 2. — P. 395—396. — PMID 8586450.
- Mink S., Haenig B., Klempnauer K. H. Interaction and functional collaboration of p300 and C/EBPbeta. (англ.) // Molecular and cellular biology. — 1997. — Vol. 17, no. 11. — P. 6609—6617. — PMID 9343424.
- NCBI. CREBBP CREB binding protein [ Homo sapiens (human) ].
- GenAge entry for CREBBP (Homo sapiens).
- Jean-Loup Huret. CBP (CREB-binding protein) // Atlas of Genetics and Cytogenetics in Oncology and Haematology. — 2000. — doi:10.4267/2042/37634.
- Kalkhoven E. CBP and p300: HATs for different occasions. (англ.) // Biochemical pharmacology. — 2004. — Vol. 68, no. 6. — P. 1145—1155. — doi:10.1016/j.bcp.2004.03.045. — PMID 15313412.
- Batut J., Duboé C., Vandel L. Expression patterns of CREB binding protein (CREBBP) and its methylated species during zebrafish development. (англ.) // The International journal of developmental biology. — 2015. — Vol. 59, no. 4-6. — P. 229—234. — doi:10.1387/ijdb.140197LV. — PMID 26260685.
- Kerver H. N., Wade J. Sexually dimorphic expression of CREB binding protein in the green anole brain. (англ.) // General and comparative endocrinology. — 2015. — doi:10.1016/j.ygcen.2015.09.007. — PMID 26363452.
- Chen X. N., Korenberg J. R. Localization of human CREBBP (CREB binding protein) to 16p13.3 by fluorescence in situ hybridization. (англ.) // Cytogenetics and cell genetics. — 1995. — Vol. 71, no. 1. — P. 56—57. — PMID 7606928.
- Spiegelman B. M., Heinrich R. Biological control through regulated transcriptional coactivators. (англ.) // Cell. — 2004. — Vol. 119, no. 2. — P. 157—167. — doi:10.1016/j.cell.2004.09.037. — PMID 15479634.
- Wang F., Marshall C. B., Ikura M. Transcriptional/epigenetic regulator CBP/p300 in tumorigenesis: structural and functional versatility in target recognition. (англ.) // Cellular and molecular life sciences : CMLS. — 2013. — Vol. 70, no. 21. — P. 3989—4008. — doi:10.1007/s00018-012-1254-4. — PMID 23307074.
- Entrez Gene: CREBBP (CREB-binding protein).
- Yao T. P., Oh S. P., Fuchs M., Zhou N. D., Ch'ng L. E., Newsome D., Bronson R. T., Li E., Livingston D. M., Eckner R. Gene dosage-dependent embryonic development and proliferation defects in mice lacking the transcriptional integrator p300. (англ.) // Cell. — 1998. — Vol. 93, no. 3. — P. 361—372. — PMID 9590171.
- Tanaka Y., Naruse I., Hongo T., Xu M., Nakahata T., Maekawa T., Ishii S. Extensive brain hemorrhage and embryonic lethality in a mouse null mutant of CREB-binding protein. (англ.) // Mechanisms of development. — 2000. — Vol. 95, no. 1-2. — P. 133—145. — PMID 10906457.
- Kasper L. H., Fukuyama T., Biesen M. A., Boussouar F., Tong C., de Pauw A., Murray P. J., van Deursen J. M., Brindle P. K. Conditional knockout mice reveal distinct functions for the global transcriptional coactivators CBP and p300 in T-cell development. (англ.) // Molecular and cellular biology. — 2006. — Vol. 26, no. 3. — P. 789—809. — doi:10.1128/MCB.26.3.789-809.2006. — PMID 16428436.
- Xu W., Fukuyama T., Ney P. A., Wang D., Rehg J., Boyd K., van Deursen J. M., Brindle P. K. Global transcriptional coactivators CREB-binding protein and p300 are highly essential collectively but not individually in peripheral B cells. (англ.) // Blood. — 2006. — Vol. 107, no. 11. — P. 4407—4416. — doi:10.1182/blood-2005-08-3263. — PMID 16424387.
- Goodman R. H., Smolik S. CBP/p300 in cell growth, transformation, and development. (англ.) // Genes & development. — 2000. — Vol. 14, no. 13. — P. 1553—1577. — PMID 10887150.
- Goldman P. S., Tran V. K., Goodman R. H. The multifunctional role of the co-activator CBP in transcriptional regulation. (англ.) // Recent progress in hormone research. — 1997. — Vol. 52. — P. 103—119. — PMID 9238849.
- Siddique H., Rao V. N., Reddy E. S. CBP-mediated post-translational N-glycosylation of BRCA2. (англ.) // International journal of oncology. — 2009. — Vol. 35, no. 2. — P. 387—391. — PMID 19578754.
- Mayr B., Montminy M. Transcriptional regulation by the phosphorylation-dependent factor CREB. (англ.) // Nature reviews. Molecular cell biology. — 2001. — Vol. 2, no. 8. — P. 599—609. — doi:10.1038/35085068. — PMID 11483993.
- Miller C. A. Forgot your HAT? CBP might be to blame. (англ.) // Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. — 2011. — Vol. 36, no. 8. — P. 1543—1544. — doi:10.1038/npp.2011.79. — PMID 21673711.
- Bito H., Takemoto-Kimura S. Ca(2+)/CREB/CBP-dependent gene regulation: a shared mechanism critical in long-term synaptic plasticity and neuronal survival. (англ.) // Cell calcium. — 2003. — Vol. 34, no. 4-5. — P. 425—430. — PMID 12909086.
- Lopez-Atalaya J. P., Ciccarelli A., Viosca J., Valor L. M., Jimenez-Minchan M., Canals S., Giustetto M., Barco A. CBP is required for environmental enrichment-induced neurogenesis and cognitive enhancement. (англ.) // The EMBO journal. — 2011. — Vol. 30, no. 20. — P. 4287—4298. — doi:10.1038/emboj.2011.299. — PMID 21847097.
- Torres L. C., Kulikowski L. D., Ramos P. L., Sugayama S. M., Moreira-Filho C. A., Carneiro-Sampaio M. Disruption of the CREBBP gene and decreased expression of CREB, NFκB p65, c-JUN, c-FOS, BCL2 and c-MYC suggest immune dysregulation. (англ.) // Human immunology. — 2013. — Vol. 74, no. 8. — P. 911—915. — doi:10.1016/j.humimm.2013.04.024. — PMID 23643710.
- Matt T. Transcriptional control of the inflammatory response: a role for the CREB-binding protein (CBP). (англ.) // Acta medica Austriaca. — 2002. — Vol. 29, no. 3. — P. 77—79. — PMID 12168567.
- Du Y., Teng X., Wang N., Zhang X., Chen J., Ding P., Qiao Q., Wang Q., Zhang L., Yang C., Yang Z., Chu Y., Du X., Zhou X., Hu W. NF-κB and enhancer-binding CREB protein scaffolded by CREB-binding protein (CBP)/p300 proteins regulate CD59 protein expression to protect cells from complement attack. (англ.) // The Journal of biological chemistry. — 2014. — Vol. 289, no. 5. — P. 2711—2724. — doi:10.1074/jbc.M113.525501. — PMID 24338025.
- Miller R. S., Wolfe A., He L., Radovick S., Wondisford F. E. CREB binding protein (CBP) activation is required for luteinizing hormone beta expression and normal fertility in mice. (англ.) // Molecular and cellular biology. — 2012. — Vol. 32, no. 13. — P. 2349—2358. — doi:10.1128/MCB.00394-12. — PMID 22508984.
- Zhang Y. L., Xia Y., Yu C., Richards J. S., Liu J., Fan H. Y. CBP-CITED4 is required for luteinizing hormone-triggered target gene expression during ovulation. (англ.) // Molecular human reproduction. — 2014. — Vol. 20, no. 9. — P. 850—860. — doi:10.1093/molehr/gau040. — PMID 24878634.
- Pradhan M., Baumgarten S. C., Bembinster L. A., Frasor J. CBP mediates NF-κB-dependent histone acetylation and estrogen receptor recruitment to an estrogen response element in the BIRC3 promoter. (англ.) // Molecular and cellular biology. — 2012. — Vol. 32, no. 2. — P. 569—575. — doi:10.1128/MCB.05869-11. — PMID 22083956.
- Hennig A. K., Peng G. H., Chen S. Transcription coactivators p300 and CBP are necessary for photoreceptor-specific chromatin organization and gene expression. (англ.) // Public Library of Science ONE. — 2013. — Vol. 8, no. 7. — P. e69721. — doi:10.1371/journal.pone.0069721. — PMID 23922782.
- Li Q., Peng H., Fan H., Zou X., Liu Q., Zhang Y., Xu H., Chu Y., Wang C., Ayyanathan K., Rauscher F. J., Zhang K., Hou Z. The LIM protein Ajuba promotes adipogenesis by enhancing PPARγ and p300/CBP interaction. (англ.) // Cell death and differentiation. — 2015. — doi:10.1038/cdd.2015.83. — PMID 26113042.
- Arensman M. D., Telesca D., Lay A. R., Kershaw K. M., Wu N., Donahue T. R., Dawson D. W. The CREB-binding protein inhibitor ICG-001 suppresses pancreatic cancer growth. (англ.) // Molecular cancer therapeutics. — 2014. — Vol. 13, no. 10. — P. 2303—2314. — doi:10.1158/1535-7163.MCT-13-1005. — PMID 25082960.
- Unzue A., Xu M., Dong J., Wiedmer L., Spiliotopoulos D., Caflisch A., Nevado C. Fragment-Based Design of Selective Nanomolar Ligands of the CREBBP Bromodomain. (англ.) // Journal of medicinal chemistry. — 2015. — doi:10.1021/acs.jmedchem.5b00172. — PMID 26043365.
- Xu M., Unzue A., Dong J., Spiliotopoulos D., Nevado C., Caflisch A. Discovery of CREBBP Bromodomain Inhibitors by High-Throughput Docking and Hit Optimization Guided by Molecular Dynamics. (англ.) // Journal of medicinal chemistry. — 2015. — doi:10.1021/acs.jmedchem.5b00171. — PMID 26125948.
- Petrij F., Giles R. H., Dauwerse H. G., Saris J. J., Hennekam R. C., Masuno M., Tommerup N., van Ommen G. J., Goodman R. H., Peters D. J. Rubinstein-Taybi syndrome caused by mutations in the transcriptional co-activator CBP. (англ.) // Nature. — 1995. — Vol. 376, no. 6538. — P. 348—351. — doi:10.1038/376348a0. — PMID 7630403.
- Kumar S., Suthar R., Panigrahi I., Marwaha R. K. Rubinstein-Taybi syndrome: Clinical profile of 11 patients and review of literature. (англ.) // Indian journal of human genetics. — 2012. — Vol. 18, no. 2. — P. 161—166. — doi:10.4103/0971-6866.100751. — PMID 23162289.
- Huh R., Cho S. Y., Kim J., Ki C. S., Jin D. K. Letter to the Editor: A Novel Mutation in the CREBBP Gene of a Korean Girl with Rubinstein-Taybi syndrome. (англ.) // Annals of clinical and laboratory science. — 2015. — Vol. 45, no. 4. — P. 458—461. — PMID 26275701.
- Rusconi D., Negri G., Colapietro P., Picinelli C., Milani D., Spena S., Magnani C., Silengo M. C., Sorasio L., Curtisova V., Cavaliere M. L., Prontera P., Stangoni G., Ferrero G. B., Biamino E., Fischetto R., Piccione M., Gasparini P., Salviati L., Selicorni A., Finelli P., Larizza L., Gervasini C. Characterization of 14 novel deletions underlying Rubinstein-Taybi syndrome: an update of the CREBBP deletion repertoire. (англ.) // Human genetics. — 2015. — Vol. 134, no. 6. — P. 613—626. — doi:10.1007/s00439-015-1542-9. — PMID 25805166.
- Vizmanos J. L., Larráyoz M. J., Lahortiga I., Floristán F., Alvarez C., Odero M. D., Novo F. J., Calasanz M. J. t(10;16)(q22;p13) and MORF-CREBBP fusion is a recurrent event in acute myeloid leukemia. (англ.) // Genes, chromosomes & cancer. — 2003. — Vol. 36, no. 4. — P. 402—405. — doi:10.1002/gcc.10174. — PMID 12619164.
- Gupta A., Patnaik M. M., Naina H. V. MYST3/CREBBP Rearranged Acute Myeloid Leukemia after Adjuvant Chemotherapy for Breast Cancer. (англ.) // Case reports in oncological medicine. — 2014. — Vol. 2014. — P. 361748. — doi:10.1155/2014/361748. — PMID 25548695.
- Bordonaro M., Lazarova D. L. CREB-binding protein, p300, butyrate, and Wnt signaling in colorectal cancer. (англ.) // World journal of gastroenterology. — 2015. — Vol. 21, no. 27. — P. 8238—8248. — doi:10.3748/wjg.v21.i27.8238. — PMID 26217075.
- Qin Y., Chen W., Xiao Y., Yu W., Cai X., Dai M., Xu T., Huang W., Guo W., Deng W., Wu T. RFPL3 and CBP synergistically upregulate hTERT activity and promote lung cancer growth. (англ.) // Oncotarget. — 2015. — Vol. 6, no. 29. — P. 27130—27145. — PMID 26318425.
- Gao Y., Geng J., Hong X., Qi J., Teng Y., Yang Y., Qu D., Chen G. Expression of p300 and CBP is associated with poor prognosis in small cell lung cancer. (англ.) // International journal of clinical and experimental pathology. — 2014. — Vol. 7, no. 2. — P. 760—767. — PMID 24551300.
- Xia Z., Guo M., Liu H., Jiang L., Li Q., Peng J., Li J. D., Shan B., Feng P., Ma H. CBP-dependent Wnt/β-catenin signaling is crucial in regulation of MDR1 transcription. (англ.) // Current cancer drug targets. — 2015. — PMID 25968898.
- Lee J. W., Park H. S., Park S. A., Ryu S. H., Meng W., Jürgensmeier J. M., Kurie J. M., Hong W. K., Boyer J. L., Herbst R. S., Koo J. S. A Novel Small-Molecule Inhibitor Targeting CREB-CBP Complex Possesses Anti-Cancer Effects along with Cell Cycle Regulation, Autophagy Suppression and Endoplasmic Reticulum Stress. (англ.) // Public Library of Science ONE. — 2015. — Vol. 10, no. 4. — P. e0122628. — doi:10.1371/journal.pone.0122628. — PMID 25897662.
- Song H., Moon M., Choe H. K., Han D. H., Jang C., Kim A., Cho S., Kim K., Mook-Jung I. Aβ-induced degradation of BMAL1 and CBP leads to circadian rhythm disruption in Alzheimer's disease. (англ.) // Molecular neurodegeneration. — 2015. — Vol. 10, no. 1. — P. 13. — doi:10.1186/s13024-015-0007-x. — PMID 25888034.
- Rouaux C., Loeffler J. P., Boutillier A. L. Targeting CREB-binding protein (CBP) loss of function as a therapeutic strategy in neurological disorders. (англ.) // Biochemical pharmacology. — 2004. — Vol. 68, no. 6. — P. 1157—1164. — doi:10.1016/j.bcp.2004.05.035. — PMID 15313413.
- Korzus E., Rosenfeld M. G., Mayford M. CBP histone acetyltransferase activity is a critical component of memory consolidation. (англ.) // Neuron. — 2004. — Vol. 42, no. 6. — P. 961—972. — doi:10.1016/j.neuron.2004.06.002. — PMID 15207240.
- Crisafulli C., Chiesa A., Han C., Lee S. J., Shim D. S., Balzarro B., Andrisano C., Sidoti A., Patkar A. A., Pae C. U., Serretti A. Possible influence of CREB1, CREBBP and CREM variants on diagnosis and treatment outcome in patients with schizophrenia. (англ.) // Neuroscience letters. — 2012. — Vol. 508, no. 1. — P. 37—41. — doi:10.1016/j.neulet.2011.12.013. — PMID 22198373.
- Crisafulli C., Shim D. S., Andrisano C., Pae C. U., Chiesa A., Han C., Patkar A. A., Lee S. J., Serretti A., De Ronchi D. Case-control association study of 14 variants of CREB1, CREBBP and CREM on diagnosis and treatment outcome in major depressive disorder and bipolar disorder. (англ.) // Psychiatry research. — 2012. — Vol. 198, no. 1. — P. 39—46. — doi:10.1016/j.psychres.2011.08.022. — PMID 22386572.
- Kumar D., Deb I., Chakraborty J., Mukhopadhyay S., Das S. A polymorphism of the CREB binding protein (CREBBP) gene is a risk factor for addiction. (англ.) // Brain research. — 2011. — Vol. 1406. — P. 59—64. — doi:10.1016/j.brainres.2011.05.048. — PMID 21752352.
- Malvaez M., Mhillaj E., Matheos D. P., Palmery M., Wood M. A. CBP in the nucleus accumbens regulates cocaine-induced histone acetylation and is critical for cocaine-associated behaviors. (англ.) // The Journal of neuroscience : the official journal of the Society for Neuroscience. — 2011. — Vol. 31, no. 47. — P. 16941—16948. — doi:10.1523/JNEUROSCI.2747-11.2011. — PMID 22114264.
- Xiang A., Ren F., Lei X., Zhang J., Guo R., Lu Z., Guo Y. The hepatitis B virus (HBV) core protein enhances the transcription activation of CRE via the CRE/CREB/CBP pathway. (англ.) // Antiviral research. — 2015. — Vol. 120. — P. 7—15. — doi:10.1016/j.antiviral.2015.04.013. — PMID 25936964.
- Ziady A. G., Sokolow A., Shank S., Corey D., Myers R., Plafker S., Kelley T. J. Interaction with CREB binding protein modulates the activities of Nrf2 and NF-κB in cystic fibrosis airway epithelial cells. (англ.) // American journal of physiology. Lung cellular and molecular physiology. — 2012. — Vol. 302, no. 11. — P. 1221—1231. — doi:10.1152/ajplung.00156.2011. — PMID 22467641.
Литература
- Marcello A., Zoppé M., Giacca M. Multiple modes of transcriptional regulation by the HIV-1 Tat transactivator. (англ.) // IUBMB life. — 2001. — Vol. 51, no. 3. — P. 175—181. — doi:10.1080/152165401753544241. — PMID 11547919.
- Matt T. Transcriptional control of the inflammatory response: a role for the CREB-binding protein (CBP). (англ.) // Acta medica Austriaca. — 2002. — Vol. 29, no. 3. — P. 77—79. — PMID 12168567.
- Combes R., Balls M., Bansil L., Barratt M., Bell D., Botham P., Broadhead C., Clothier R., George E., Fentem J., Jackson M., Indans I., Loizu G., Navaratnam V., Pentreath V., Phillips B., Stemplewski H., Stewart J. An assessment of progress in the use of alternatives in toxicity testing since the publication of the report of the second FRAME Toxicity Committee (1991). (англ.) // Alternatives to laboratory animals : ATLA. — 2002. — Vol. 30, no. 4. — P. 365—406. — PMID 12234245.
- Minghetti L., Visentin S., Patrizio M., Franchini L., Ajmone-Cat M. A., Levi G. Multiple actions of the human immunodeficiency virus type-1 Tat protein on microglial cell functions. (англ.) // Neurochemical research. — 2004. — Vol. 29, no. 5. — P. 965—978. — PMID 15139295.
- Kino T., Pavlakis G. N. Partner molecules of accessory protein Vpr of the human immunodeficiency virus type 1. (англ.) // DNA and cell biology. — 2004. — Vol. 23, no. 4. — P. 193—205. — doi:10.1089/104454904773819789. — PMID 15142377.
- Greene W. C., Chen L. F. Regulation of NF-kappaB action by reversible acetylation. (англ.) // Novartis Foundation symposium. — 2004. — Vol. 259. — P. 208—217. — PMID 15171256.
- Liou L. Y., Herrmann C. H., Rice A. P. HIV-1 infection and regulation of Tat function in macrophages. (англ.) // The international journal of biochemistry & cell biology. — 2004. — Vol. 36, no. 9. — P. 1767—1775. — doi:10.1016/j.biocel.2004.02.018. — PMID 15183343.
- Pugliese A., Vidotto V., Beltramo T., Petrini S., Torre D. A review of HIV-1 Tat protein biological effects. (англ.) // Cell biochemistry and function. — 2005. — Vol. 23, no. 4. — P. 223—227. — doi:10.1002/cbf.1147. — PMID 15473004.
- Bannwarth S., Gatignol A. HIV-1 TAR RNA: the target of molecular interactions between the virus and its host. (англ.) // Current HIV research. — 2005. — Vol. 3, no. 1. — P. 61—71. — PMID 15638724.
- Le Rouzic E., Benichou S. The Vpr protein from HIV-1: distinct roles along the viral life cycle. (англ.) // Retrovirology. — 2005. — Vol. 2. — P. 11. — doi:10.1186/1742-4690-2-11. — PMID 15725353.
- Gibellini D., Vitone F., Schiavone P., Re M. C. HIV-1 tat protein and cell proliferation and survival: a brief review. (англ.) // The new microbiologica. — 2005. — Vol. 28, no. 2. — P. 95—109. — PMID 16035254.
- Hetzer C., Dormeyer W., Schnölzer M., Ott M. Decoding Tat: the biology of HIV Tat posttranslational modifications. (англ.) // Microbes and infection / Institut Pasteur. — 2005. — Vol. 7, no. 13. — P. 1364—1369. — doi:10.1016/j.micinf.2005.06.003. — PMID 16046164.
- Peruzzi F. The multiple functions of HIV-1 Tat: proliferation versus apoptosis. (англ.) // Frontiers in bioscience : a journal and virtual library. — 2006. — Vol. 11. — P. 708—717. — PMID 16146763.