Массовое вымирание
Ма́ссовые вымира́ния — глобальные катастрофы в истории Земли, когда высокая (по сравнению с фоновым уровнем) доля видов большого числа высших таксонов вымирала в продолжение короткого по геологическим масштабам времени. Общепринятая в настоящий момент концепция разработана в 1980-х годах американскими палеонтологами Д. Сепкоски и Д. Раупом[1].
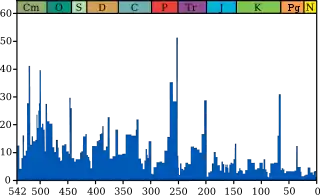
На протяжении фанерозоя (последние 540 миллионов лет) имело место пять крупных массовых вымираний и порядка 20 менее масштабных[прим. 1]. Последнее массовое вымирание произошло примерно 65 миллионов лет назад и было не самым значительным, однако оно наиболее известно из-за вымирания динозавров. Наикрупнейшее же из массовых вымираний (так называемое «Великое вымирание») 250 млн лет назад уничтожило 90 % существовавшего тогда биоразнообразия[2].
Основные гипотезы о причинах массовых вымираний — вулканические явления (трапповый магматизм) планетарного масштаба и импактные события[3].
В ряде работ в динамике вымираний обнаружены циклы различной продолжительности[4][5]; другие авторы отрицают их[6]. Эту возможную цикличность связывают чаще всего с космическими периодическими процессами[7][8].
История изучения

Исследование феномена глобального исчезновения видов организмов имеет довольно давнюю историю.
Первым в этой области был французский натуралист, барон Жорж Кювье, занимавшийся изучением вымерших организмов и разработавший в начале XIX века теорию катастрофизма. Она предполагала, что те или иные виды организмов господствовали в каждый геологический период, который затем завершался их резким исчезновением — так называемой революцией, приводящей к смене другими видами, — вследствие резких и внезапных «катастроф». Эволюционисты, начиная с Ч. Дарвина, не принимали этой теории, полагая, что изменение видового состава происходило медленно и постепенно. В то время, однако, считалось, что возраст Земли составляет всего несколько тысяч лет, поэтому для Кювье оправданно было предполагать необходимость таких переворотов: длительные геологические процессы просто не успели бы столь существенно изменить Землю за такое короткое время[9].
В 1920-е годы советский палеонтолог Д. Н. Соболев сформулировал[10] теорию и сценарий биотического кризиса. Эволюция, согласно его концепции, представляла собой последовательность «волн жизни», регулярных скачкообразных смен флор и фаун, вызванных геологическими катастрофами— горообразованием и ростом вулканической активности[11].
Конкретно термин «массовое вымирание» ввёл в русскоязычную литературу советский геолог Б. Л. Личков[12]. Он разработал гипотезу, объединившую соболевские «волны жизни» с «переворотами» Кювье[прим. 2], о циклах горообразования, связанных с изменением скорости вращения Земли[13], которые хронологически совпадали с «волнами жизни» и продолжались в среднем 60—70 млн лет — в общей сложности 6 циклов, начиная с кембрия[14]. При этом «волны жизни» растительности предшествовали таковым для животного мира. Цикл состоит из ледниковой, умеренной и ксеротермической фаз[12][13]. Начало цикла, знаменующееся новой тектонической диастрофой, означало одновременно революцию в органическом мире, рождавшую новую волну жизни[14]. Короткая первая фаза сменялась продолжительной умеренной, когда органический мир при постоянстве условий, умеренном климате и обилии пищевых ресурсов обретал состояние равновесия; интенсивность видообразования постепенно снижалась. В ксеротермическую фазу из-за резкого сокращения запасов воды и пищи происходила массовая гибель животных, особенно «менее пластичных форм». Однако ксерофитные фазы были также короткими — от нескольких сотен до нескольких тысяч лет — и за ними следовали новые вертикальные движения земной коры с образованием складок, влекшие за собой увеличение объёма ресурсов[12][13].
В западной литературе, возможно, впервые термин появляется в работах Нормана Ньюэлла в 1950-e — 1960-е годы[9][15]. Он связывал периодические биологические революции с колебаниями уровня Мирового океана[16]. В этот же период в ряде других работ подтверждалось существование кризисов в истории органической жизни Земли, между тем, немецкий палеонтолог Отто Шиндевольф и советские астрофизики В. И. Красовский и И. С. Шкловский считали их следствием воздействия внеземных факторов, хотя эти теории во многом были чисто спекулятивными[14][16]. Советский палеонтолог Л. Ш. Давиташвили, хотя и признавал роль внешних абиотических катастрофических факторов, считал вымирание естественным эволюционным процессом, представляющим результат естественного отбора — конкурентного вытеснения менее высокоорганизованных существ более высокоорганизованными и лучше приспособленными[14][16][17]. А в 1973 году Ли ван Вален сформулировал гипотезу Чёрной королевы, согласно которой вероятность вымирания крупных групп организмов является постоянной в пределах группы и случайной среди групп на протяжении длительных отрезков времени[18].


Важной вехой стала публикация в 1980 году открытия группой Луиса Альвареса свидетельств столкновения Земли с крупным космическим объектом — астероидом или кометой — на рубеже мезозоя и кайнозоя, на основании чего был сделан вывод о том, что именно это послужило причиной знаменитого вымирания динозавров[16]. Это перевело проблему из сугубо спекулятивной плоскости в область реальных научных исследований, включающих экспериментальные[15].
В 1982 году палеонтологи Джек Сепкоски и Дэвид Рауп из Чикагского университета произвели статистический анализ палеонтологических данных по вымиранию морских животных на протяжении фанерозоя (последние 540 млн лет — так как для более ранних времён данные практически недоступны, а эволюционная динамика могла быть существенно другой[19]) на материале 3300 семейств[15][16]. На основании этого они выделили[1] пять крупных массовых вымираний биологических видов и около 20 менее масштабных, когда погибало примерно 20 % форм жизни. В общей сложности вымерло 2400 семейств, а наиболее резкое вымирание приходилось на конец перми[16]. Чуть позже эти американские учёные пришли к выводу о наличии в истории массовых вымираний периодичности.
В эти годы феномен глобальных биосферных кризисов начал изучаться гораздо более интенсивно. Так, если на протяжении 20 лет до 1954 года ежегодно в среднем выходило всего 1—3 работы по данной теме, в период с 1954 по 1957 год это число возросло до 7,25, к концу 1960-х годов достигло 23, к началу 1970-х годов — 45, к концу 1970-х годов — 80, в начале 1980-х годов превзошло 150, к концу 1980-х годов достигло 250 и в начале XXI века публиковалось уже более 330 работ ежегодно[15]. Большую роль сыграло накопление объёма палеонтологических данных о существовавших на Земле видах. Первая максимально полная «Палеонтологическая летопись» была опубликована в 1967 году Лондонским геологическим обществом[20] и содержала информацию о времени (с разрешением до уровня яруса) существования 2924 семейств (или таксонов эквивалентного уровня). Второе издание списка всех известных в ископаемом состоянии семейств микробов, водорослей, грибов, протистов, растений и животных вышло в 1993 году[21], он содержал 30 видов, 122 класса, 701 отряд и 7186 семейств. База Д. Сепкоски в последней версии 2002 г.[22] содержит порядка 36 000 родов морских организмов. С 1998 года функционирует в онлайн-формате публичная палеобиологическая база данных[23], на конец 2010-х годов над ней постоянно работают около 400 учёных из более чем 130 научных организаций в 24 странах, и она содержит информацию о более чем 370 000 таксонах, в том числе более чем 150 000 видах.
Известные вымирания
Критерии классификации

Согласно концепции Сепкоски[24],
|
массовое вымирание есть существенный рост интенсивности вымирания (то есть прерывания конкретной ветви филогенетического дерева) более чем одного географически широко распространённого высшего таксона в течение короткого по геологическим масштабам времени, в результате которого снижается, по крайней мере временно, их общее разнообразие.
|
A mass extinction is any substantial increase in the amount of extinction (i.e., lineage termination) suffered by more than one geographically wide-spread higher taxon during a relatively short interval of geologic time, resulting in an at least temporary decline in their standing diversity. |
В таком определении, как признаёт и сам Сепкоски, присутствует неопределённость в количественном плане. Так, основными критериями принадлежности события к массовым вымираниям являются:
- Короткая по геологическим меркам продолжительность[25][26]. Это отличает массовые вымирания от долговременных эволюционных трендов, предполагающих постепенное вытеснение одних видов другими[15]. Неизбежна, однако, трудность, связанная с пределом точности датировки: события продолжительностью 100 лет и 100 000 лет будут неразличимы; при самых благоприятных обстоятельствах можно говорить о пределе разрешения в несколько тысяч лет. Палеонтология имеет дело с временными интервалами порядка миллионов лет, так что любое событие, произошедшее при его рассмотрении в таких временных масштабах мгновенно и имеющее существенные последствия, можно считать катастрофическим[27]. Существуют, однако, и другие проблемы с датировкой вымираний: ископаемые остатки, имеющие меньший возраст, сохраняются лучше и в большем объёме, из-за чего наблюдается сдвиг в сторону существующих и существовавших в недалёком прошлом организмов[28], а неполнота стратиграфических данных, в частности, невозможность зарегистрировать последнее появление представителей того или иного таксона, может привести к неверной оценке динамики вымирания, когда оно кажется более плавным и произошедшим раньше, чем на самом деле (эффект Синьора—Липпса)[29][30]. Фрагментарность палеонтологической летописи приводит к проблеме регистрации «псевдовымираний» ранее срока действительного исчезновения родов (или видов, семейств и т. д.)[16].
- Локальный максимум интенсивности вымирания. Не определён, однако, порог относительно некоего фонового уровня непрерывно происходящего исчезновения видов — фактически он устанавливается произвольно для каждого события[15], и вообще вопрос принципиального отличия массового вымирания от фонового не разрешён однозначно[31][32]. Далее, важно установить, как именно определяется интенсивность вымирания — величина, адекватно отражающая критичность событий для биосферы. Сепкоски и Рауп предлагают 4 мероопределения[33]:
- Полное вымирание — общее число семейств, исчезнувших в продолжение яруса.
- Относительное (процентное) вымирание — полное вымирание, делённое на число семейств, существовавших в продолжение рассматриваемого яруса;
- Темп полного вымирания — отношение число вымерших в течение яруса семейств (полного вымирания) к продолжительности этого яруса.
- Темп относительного вымирания — отношение темпа полного вымирания к текущему разнообразию таксонов (семейств).
От того, какой параметр используется для характеристики, во многом зависят и результаты анализа данных[34]. А неизбежные погрешности при определении продолжительности временных интервалов (в частности, ярусов) также неминуемо приводят к неточностям в оценках, содержащих такие величины[30][35].
- Широкое разнообразие таксонов, подвергшихся воздействию[25][26]: это должно быть событием более масштабным, чем массовая гибель представителей одной группы, хотя конкретное число также строго не определено[15].
- Высокий ранг пострадавших таксонов[15] — часто говорят о родах или семействах[1][34], хотя некоторые специалисты считают более корректным вести речь об отрядах и более высоких рангах таксонов, тогда как для семейств и ниже биоразнообразие может изменяться со временем совсем иначе[15]. Вообще когда речь идёт о числе вымерших видов, зачастую это лишь косвенные выводы, сделанные, например, с помощью кривых разрежения для более крупных таксонов[30].
- Широкий охваченный ареал — строгих значений также нет, но для критичных экологических последствий это должна быть территория не менее половины земного шара[27]; хотя это вовсе не обязательно должен быть глобально однородный рост интенсивности вымирания — разные территории могут быть затронуты в разной мере[15].
- Высокая доля вымерших видов[25][26]: она может сильно варьироваться для различных таксонов более высокого ранга, что важно учитывать при оценке[30].
Пользуясь этими критериями, сами Сепкоски и Рауп выделяют 5 крупнейших вымираний[1]. Британский палеонтолог М. Бентон, основываясь на анализе составленной под его руководством «Палеонтологической летописи»[21], в целом подтверждает эти выводы, отмечая, однако, важность того, какой параметр используется для сравнения[34]. Однако в литературе встречались и другие точки зрения на этот счёт. Так, в более поздних работах самого Дж. Сепкоски некоторые из событий «большой пятёрки» разделяются на 2 и более биотических кризисов, и таким образом насчитывается 11 вымираний. Палеонтолог С. М. Стэнли из США исключает (из-за неточности данных о времени и числе вымерших видов) одно из вымираний «большой пятёрки», но добавляет два других[30]. Ещё один американский палеоэколог Р. Бамбах утверждает, что если формально применять критерии Сепкоски, на основании анализа его же палеонтологической базы можно выделить 18 пиков по темпу и интенсивности исчезновения видов, и все они будут подходить под определение[15]. Если же судить по абсолютной интенсивности вымираний, статистически над фоновым уровнем выделяются, по его мнению, только три из «большой пятёрки», другие же два выраженных минимума биоразнообразия могут быть обусловлены в той же мере снижением интенсивности обратного процесса — видообразования[15][36]. Некоторые исследователи и вовсе считают по-настоящему массовым вымиранием только одно событие — на рубеже перми и триаса[36]. Наконец, палеонтолог из США Дж. Р. Макги мл. выделяет те же 11 событий, но в немного другой последовательности по числу вымерших видов, предлагая при этом также отличную от этой отдельную классификацию по введённому им критерию тяжести экологических последствий[37].
Крупнейшие
Классическая «большая пятёрка» вымираний, выделенная Сепкоски и Раупом[1]:
- 450—443 млн лет назад — ордовикско-силурийское вымирание — уступало, согласно ряду классификаций, по масштабности только пермскому[30][38][39], прервав происходившее в этот период интенсивное видообразование. Происходило в 2 этапа. Первый, более чётко выраженный, в первые тысячелетия хирнатского яруса, ознаменовался исчезновением многих видов пелагических организмов, в особенности первичного фито- и зоопланктона (представленного граптолитами и хитинозоями) — в общей сложности вымерло 40 % всех родов, — и, вероятно, совпадал (но не факт, что был связан) с началом интенсивной 1,9-миллионлетней фазы позднеордовикского оледенения. Второй же соответствовал моменту завершения оледенения и подъёму уровня Мирового океана; он был менее избирателен в своей направленности, уничтожив 31 % родов, в частности, многие виды, пережившие первый этап, в том числе большую часть конодонтов, а также организмы, приспособленные в процессе оледенения к жизни в холодной воде, — они сменились примитивными экосистемами[3][40][15][37]. Всего в ходе этого события исчезло (по разным данным[прим. 3]) 22—27 % всех семейств и 43—61 % всех родов, в частности, 72—86 % видов и более 100 семейств морских[прим. 4] беспозвоночных, в том числе 2/3 всех семейств плеченогих (строфоменид, ринхонеллид и ортид), наутилоидей, криноид, конодонтов, мшанок и семейств трилобитов, питавшихся как первичным планктоном, так и детритом; довольно сильно пострадали некоторые двустворчатые (нукулоиды), ракушковые (палеокопиды) и брюхоногие[3][15][38][30][37]. Между тем, несмотря на довольно высокие значения этих показателей, экологические последствия были не так критичны: необратимы они были фактически только для трилобитов[30], которые в тот период даже не играли особо важной роли[37]; потери для атрипид, узкоротых мшанок и строматопороид составили менее 35 %[прим. 5], а по радиоляриям и текстуляриидам вообще нет данных об их исчезновении в этот период[15][37]. По некоторым гипотезам, это вымирание было вызвано гамма-всплеском[41], однако доказательное подтверждение этого проблематично[3].
.jpg.webp)
- 372 млн лет назад[прим. 6] — девонское вымирание, которое представляло собой серию эпизодов сокращения численности таксонов на протяжении практически всего среднего, верхнего и даже частично нижнего девона. Даже фоновый уровень вымирания девонского периода составлял 18,8 % родов[15]. Как правило, выделяют[42]:
- Злиховское и далейское события в эмсском ярусе — отметились вымиранием 75 % видов гониатитов, которые только начали активно появляться[3].
- Хотечское событие на границе эмсского и эйфельского ярусов[3].
- Событие Качак в позднем эйфеле (возможно, двухэтапное) — ознаменовалось существенным сокращением численности головоногих (гониатитов, наутилоидов), конодонтов и тентакулитов[3]; также пострадали некоторые семейства плеченогих и кораллов[37].
- 388—385 млн лет назад — Таганикский кризис в живетском ярусе[прим. 7] — ударил не только по гониатитам, но и таксонам, обитавшим в придонной области, особенно в низких широтах, — кораллам и строматопороидам — организмам, формировавшим рифы[3]. С этим периодом связана так называемая великая девонская перемена — смена эндемичных фаун различных областей Лавруссии и Гондваны инвазивными видами космополитной фауны предположительно из-за подъёмов уровня Мирового океана[37]. Наибольший процент вымирания отмечен для теребратулид, наутилид, трилобитов (таких как кальмонииды), стебельчатых иглокожих (криноидей и бластоидей), плакодермов и филлокарид — более 57 %, что вдвое превышает общий средний показатель этого события. Гораздо меньше пострадали фузулиниды, узкоротые и брюхоногие[15][37].
- 378—374 млн лет назад — Нижнее и верхнее событие Келлвассера (франско-фаменское), которые часто и подразумеваются под девонским вымиранием — одним из «большой пятёрки» Сепкоски, хотя и он сам в том числе позднее называл его относительно некрупным кризисом[39]: вымерло 35 %, а под другим данным — всего 16—20 % родов[30], и минимум биоразнообразия обусловлен в большей степени снижением видообразования[15]. Однако с другой стороны, разрушение рифовых экосистем, до этого периода переживавших расцвет, а после так и не восстановившихся до такого масштаба лишь через 25 млн лет, ставит события Келлвассера на высокое место по серьёзности экологических последствий[3][30][37]. Вымирание последних гомоктенид ознаменовало полное исчезновение класса тентакулитов — основной составляющей девонского зоопланктона[43]. Первый этап вымирания длился 400 тыс. лет и совпадал с понижением уровня Мирового океана, а второй — 50 тыс. лет, соответствовал подъёму уровня моря и ударил уже по сухопутным организмам. Помимо основных строителей рифов — строматопороид и табулятов — в ходе франско-фаменского кризиса вымерло более 2/3 родов шестилучевых губок, ругоз, строфоменид, трилобитов, плакодермов и остракодермов[37]. В то же время менее четверти состава потеряли радиолярии, узкоротые, беззамковые брахиоподы, ракушковые и конодонты[15].
- 364—359 млн лет назад — Хангенбергское событие в самом конце позднего девона, практически на рубеже карбона — существенно замедлило появление новых видов и было связано с вымиранием конодонтов, трилобитов, цефалоподов (в частности, аммоноид), брахиоподов (в особенности ринхонеллид), пластинокожих и костных рыб, строматопороид и кораллов; уничтожило в общей сложности 21—31 % морских родов и 16 % семейств[3][15][39]. При этом для узкоротых, брюхоногих, двустворчатых и криноид уровень вымирания был ниже, чем средний фоновый по периоду[15].
- Всего в ходе девонского вымирания исчезло 19 % всех семейств и 50 % всех родов. Оно затронуло в основном гидробионтов: численность их видов сократилась на 70 %, в частности, практически полностью исчезли организмы, формировавшие коралловые рифы[38]. Последнее необратимо изменило глобальную экосистему Мирового океана. События девонского периода повлекли кризис и на суше — для растений, а также существенно затронули только появившихся к тому времени лопастепёрых рыб и четвероногих позвоночных[43].

- 253—251 млн лет назад — «великое» пермское вымирание, самое массовое вымирание из всех, приведшее к исчезновению 57 % всех семейств и 83 % всех родов, более 90 % (по некоторым оценкам, даже 95 %) морских видов (наибольшие потери зафиксированы для фузулинид, ругоз, ракушковых и различных классов плеченогих; также пострадали мшанки, гастроподы, губки и сидячие двустворчатые[30][15]) и более 70 % видов наземных позвоночных[38]. При этом поражающие механизмы действовали избирательно: среди сидячих организмов с массивным карбонатным скелетом вымерло до 90 % родов, для обладателей менее массивных скелетов, в зависимости от возможности метаболически контролировать диффузионный газообмен, этот показатель составлял от 28 % до 78 %, а для таксонов с лёгким или вообще отсутствующим карбонатным скелетом — всего 10 %[15]; также показано, что более сильному вымиранию подверглись таксоны, характеризовавшиеся бо́льшим (на начало кризиса) разнообразием и более высоким уровнем фонового вымирания[30]. Примечательно, что пермская катастрофа произошла за очень короткий по геологическим меркам промежуток времени — менее 200 тыс. лет[прим. 8][3][15]. После неё, судя по окаменелостям, например, от всего разнообразия наземных пресмыкающихся, как и гидробионтов, осталось только по одному виду; на восстановление биоразнообразия наземных организмов понадобилось 50 млн лет, а морских — до 100 млн лет[38]. Хотя существуют альтернативные оценки, согласно которым потери составили только 62 % родов и 81 % видов (так, для агглютинированных фораминифер, спумеллярий и глубоко зарывающихся двустворчатых был характерен процент вымирания ниже среднего[15]), в частности, более 90 отрядов (90 % их общей тогдашней численности) и 220 семейств высших морских животных успешно пережили этот кризис[30], он единодушно признаётся всеми исследователями крупнейшей биосферной катастрофой в истории Земли. Все экологические связи были разрушены и впоследствии выстраивались заново[15].
- 208—200 млн лет назад — триасовое вымирание, — в результате которого вымерло 23 % всех семейств и 48 % всех родов, по меньшей мере, половина известных сейчас видов, живших на Земле в то время, в частности, 20 % морских видов (в наибольшей степени — двустворчатые, конодонты, известковые губки, каменистые кораллы, такие отряды плеченогих как спирифериды и теребратулиды, подклассы головоногих наутилоиды и аммоноиды[15]), некоторые растения, множество пресмыкающихся, а также последние крупные земноводные, что, между прочим, освободило экологическую нишу для динозавров[38]. Между тем, некоторые учёные[44][45] сомневаются в том, что на рубеже триаса и юры произошло именно массовое вымирание, полагая, что некоторые виды исчезали лишь локально, а другие — постепенно в течение всего рэтского и даже предшествующего норийского яруса — средний фоновый уровень вымирания в триасовом периоде вообще был довольно высок (34 %). Приводится аргумент, что неизвестно, когда точно произошли (в частности, одновременно ли) кризисы растений и позвоночных на суше и морских видов[30][3][15]. К тому же, как и для фанско-фаменского кризиса, спад биоразнообразия может являться естественной убылью из-за снижения видообразования[15].
- 65,5 млн лет назад — мел-палеогеновое вымирание — последнее массовое вымирание, уничтожившее 17 % всех семейств и 39—47 % всех родов, 68—75 % всех видов[30], в том числе и динозавров. Исчезли также последние аммониты и белемниты, рудисты, отряды фораминифер милиолиды и глобигериниды, многие диатомеи, динофиты, теребратулиды, известковые и шестилучевые губки, рыбы и морские пресмыкающиеся (плезиозавры и мозазавры), а на суше — растения, птицы, ящерицы, змеи, насекомые[3][15][38]. По-видимому, для гидробионтов решающую роль сыграла форма питания: таксоны, питавшиеся взвешенными в толще воды фотосинтезирующими организмами, пострадали гораздо сильнее тех, кому для этого требовался осаждённый детрит[15]. Хотя на восстановление биоразнообразия потребовалось 10 млн лет, это вымирание всё же было не самым значительным, уступая по масштабности другим из «Большой пятёрки», тем не менее, оно вызывает большой интерес, поскольку форсировало эволюцию млекопитающих (в частности, и появление человека)[38]; оно уничтожило динозавров и, возможно, было вызвано падением метеорита, благодаря чему популярно в массовой культуре[3].
Недавние (за последние 100 млн. лет)
- 95—93 млн лет назад — сеномано-туронское пограничное биотическое событие в совокупности с предшествовавшим ему первым аптским бескислородным событием — резкое сокращение объёма кислорода, возможно, из-за подводных вулканических извержений. Исчезло около 27 % морских беспозвоночных — такие как теребратулиды и аммониты, также пострадали костные рыбы, плиозавриды и ихтиозавры, из наземных пострадали представители надсемейства Spinosauroidea и некоторые другие динозавры[46][47]. В общей сложности вымерло 25 % родов, что существенно превышает среднефоновый уровень мелового периода в 10 %[15].
- 37—34 млн лет назад — эоцен-олигоценовое вымирание — единственный глобальный кризис на протяжении всего кайнозоя (характеризовавшегося фоновым уровнем вымирания в 8 %), уничтоживший 15 % всех родов, в частности, более 30 % милиолид и морских ежей и от 20 до 30 % глобигеринид и роталиид, каменистых кораллов и десятиногих раков[15]. Он совпал и с важным событием на суше, названном «Великим переломом» — резким изменением видового состава млекопитающих на территории Европы, которое представляло собой, в частности, полное исчезновение родов палеотериев и аноплотериев и семейств Xiphodontidae и Amphimerycidae. Это было связано, вероятно, с падением метеорита[48], и/или похолоданием климата[49]. В этот период зафиксирован и отрицательный сдвиг изотопа углерода.
- 130—1 тыс. лет назад — позднечетвертичное вымирание, во многом связанное с деятельностью человека. Наиболее пострадала мегафауна — наземные животные массой более 45 кг, являющиеся удобной добычей для человека. За этот относительно короткий (в плане видообразования) период вымерли 177 известных видов наземных млекопитающих массой более 10 кг, в том числе 154 вида массой более 45 кг[50], погибло 65 % известных науке видов мегафауны[51].
- 12 тыс. лет назад - настоящее время. Некоторые учёные предлагают вести отсчёт этого вымирания с 1500 года н.э., либо с начала нашей эры, как начала истории современного человечества[52][53].
- Некоторые учёные придерживаются мнения, что мы живём во время шестого массового вымирания, причём являющегося следствием деятельности человека[38][54][53][52]. Оно получило название голоценового. Теория основана на оценках скорости исчезновения видов, которая примерно в 100 раз быстрее, чем фоновая скорость исчезновения видов между массовыми вымираниями[53][52]. Приверженцы экологического скептицизма[прим. 9] высказываются против этой гипотезы, считая что скорость исчезновения биологических видов сложно оценить из-за недостатка сведений[55]. Между тем, из-за экстенсивной деятельности человека по всему миру (обезлесение, преобразование природных экосистем в пашни, пастбища и антропогенные ландшафты, прямое уничтожение животных и растений, промышленная добыча рыбы в мировом океане, добыча полезных ископаемых, загрязнение окружающей среды) деградируют природные экосистемы на 75% суши, 40% мирового океана, 50% пресных вод. В настоящее время под угрозой исчезновения находятся 25 % всех видов животных и растений (в среднем по всем видам в мире). Вымирание угрожает более 40 процентам видов амфибий, более 30 процентам видов морских млекопитающих и рыб[56].
Прочие
- 521—509 млн лет назад — раннекембрийское вымирание — по разным теориям однократное, либо несколько последовательных событий в ботомском ярусе (Синское), и/или в тойонском ярусе, и/или на границе нижнего и среднего кембрия. Есть данные об исчезновении в этот период археоциатов и различных представителей т. н. проблематичной мелкораковинной фауны (например, анабаритид), однако это может быть просто следствием снижения в кембрии доступного объёма фосфатов, необходимых для сохранения скелетных остатков[3]. Также вымерло множество родов редлихиид и оленеллид, хиолитов, томмотиид[57]; в меньшей степени пострадали ракушковые и беззамковые двустворчатые[15]. Совокупные потери биоразнообразия составляли около 50 %, а по некоторым оценкам, до 70 %; правда, это значение есть процентное отношение от общего числа таксонов, которое на тот момент в абсолютном выражении было невелико[3][34].
- 500 млн лет назад — позднекембрийское вымирание в начале дресбахского яруса. Некоторые данные (окаменелости трилобитов) свидетельствуют о вымирании 40—65 % гидробионтов, но они обнаружены только в Северной Америке и Австралии, в то время как о глобальном распространении информации нет, и вообще ископаемых свидетельств настолько мало и они неточны, что это может быть их погрешностью. Между тем, в породах этого возраста достоверно зафиксирована[58] глобальная аномалия δ13C, что, вероятно, доказывает деструкцию биосферы[3][15].
- 432—424 млн лет назад — серия силурийских вымираний:
- 433,4 млн лет назад, на границе лландоверийского и венлокского отделов (теличского и шейнвудского ярусов) — иревикенское событие. Уничтожило 80 % конодонтов (48 из 60 видов) и граптолитов, 50 % (локально) трилобитов, значительно сократило число видов акритарх, хитинозоев, брахиоподов (хотя и не так много видов в абсолютном выражении, но они составляли существенную часть ракушечной биомассы) и кораллов[26][59], в то время как на мелководных строителей рифов повлияло мало[3].
- Событие Мульде в середине Гомерского яруса венлокского отдела — затронуло в основном пелагические организмы: до 95 % граптолитов (из 50 видов осталось 2—3) и радиолярий (выжило 2 вида из 28), 50 % акритархов, отчасти хитинозоев, цефалоподов и конодонтов, а также рыб[26].
- Событие Лау (лудфордское) в лудфордском ярусе[прим. 10] лудловского отдела — повлияло как на пелагическую, так и бентическую фауну, причём сильнее — на сообщества организмов[26]. Кризис начался, вероятно, в глубоководной среде и лишь затем распространился в более мелкие береговые моря[37]. Больше всего свидетельств о вымирании конодонтов (17 из 23 видов), однако известно, что существенную реорганизацию претерпели также морские позвоночные, наутилоиды, пентамериды и другие брахиоподы, кораллы, двустворчатые, хитинозои, остракоды, полихеты, щитковые и колючкозубые рыбы и акритархи[26][37]. Граптолиты же пострадали меньше, чем в более ранних событиях[3]. Между тем, отмечается, что освобождение соответствующих экологических ниш стимулировало появление и развитие новых классов рыб и плеченогих[37].
- Все три события силурийского периода, хотя и не относятся к крупным, тем не менее являются массовыми вымираниями: они совпадали с похолоданиями климата, изменением уровня Мирового океана и резкими колебаниями содержания изотопа 13C и 18O[3]; длились не более 200 тыс. лет, оказали воздействие на широкий ареал таксонов, занимавших широкий ареал мест обитания на всех океанических глубинах, — как планктон, так и нектон и бентос. Правда, число групп, в которых вымерла значительная доля видов, не так велико, экологические последствия были весьма существенны, к тому же, возможно, о некоторых видах просто недостаточно информации. Есть свидетельства проявления связанного с этими событиями эффекта лиллипута — характерного уменьшения размеров тела организмов, сопровождавшего все крупнейшие массовые вымирания[26].
- 326—322 млн лет назад — серпуховское вымирание вскоре после начала позднепалеозойского оледенения — относительно некрупное событие, затронувшее всего 15—30 % существовавших тогда родов, хотя последних было не так много: рифовые экосистемы не успели ещё, по некоторым оценкам, восстановиться после девонского кризиса[30]. Тем не менее, они пострадали настолько, что для возврата к раннесерпуховским объёмам производства карбонатов понадобилось 30 млн лет. Произошла коренная перестройка не только рифовых, но и бентических криноидных экосистем[37]. Да и в целом событие выделялось на фоне среднего уровня, характерного для нижнего карбона (21 % родов) — это сравнимо по масштабам с Хангенбергским событием. Вымерло более 40 % аммонитов, криноид, конодонтов и хрящевых рыб[15].
- 265—260 млн лет назад — гваделупское (кептенское) вымирание в середине пермского периода, которое ряд специалистов относительно недавно[60][61] начали отделять от крупнейшего эпизода в его конце, хотя мнения по поводу продолжительности и интенсивности существенно разнятся — от 24 % родов до 48 % в общей сложности в продолжение всего яруса (в любом случае, это выше среднефонового уровня пермского периода — 14,5 %)[3][15][30]. Тем не менее, ряд данных свидетельствует о том, что это событие сравнимо по масштабам экологического воздействия с «Большой пятёркой»: значительный урон был нанесён бентическим рифовым экосистемам (в частности, губкам и табулятам), которым потребовалось для восстановления после этого до 7 млн лет; существенной перестройке подверглись бентические крупные фораминиферовые фотосимбиотические системы — вымерла большая часть фузулинид (в частности, семейства неошвагеринид и вербеекинид), а оставшиеся фораминиферы сократились в размерах и морфологической сложности; наконец, типично палеозойские некробентические аммоноидные фауны (отряды гониатитид и пролеканитид), сменились пелагическими (отряд цератитов), характерными для мезозоя[37]. В этот период исчезли многие брахиоподы (строфомениды и ринхонеллиды), мшанки, двустворчатые, трилобиты и криноиды, а также сухопутные организмы — локально (на территории современного Китая) до 56 % видов растений и, вероятно, дейноцефалы[3][15][30]. В меньшей степени пострадали текстулярииды, известковые губки и ракушковые[15].
- 249 млн лет назад — поздне-нижнетриасовое (смитско-спэтское[прим. 11]) вымирание — относительно некрупное, но тем не менее важное событие, следовавшее за восстановлением после глобальной Пермской катастрофы и замедлившее его. Геологические данные свидетельствуют о характерных процессах, сопровождавших и вызывавших массовые вымирания: парниковом эффекте, дефиците питательных веществ (из-за нарушения апвеллинга) и кислорода, присутствии токсичных металлов[3][62].
- 230 млн лет назад — карнийское плювиальное событие[63] — относительно краткосрочный период обильных осадков, сменивший (и предварявший) засушливый климат, характеризовался сильными муссонами на Пангее и напоминал пермское вымирание, но в меньшем масштабе. Сильно пострадал морской нектон (головоногие и конодонты) и бентос (криноиды, морские гребешки, кораллы, фузулиниды), а на суше изменение видового состава флоры привело к исчезновению некоторых травоядных четвероногих[3].
- 186—179 млн лет назад — ранне-тоарское изменение[64], захватившее и часть плинсбахского яруса, — относительно несущественное по интенсивности (всего 5 % семейств и 20—25 % родов), однако глобальное событие, нанесшее значительный урон мелководным морским моллюскам[3], каменистым кораллам, ринхонеллидам и теребратулидам, аммонитам, костным рыбам и морским пресмыкающимся. Зафиксированы и характерные признаки биотического кризиса — резкий отрицательный скачок δ13C, аноксия[15].
Причины
Причины массовых вымираний являются предметом активных дискуссий. Их можно разделить на факторы, непосредственно влекущие исчезновение организмов (поражающие), и первичные (триггерные), являющиеся источником этих факторов[3][65]. Последние, в свою очередь, могут подразделяться на внешние и внутренние. В настоящий момент наибольшее подтверждение среди специалистов получили:
- Понижение уровня Мирового океана — исторически первая[66] гипотеза, объясняющая события массовых вымираний разрушением среды обитания основной массы бентоса в прибрежных водах из-за обмеления. Такие события могли быть следствием тектоники плит, а также таяния ледников и других изменений климата[6][67][68][69]. Впоследствии было выявлено, что хотя в периоды массовых вымираний и имели место колебания уровня моря, минимумы более или менее точно совпадают только с событиями конца гваделупского отдела и триасово-юрского рубежа; позднеордовикское и мел-палеогеновое вымирания соответствуют участкам вблизи локальных минимумов, однако уже на этапе роста; франско-фаменское и пермское вымирания, а также сеномано-туронское событие происходили во время повышения уровня Мирового океана; а для Хангенбергского кризиса и раннекембрийских событий вообще характерна резкая смена динамики. Таким образом, однозначной корреляции не прослеживается[70].
- Выбросы углекислого газа той или иной природы, вызывающие закисление Мирового океана и гиперкапнию. Резкое (до 10000—30000 ppm) повышение содержания CO2 в атмосфере приводит к быстрой гибели организмов из-за нарушения окислительных процессов при дыхании (эффекты Бора и Рута), однако увеличение даже на 200 ppm относительно среднего уровня при длительном воздействии снижает способность к росту, размножению и выживанию. Для примитивных видов в основании пищевой цепи понижение pH также затрудняет рост и размножение, кроме того, нарушает карбонатную биоминерализацию — как и рост концентрации CO2, приводящий к снижению содержания во внешней среде иона карбоната СО32−. У более же сложных организмов из-за взаимодействия с химическими рецепторами нарушается работа сенсорных систем, что снижает способность к распознаванию как потенциальной угрозы (хищников), так и возможных партнёров для спаривания. При этом, однако, факторы увеличения[прим. 12] кислотности и содержания углекислого газа не являются и не являлись губительными для всех видов поголовно: для некоторых из них они оказывались не столь существенными или даже выгодными, в том числе именно за счёт роста смертности среди конкурирующих особей[3]. Так, морские виды гораздо более чувствительны к гиперкапнии вследствие лучшей растворимости углекислого газа в воде; тогда как для наземных растений небольшой избыток CO2 даже полезен, однако при сильном росте его содержания происходит закисление почвы. В целом восприимчивость к этому фактору у различных видов сильно варьируется, чем объясняется, в частности, селективность крупнейшего пермского вымирания[65].
- Глобальное похолодание либо потепление[38]. При повышении температуры среды и невозможности миграции в район с привычными условиями организмы вынуждены увеличивать интенсивность своего аэробного метаболизма, причём это происходит экспоненциально — в 2 раза на каждые 10 °C[71], то есть резко возрастает потребность в кислороде, тогда как организм может быть неспособен получать его из среды в таком количестве. Так что, с одной стороны, это даёт преимущество менее активным видам, имеющим более медленный обмен веществ (таким как брахиоподы), с другой — виды с более быстрым метаболизмом (например, двустворчатые) способны легче подстраивать пределы возможного объёма поглощаемого кислорода. Возможно, играют роль также изначальные условия среды и предварительное воздействие температурного стрессора. На настоящий день нет однозначного мнения, насколько критичным является этот фактор и для каких именно видовых групп[3]. Тем не менее была выявлена выраженная временная корреляция между повышением температуры (парниковым эффектом), снижением биоразнообразия и интенсивностью вымирания[72], в частности, потепление имело место в крупнейших событиях массовых вымираний на рубеже мелового и палеогенового периода, в позднем девоне и на границе перми и триаса, а при ордовикско-силурийском вымирании имело место похолодание[3].
- Дефицит кислорода, обычно сопровождающий повышение температуры и непосредственно вызывающий при достаточно длительном (более 60 дней) воздействии гибель живых организмов[3]. Тем не менее, аноксия никогда не была глобальной, и даже в верхних слоях океана (а тем более в атмосфере) оставались участки с относительно приемлемыми условиями[65].
- Отравление сероводородом, избыток которого может быть, например, прямым следствием дефицита кислорода в среде в результате увеличения интенсивности анаэробного дыхания сульфатредуцирующих бактерий. Сероводород токсичен сам по себе практически для всех эукариотических клеток, а также вызывает рост концентрации метана в тропосфере и, как следствие, разрушение озонового слоя[3][65].
- Отравление токсичными металлами[3].
- Воздействие солнечной радиации из-за разрушения озонового слоя[3] — более фатально для наземных позвоночных, чем для примитивных морских организмов[65].
- Падение величины первичной продукции[73], которое некоторые исследователи связывают с выбросами в атмосферу пыли и аэрозолей, блокирующих солнечный свет, необходимый для фотосинтеза. Однако в этом случае наблюдался бы переизбыток кислорода, тогда как данные свидетельствуют о том, что в периоды крупнейших массовых вымираний имел место, наоборот, дефицит кислорода и избыток сульфидов — продуктов анаэробного дыхания. Более вероятно, что сыграло роль изменение состава морского фитопланктона: эвтрофикация гетеротрофных бактерий (в частности, сульфатредуцирующих микроорганизмов) и автотрофных (цианобактерий и зелёных серобактерий), вытеснивших более разнообразные по составу эукариотические водоросли, привело к дефициту производимых последними стеринов, из-за чего существенно пострадали многие беспозвоночные, такие как членистоногие и моллюски[65][74]. Такие процессы происходили в конце пермского, триасового и, скорее всего, ордовикского периодов, а также в позднем девоне, и они совпадали с периодами глобальной аноксии и эвксинии Мирового океана[74].
Между тем, ни один фактор, по-видимому, не был фатальным, и большинство специалистов приходят к выводу, что они действовали в комплексе, синергически[3][65][75][76].
Наиболее популярными версиями первичных причин являются:

- Столкновение с небесными телами. По своей природе импактные объекты могли быть околоземными астероидами — атонами, аполлонами и амурами; либо кометами из облака Оорта, которые могли приближаться к Земле при возмущении гравитационного поля каким-либо внешним воздействием, например, от звёзд или молекулярных облаков других звёздных систем, либо галактическими приливными силами[6]. Такие события могут приводить к повреждениям от ударной волны, цунами, землетрясениям, оползням, лесным пожарам, а также выбросам в атмосферу большого количества парниковых газов и пыли, что, в свою очередь, может повлечь разрушение озонового слоя, изменения климата (как потепление, так и похолодание), кислотные дожди, блокирование солнечного света («импактная зима») и закисление вод Мирового океана[3][6][38][61][77][78]. Если даже с Землёй не сталкивались сами кометы, пыль из их хвостов могла, попадая в атмосферу, влиять на климат[6]. Высказывались предположения, что падения небесных тел могли также усиливать тектонику литосферных плит относительно интенсивности их движения, вызванного конвекцией в мантии, и как следствие, провоцировать вулканическую активность (см. далее)[78]. Однако при том, что известно множество случаев столкновения Земли с астероидами различной величины и кометами (на 2018 год — 190 ударных кратеров[79]), с высокой достоверностью доказана только одновременность мел-палеогенового вымирания и падения метеорита[80], сформировавшего кратер Чиксулуб[61][48][81]. Кроме того, ударный кратер Сильян образовался, по недавним оценкам, за несколько миллионов лет до события Келлвассера — основного этапа девонского вымирания[82]. Также некоторые специалисты связывают события на рубеже эоцена и олигоцена с серией падений метеоритов, сформировавших Попигайскую и Чесапикскую астроблемы[61][78]. Гипотеза же о временной корреляции образования импактных (предположительно) структур Беду[83] и аномалия Земли Уилкса[84] и пермского вымирания на настоящий день считается спорной[75][85][86], как и связь триасового вымирания с кратером Рошшуар[87][88]. Не выявлено и корреляции между интенсивностью вымирания и диаметром образовавшегося максимально незадолго до него кратера (то есть энергией соударения небесного тела)[75]. Имеются кратеры, гораздо более крупные, чем знаменитый Чиксулуб, связанные таким образом с импактными событиями более мощными, но не имевшими катастрофических последствий для биосферы[76]. К тому же, если бы столкновения с небесными телами были причиной биотических кризисов, причём единственной, последние были бы весьма скоротечны, что не подтверждается данными[3]. С этим связана также трудность, заключающаяся в том, что датировка массовых вымираний возможна лишь с точностью на уровне ярусов, то есть порядка миллиона лет, тогда как разрушительные последствия импактных событий могли длиться не более 2—24 месяцев[77], поэтому проблематично делать заключения о причинно-следственной связи[89], однако и отсутствие её не доказано[61].
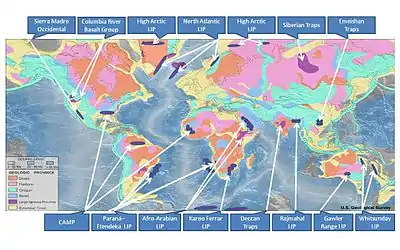
- Излияния базальта. При вулканических извержениях в атмосферу выбрасываются углекислый газ, диоксид серы, а также фтороводород, хлороводород и метан, вызывающие изменения климата, аноксию, разрушение озонового слоя, кислотные дожди и закисление океана и почвы[прим. 13][3][65]. Выбросы больших объёмов парниковых газов (таких как CO2 и CH4), возможно, происходили и при нагревании угля и других органических пород в процессе интрузивного вулканизма[90] на суше и/или посредством механизма «метангидратного ружья»[3][65].
- В последние 300 млн лет зарегистрировано 11 случаев этого типа, из которых порядка половины[прим. 14] совпало по времени с массовыми вымираниями, в том числе крупнейшими[3][91][92][93]:
Извержение Время извержения, млн лет назад Массовое вымирание Глобальное потепление δ13C Дефицит кислорода в Мировом океане Вулканизм в Волынской области 580—545 Позднеэдиакарское вымирание Извержение в районе Калкариндджи 510—503 Раннекембрийское (тойонское/ботомское) вымирание −4,0 + Центральноазиатский внутриплитовый магматизм 510—470 Позднекембрийское (дресбахское) вымирание +5,0...+6,0 (кембрийский положительный сдвиг изотопа углерода) + Вулканизм в Окчхонском рифтовом поясе 480—430 (?) Ордовикско-силурийское вымирание + (на втором этапе, наряду с оледенением на первом) +7,0, затем −7,0 Алтае-Саянский магматизм 408—393 Позднесилурийское вымирание (?) Вулканизм в Вилюйском и Днепровско-Припятском бассейнах 340—380 Девонское (франско-фаменское) (+ позднетурнейское ?) вымирание + (+6 °C) (чередующееся с промежутками похолодания) от +2,0 до +4,0 + Днепровско-Припятский вулканизм 364—367 (?) Хангенбергское событие + (наряду с похолоданием, в т. ч. эпизодами оледенения) до +6,0 + Скагерракское и Баргузин-Витимское извержение 297 ± 4 и 302—275 (?) соответственно Кризис карбоновых лесов Эмэйшанские + Панджалские траппы 265—252 (?) Гваделупское (кептенское) вымирание + (наряду с похолоданием) (?) до −6,0 локально Сибирские траппы 250 ± 3 Великое пермское вымирание + до −8,0 + Сибирские траппы 249 Поздне-нижнетриасовое (смитско-спэтское) вымирание + (+6 °C) −6,0, затем +6,0 + вулканизм в области Врангеллия 230 Карнийское плювиальное событие + (+7 °C) −5,0 + Центрально-атлантический вулканизм 200 ± 5 Триасовое вымирание + (+6 °C) −6,0 + (?) Излияние в Кару-Феррар 184 - 172 Ранне-тоарское изменение + (+7 °C) −7,0, затем +7,0 + Карибско-Колумбийский вулканизм 90 ± 3 Сеномано-туронское событие + +2 + Деканские извержения 65 ± 3 Мел-палеогеновое вымирание + (+4 °C в океане, +8 °C на суше) −2
· Периодические объединения литосферных плит в результате тектонических движений (тектоническая гипотеза), приводящие к снижению фактора географической изоляции биологических видов, резкому росту межвидовой конкуренции и вымиранию менее приспособленных к изменившимся условиям видов в соответствии с правилом конкурентного исключениях[94][95][96]. В истории биосферы Земли объединения континентов происходили регулярно, длительность таких объединений примерно совпадает с длительностью массовых вымираний (миллионы лет). При этом влияние фактора изоляции более значительно для представителей животного мира, что объясняет меньшую долю растений среди вымерших видов. Ярким подтверждением рассматриваемого механизма является великое пермское вымирание, когда практически все материки объединились в единый суперконтинент Пангею. В другие периоды объединений материков также наблюдали снижение биоразнообразия. Например, неоднократное объединение Евразии и Сев. Америки в результате образования Берингова перешейка сопровождалось позднеплейстоценовым вымиранием мегафауны. Объединение литосферных плит выступает как бы пусковым механизмом для процесса вымирания: определённую роль при этом играют также изменения климата, геохимии и др. природных условий. Отмечается также, что современный этап глобализации, приводящий как к преднамеренной интродукции, так и случайной инвазии биологических видов на чужеродные территории, ведёт к сокращению биоразнообразия в силу роста межвидовой конкуренции и сокращения ареалов обитания аборигенных видов благодаря человеку, что позволяет говорить о шестом массовом вымирании[97].

- Кроме того, ряд эпизодов совпал с периодами глобального потепления и дефицита кислорода в Мировом океане, которые также являлись факторами, влиявшими на эволюционную динамику[92]. Возможно, что эти явления, в свою очередь, были вызваны импактными событиями[61][78][98][99][100]. Между тем, отсутствует взаимосвязь с глобальными похолоданиями, не прослеживается корреляция с интенсивностью вулканических явлений, кроме того, более точное рассмотрение показывает, что основная фаза вымираний происходила до начала извержений[прим. 15][61][92]. Требует объяснения и факт, что имели место как массовые вымирания, не совпадавшие по времени с излияниями базальта, так и извержения, совпадавшие с глобальными потеплениями и аноксией, но не сопровождавшиеся вымираниями, например, сформировавшие провинцию Парана-Этендека и плато Онтонг[3][61][92][78]. Возможно, излияния, приводившие к массовым вымираниям, происходили только до разделения Пангеи[3]. Таким образом, причинно-следственная связь с высокой определённостью не установлена[92], однако она и не опровергается[61].
На оценку вероятности обеих потенциальных причин — как импактных событий, так и вулканизма — влияет то обстоятельство, что точная датировка событий, особенно палеозоя и более ранних, зачастую затруднена, так как соответствующие окаменелости и породы подверглись сильным изменениям. А даже если временно́е совпадение и имеет место, оно, строго говоря, не означает причинности[91][92][61]. Тем не менее, при взгляде на совокупную статистическую картину геологических свидетельств, всё же можно наблюдать закономерную кластеризацию падений метеоритов, проявлений вулканизма и явлений, связанных с массовыми вымираниями (дефицитом кислорода, глобальными потеплениями и т. д.). Это даёт возможность сделать вывод, что совпадения во времени двух этих факторов, которые статистически возможны раз в период порядка 100 млн лет, и могли приводить к глобальным биотическим кризисам, в то время как по отдельности они вполне могли и не быть столь фатальны[89][101].
Помимо названных возможных первичных причин, назывались более экзотические внеземные факторы:

- Космические лучи. Высказывались предположения, что они могут влиять на климат путём формирования облаков[102][103][104], хотя механизм, посредством которого это могло бы происходить, не очень ясен, и вообще это влияние, по оценкам других специалистов, весьма слабо по сравнению с другими факторами[6][105][75]. Однако при попадании космических лучей в атмосферу могут возникать потоки мюонов, оказывающие, возможно, либо непосредственное смертельное воздействие на живые организмы, либо повреждающие их ДНК[6]. Также они способствуют формированию в атмосфере ядовитых оксидов азота, вызывающих кислотные дожди и разрушение озонового слоя[102]. Достигающие земной атмосферы космические лучи могут происходить как из внегалактических источников или от вспышек сверхновых — их поток модулируется активностью Солнца посредством изменения защищающего от них Землю поля, — так и от самого Солнца[6][104][106].
- Вспышки сверхновых на близком (менее 10 пк) расстоянии — вызывают потоки космических лучей (см. выше) и гамма-излучения. Последнее как непосредственно разрушает живые клетки, так и повреждает их ДНК. Ионизирующее излучение формирует в атмосфере моноксид азота, который разрушает озоновый слой, защищающий от солнечной радиации, а также образует диоксид азота, поглощающий видимое излучение Солнца, что приводит к глобальному похолоданию[6][107][108]. Примерно такой же эффект, что и сверхновые, теоретически могут оказывать гамма-всплески. Моделирование частоты их возникновения на близком от Земли расстоянии показало, что теоретически они могли вызывать глобальные биотические кризисы 2 раза за миллиард лет или даже чаще. Особенно серьёзно оценивается вероятность возможного влияния этого фактора в событиях конца ордовикского периода[6][109].
- Взаимодействие с облаками межзвёздной пыли, которая теоретически может блокировать солнечное излучение, приводя к глобальному похолоданию. Однако высказывались и предположения, что при падении этого вещества на Солнце из-за высвобождения гравитационной энергии его светимость и, как следствие, температура на поверхности Земли, наоборот, увеличивается[6][104][107].
Долгосрочная динамика
Периодичность
Некоторые исследователи приходили к выводу, что в изменениях со временем интенсивности вымирания прослеживается периодичность, в частности, авторы концепции массовых вымираний Д. Рауп и Дж. Сепкоски оценивали длину периода в 26 млн лет[4][110].
Отмечается также изменение разнообразия видов — параметра, на который непосредственно влияют вымирания, — с периодом порядка 62 млн лет[прим. 16][5][111]. Также, по некоторым данным, в динамике биоразнообразия присутствует цикл продолжительностью порядка 140 млн лет[5][104], хотя другие специалисты считают, что это лишь артефакт метода анализа данных, к тому же за изучаемый промежуток времени с начала фанерозоя прошло бы всего 3 таких цикла, чего недостаточно для статистической значимости[111].
Данный феномен, по мнению учёных, может объясняться цикличностью процессов, предположительно связанных с биотическими кризисами. Прежде всего, предпринимались попытки её поиска в данных по импактным событиям, и некоторые специалисты, анализируя статистику ударных кратеров, действительно приходили к заключению, что падения метеоритов происходили с периодом порядка 30 млн лет, то есть близком к заявленному Раупом и Сепкоски[112][113][114]. Эти события, как и другие инициировавшие вымирания явления, могли, в свою очередь, вероятно, быть вызваны некими первичными периодическими причинами.
- Внеземные факторы:
- Колебания Солнечной системы относительно плоскости галактики при вращении вокруг её центра. Существует гипотеза, что поток высокоэнергетических космических лучей (с энергией порядка ТэВ и даже ПэВ), а также мюонов вызван взаимодействием со сверхскоплением Девы и возрастает при смещении к северу, где ослабевает экранирование галактическим магнитным полем[111]. Она основана на обнаруженном американскими физиками Робертом Роде и Ричардом Мюллером совпадении (раз в 62 ± 3 млн лет) моментов максимального удаления к северу от плоскости Галактики, максимумов интенсивности потока космических лучей и минимумов биоразнообразия[102][115]. Другие учёные считают, что более критично по своим последствиям прохождение через саму плоскость Млечного пути (по разным оценкам, раз в 26—42 млн лет[75][116][117][118]), например, помимо воздействия космических лучей[69], из-за высокой концентрации в этой области облаков межзвёздного газа и пыли[112], либо тёмной материи, что, предположительно, может вызывать возмущения в облаке Оорта, из-за чего гипотетически могло увеличиваться число столкновений Земли с кометами (падения которых, наряду с астероидами, возможно, были одной из вероятных причин массовых вымираний[78][119])[113][114], а также разогревать земное ядро, что, возможно, влекло усиление интенсивности тектонических и вулканических процессов[120]. Наконец, в основной галактической плоскости более интенсивно происходит звездообразование и выше концентрация массивных звёзд и, как следствие, вероятность вспышек сверхновых, также представляющих возможную опасность[6][117]. Между плотностью звёзд в окрестности Солнца и интенсивностью вымираний, согласно некоторым оценкам, действительно прослеживается определённая временна́я корреляция[117]. Однако строгая периодичность такого вертикального движения Солнечной системы не доказана: такие колебания носят скорее, возможно, лишь квазипериодический характер[117], период может варьироваться минимум на 20 %[75], а при прохождении спиральных рукавов даже уменьшаться вдвое[5]. Критичность последствий для биосферы Земли также достоверно не установлена[5][75]. Противники этой гипотезы приводят, например, аргумент, что за последние 15 млн лет ни одного массового вымирания не было, тогда как именно 15 млн лет назад, согласно некоторым оценкам, состоялось последнее прохождение Солнечной системы через главную плоскость[6][107]. По результатам других расчётов, это произошло в гораздо менее далёком прошлом — 1—3 млн лет назад[103][116][113], однако в любом случае, ни с каким вымиранием это явно не коррелирует[121]. Наконец, эта теория не объясняет различий в интенсивности биотических кризисов[107].
- Проход Солнечной системы через спиральные рукава Млечного пути, молекулярные облака или другие структуры с повышенной концентрацией газа, пыли и массивных звёзд, вызывающий предположительно те же эффекты, что и гипотетические последствия пересечения главной плоскости Галактики: возмущения в облаке Оорта, и, как следствие, рост число столкновений с кометами[4][5][110][119][107], возрастание потока космических лучей[103][104][118] увеличение частоты близких вспышек сверхновых[107] и гамма-всплесков[6][119]. Некоторые исследователи связывают такие крупнейшие события вымирания как пермское, мел-палеогеновое и, возможно, раннекембрийское с прохождением через рукава Щита-Центавра 245 млн лет назад, Стрельца 70—60 млн лет назад, а также, вероятно, Лебедя 490 млн лет назад соответственно[78][107]. Другие специалисты, однако, датируют пересечения рукавов Стрельца и Щита-Центавра временами 30 и 142 млн лет назад соответственно[118]. По результатам других расчётов, за последний миллиард лет произошло 7 прохождений спиральных рукавов, сопровождавшихся с задержкой в 21—35 млн лет ледниковыми периодами 20, 160, 310, 446, 592, 747 и 920 млн лет назад — в этой динамике, таким образом, был зафиксирован цикл длительностью порядка 140 млн лет[104]. Проведённые чуть позже исследования подтвердили 4 пересечения спиральных рукавов за последние 500 млн лет — 80, 156, 310 и 446 млн лет назад, также соответствовавших ледниковыми периодам[103][прим. 17]. А по самым оптимистичным оценкам, нахождение Солнечной системы в этих областях Галактики, которое могло иметь место в продолжение в общей сложности до 60 % всего времени её существования, совпадало не только со всеми пятью основными, но и с менее шестью другими менее масштабными вымираниями: позднекембрийским, эоцен-олигоценовым, силурийскими вымираниями, кризисом карбоновых лесов и др[119]. Наконец, предлагалась гипотеза комплексного 180-миллионолетнего цикла, согласно которой за последние 700 млн лет осуществлялись последовательно проходы через рукава Персея, Лебедя, Щита-Центавра и Стрельца, перемежаемые «суперхронами» стабильности 120—84, 312—264 и 485—463 млн лет назад; каждый такой цикл делился примерно на три цикла по 60 млн лет и шесть циклов по 30 млн лет — таким образом объяснялись зафиксированные в других работах периоды соответствующей длительности, кроме 140 млн лет. В рамках этой теории в так называемые зоны повышенной интенсивности вымираний попадают 20 из 22 биотических кризисов, причём они коррелируют по времени с 19 из 25 импактных событий, а также рядом эпизодов излияний базальта и зафиксированных сдвигов изотопа углерода[115].
- Однако другие учёные считают подобные заключения неубедительными, поскольку при различных попытках моделирования таких процессов присутствовал ряд неопределённых параметров и допущений[117], и даже при этом получаемые периоды оказывались далеки и от 26—30, и от 62 млн лет[6]. Особенно значительная неопределённость связана со строением рукавов: использование различных его моделей приводит к совершенно различным результатам, в том числе и полному отсутствию какой бы то ни было периодичности пересечений с рукавами и даже вообще факта их осуществления (например, это касается того же рукава Щита-Центавра)[122]. Скорее всего, прохождения Солнечной системы через спиральные рукава действительно случались не единожды за её историю — это могло происходить раз в 100—200 млн лет — однако, во-первых, динамика этого процесса весьма трудна для моделирования и, по всей видимости, более сложна, чем обычный цикл с постоянным периодом[6][117][122], а во-вторых, не доказано, что он действительно связан с усилением воздействия факторов, предположительно вызывавших биотические кризисы[6].
- Изменение расстояния до центра Галактики при вращении Солнца вокруг её центра, в котором также некоторыми учёными была выявлена периодичность в 180 млн лет[123]. Однако статистика массовых вымираний плохо укладывается в такую модель цикла, к тому же, продолжительность этого периода слишком велика, чтобы говорить о статистической значимости[6].
- Солнечные циклы — хотя и во многом определяют земной климат, имеют гораздо меньшую продолжительность, чем описываемые периоды[5][6]. Однако возможны и более долгосрочные, формирующие сложный цикл процессы, связанные с изменением магнитного поля Солнца: его смена с тороидального на полоидальное (так, что пятна формируются вдоль меридианов, а не параллелей)[прим. 18], наложившись на смену полярности земного магнитного поля, флуктуации размера Солнца и изменение его гравитационного поля теоретически могла бы повлечь усиление потока солнечной радиации, космических лучей, а также достигающих земной поверхности метеоритов. Также это повлияло бы на процессы в земной мантии и климат. В совокупности эти факторы вполне могли приводить к массовым вымираниям[106]. Вообще, Солнце как само является источником космических лучей (например, при вспышках) с энергией до 1 ГэВ, так и влияет на число их, попадающих извне: солнечный ветер формирует гелиосферу и магнитосферу Земли, защищающие от проникновения этих частиц в земную атмосферу[104][6].
- Колебания земной орбиты, которые могли оказывать влияние на климат. Однако при их моделировании не выявлено циклов длительностью, о которой идёт речь — порядка миллионов лет[5][6].
- Наличие гипотетической звезды-компаньона Солнца[124][125] теоретически способно было бы вызывать периодическое усиление потока комет, однако орбита с требуемыми (для периодов в 26 или 62 млн лет) параметрами была бы нестабильна[6][121], хотя эта нестабильность объясняла бы отсутствие строгой периодичности биотических кризисов[6]. Воздействие же более чем одной такой звезды вообще не могло бы быть периодическим[5]. В любом случае, ничего похожего на подходящих кандидатов в звёзды-компаньоны Солнца на настоящий день не обнаружено[5].
- Воздействие Девятой планеты[5][126].
- Высказывается также гипотеза наличия в Солнечной системе до сих пор не обнаруженного массивного объекта, состоящего из тёмной материи[8].
- Факторы, не связанные с космическими процессами[127]:
- Колебания уровня океана — теоретически могли бы послужить объяснением и действительно совпадают по времени со многими эпизодами вымирания[67]; в некоторых работах в их динамике был выявлен цикл продолжительностью в 36 млн лет и корреляция минимумов этой величины с усилением потока космических лучей (которое гипотетически вызывает периодические — согласно данной теории — глобальные похолодания)[69]. Однако другие специалисты такую периодичность не подтверждают; строго не доказана и причинно-следственная связь[111].
- По результатам моделирования, мантийные плюмы, отвечающие за излияния базальта, также могли образовываться периодически[128], однако в ходе других исследований цикличности в этих вулканических явлениях не выявлено[129].
- Тектоника плит — также теоретически провоцирует изменения климата и колебания уровня Мирового океана, которые, вероятно, связаны с биотическими кризисами. Эти процессы длятся сотни миллионов лет, и как раз в этом временном масштабе происходит, например, цикл Уилсона[69].
С другой стороны, есть и работы, где достоверной периодичности не обнаружено[36][117]. В частности, приводится контраргумент, что периодичность, возможно, имела место в процессах, определяющих только формирование ископаемых остатков (например, седиментации), на основании анализа которых делаются заключения о динамике биоразнообразия (в частности, массовых вымираний), а не само биоразнообразие[5][6][130]. Отмечались и различные недостатки методов анализа временных рядов применительно к палеонтологическим данным, с помощью которых делались заключения о периодичности как самих массовых вымираний[6], так и их предполагаемых причин[117][131]. При использовании альтернативных методов Фурье-анализа временно́й статистики вымираний пик на 27 миллионолетнем периоде оказывается по интенсивности одним из многих других, что делает его случайным[75]. А неопределённость самой датировки событий вымирания в 6 млн лет составляет 23 % от гипотетического цикла в 26 млн лет, что явно нивелирует достоверность подобных выводов[6].
Другие закономерности
По результатам ряда исследований, изменения со временем интенсивности массовых вымираний носят более сложный характер, чем периодические подъёмы и спады[36]. В самом деле, если биотические кризисы имели не единственную причину, а вызывались влиянием нескольких факторов в комплексе (а это, скорее всего, так[3][76]), то если даже воздействие какого-либо или каждого из них по отдельности было циклическим, в совокупности при наложении этих циклов разной продолжительности периодической картины получаться не должно[6]. Предлагалась теория о наложении на слабое периодическое действие одного либо нескольких факторов случайных сильных стрессоров[132] — она могла бы объяснить неодинаковую интенсивность вымираний, слабость и неоднозначность наблюдаемой цикличности[133].
Имеет смысл рассматривать вымирания в комплексе с обратным процессом — видообразованием и биоразнообразием — параметром, непосредственно определяемым этими явлениями в совокупности. В их динамике выявлены следующие закономерности:
- Имеет место обратная корреляция биоразнообразия с интенсивностью видообразования и прямая — с интенсивностью вымирания: чем больше разнообразие видов, тем меньше образовывалось новых и больше вероятность вымирания; видообразование и вымирание также находятся в прямой зависимости. Такие колебания возвращают глобальную экосистему к равновесию[36].
- Естественными (но не основными) механизмами регуляции являются конкуренция и хищничество. Гипотеза о самоорганизованной критичности биосферы[134] не нашла подтверждения[36], хотя в её эволюции действительно проявляются нелинейные эффекты, относящиеся, однако, к отклику системы на внешние воздействия[6].
- Эволюция в традиционном дарвиновском понимании происходит так, что более приспособленные виды более интенсивно занимают экологические ниши, освободившиеся в результате вымираний, которые тем самым играют важнейшую роль. Таким образом, концепция массовых вымираний не противоречит теории эволюции[135].
- Хотя вымирания и ограничивают рост величины разнообразия видов, на протяжении фанерозоя она всё же постепенно (а по некоторым оценкам — и экспоненциально) повышалась, тогда как интенсивность вымираний — снижалась[1][75][34]. Однако этот тренд вполне может быть артефактом метода анализа данных[36].
- На восстановление биоразнообразия после массового вымирания необходимо порядка 10 млн лет, а после такого крупного как пермское — до 40 млн лет[36]. Снижение разнообразия влечёт уменьшение вероятности нового вымирания сразу после предыдущего, на протяжении периода восстановления, что и может проявляться в виде цикла[136].
Примечания
- Примечания
- Более древние периоды не рассматриваются из-за сильных различий в эволюционной динамике, а также меньшей доступности и надёжности палеонтологических данных.
- Личков отмечает, что взгляды Кювье отнюдь не противоречили эволюционной теории, притом что из них, в отличие от теории Дарвина, по его мнению, логически следовало существование «волн жизни».
- Здесь и далее для других вымираний приводятся диапазоны соответствующих величин, согласно различным источникам.
- Жизнь в то время присутствовала только в воде.
- По другим оценкам, потери строматопороид и других губок были больше — порядка 70 % видов; тем не менее, впоследствии их разнообразие восстановилось.
- Примерное время основной фазы.
- В некоторых работах выделяется дополнительно так называемое франское событие как второй этап живетско-франского вымирания
- По некоторым оценкам, вымирание морских видов произошло даже менее чем за 8 тыс. лет.
- Считающегося[кем?], впрочем, маргинальной теорией.
- В одних работах выделяется один кризис в его конце, в других — 3 последовательных пика вымирания в начале, середине и конце яруса.
- По устоявшейся терминологии, событие именуется по редко использующимся сейчас названиям ярусов нижнего триаса.
- Важно не столько абсолютное значение, сколько градиент.
- Те же последствия, что и теоретически вызываемые импактными событиями, поэтому анализ геологических данных зачастую не даёт возможности сделать вывод о той или иной причине.
- Даются различные оценки — от 4 до 7.
- Хотя это может быть искусственным эффектом неточности датировки: излияния базальта могли длиться до миллиона лет.
- Причём само по себе это вовсе не предполагает периодичности в динамике самих массовых вымираний.
- Согласно этой гипотезе, задержка объясняется тем, что волны повышенной плотности и звездообразования, которыми являются рукава, движутся со скоростью, отличной от скорости самих звёзд и плотных облаков
- Подобные эффекты наблюдались для более молодых звёзд, тогда как их возможность для Солнца лишь моделировалась теоретически.
- Сноски
- Raup, D. M., Sepkoski, J. J., Jr. Mass extinctions in the marine fossil record : [англ.] // Science. — 1982. — Т. 215, вып. 4539 (19 March). — С. 1501—1503. — doi:10.1126/science.215.4539.1501.
- Benton M. J. When Life Nearly Died: The Greatest Mass Extinction of All Time (англ.). — Thames & Hudson, 2005. — ISBN 978-0500285732.
- David P. G. Bond, Stephen E. Grasby. On the causes of mass extinctions : [англ.] // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2017. — Т. 478 (15 July). — С. 3—29. — doi:10.1016/j.palaeo.2016.11.005.
- D. M. Raup, J. J. Sepkoski Jr. Periodicity of Extinctions in the Geologic Past : [англ.] // Proceedings of the National Academy of Sciences of the United States of America. — 1984. — Т. 81, вып. 3 (1 February). — С. 801—805. — . — doi:10.1073/pnas.81.3.801. — PMID 6583680.
- Robert A. Rohde & Richard A. Muller. Cycles in fossil diversity : [англ.] // Nature. — 2005. — Т. 434, вып. 7030 (10 March). — С. 208—210. — . — doi:10.1038/nature03339. — PMID 15758998.
- C.A.L. Bailer-Jones. The evidence for and against astronomical impacts on climate change and mass extinctions: a review : [англ.] // International Journal of Astrobiology. — 2009. — Т. 8, вып. 3. — С. 213—239. — arXiv:0905.3919. — . — doi:10.1017/S147355040999005X.
- Rampino M. R., Caldeira K. Periodic impact cratering and extinction events over the last 260 million years (англ.) // Monthly Notices of the Royal Astronomical Society : journal. — Oxford University Press, 2015. — Vol. 454, no. 4. — P. 3480—3484. — doi:10.1093/mnras/stv2088. — .
- Рэндалл, 2016, с. 10.
- Norman D. Newell. Crises in the history of life : [англ.] // Scientific American. — 1963. — Т. 208, № 2. — С. 76—92. — doi:10.1038/scientificamerican0263-76.
- Соболев Д. Н. Начала исторической биогенетики. — Харьков : Государственное издательство Украины, 1924.
- Чайковский Ю. В. Соболев и его «историческая биогенетика» // Наука о развитии жизни. Опыт теории эволюции. — Москва: Товарищество научных изданий КМК, 2006. — С. 175—181. — 712 с. — 1000 экз. — ISBN 5-87317-342-7.
- Б. Л. Личков. Геологическое время, волны жизни и изменение геологического мира // К основам современной теории Земли / отв. ред. проф. И. И. Шафрановский и проф. Б. П. Бархатов. — Ленинград: Изд-во Ленинградского Университета, 1965. — С. 87—99. — 119 с.
- Назаров В. И. Глава 8. Теллурические гипотезы сопряженной мегаэволюции и смены биот // Эволюция не по Дарвину: Смена эволюционной модели. — М.: КомКнига, 2005. — С. 252—266. — 520 с. — ISBN 5-484-00067X.
- Резанов И. А. Исчезновение динозавров // [coollib.com/b/113038/read Великие катастрофы в истории Земли]. — Москва: Наука, 1972. — 174 с. — (Общенаучные популярные издания). — 30 000 экз.
- Richard K. Bambach. Phanerozoic Biodiversity Mass Extinctions : [англ.] // Annual Review of Earth and Planetary Sciences. — 2006. — Т. 34 (June). — С. 127—155. — . — doi:10.1146/annurev.earth.33.092203.122654.
- Назаров В. И. Глава 9. «Космические» гипотезы этапности развития органического мира // Эволюция не по Дарвину: Смена эволюционной модели. — М.: КомКнига, 2005. — С. 252—266. — 520 с. — ISBN 5-484-00067X.
- Давиташвили Л. Ш. Причины вымирания организмов. — Москва: Наука, 1969. — 440 с. — 3000 экз.
- Leigh Van Valen. A new evolutionary law : [англ.] // Evolutionary Theory. — 1973. — Т. 1. — С. 1—30.
- Рэндалл, 2016, с. 230.
- The Fossil Record: A symposium with documentation / Harland W. B., Holland C. H., House M. R., Hughes N. F., Reynolds A. B., et al. eds // London (Geological Society). — 1967. — С. 1—828.
- The Fossil Record 2 / Benton, M. J. (editor). — London: Chapman & Hall, 1993. — 845 с.
- Sepkoski J. J. A Compendium of Fossil Marine Animal Genera : [англ.] / ed. D. Jablonski, M. Foote // Bulletins of American Paleontology. — 2002. — Т. 363. — С. 1—560.
- Revealing the history of life (англ.). The Paleobiology Database. Дата обращения: 18 июля 2018.
- Sepkoski, J. J., Jr. Phanerozoic Overview of Mass Extinction // Patterns and Processes in the History of Life: Report of the Dahlem Workshop on Patterns and Processes in the History of Life Berlin 1985, June 16–21 / Raup, D. M., Jablonski, D. (Eds.). — Berlin Heidelberg: Springer-Verlag, 1986. — С. 277—295. — XII, 450 с. — ISBN 978-3-642-70831-2.
- Patrick J. Brenchley, David A. T. Harper. Palaeoecology: Ecosystems, Environments and Evolution : [англ.]. — London, UK : Chapman & Hall, 1998. — С. 322. — 402 с. — ISBN 0412-434504.
- Calner M. Silurian global events – at the tipping point of climate change : [англ.] / Elewa, Ashraf M.T. (Ed.). — Berlin, Heidelberg : Springer, 2008. — Кн. Mass Extinction. — С. 21—57. — ISBN 978-3-540-75915-7.
- Hallam, 1997, с. 1.
- Hallam, 1997, с. 8.
- Hallam, 1997, с. 16.
- Steven M. Stanley. Estimates of the magnitudes of major marine mass extinctions in earth history : [англ.] // Proceedings of the National Academy of Sciences. — 2016. — Т. 113, вып. 42 (18 October). — С. E6325—E6334. — doi:10.1073/pnas.1613094113.
- Hallam, 1997, с. 17—19.
- David Jablonski. Background and Mass Extinctions: The Alternation of Macroevolutionary Regimes : [англ.] // Science. — 1986. — Т. 231, вып. 4734 (10 January). — С. 129—133. — doi:10.1126/science.231.4734.129.
- Hallam, 1997, с. 8—9.
- M.J. Benton. Diversification and extinction in the history of life : [англ.] // Science. — 1995. — Т. 268, вып. 5207 (7 April). — С. 52—58. — doi:10.1126/science.7701342.
- Hallam, 1997, с. 9.
- John Alroy. Dynamics of origination and extinction in the marine fossil record : [англ.] // Proc Natl Acad Sci U S A. — 2008. — Т. 105 (Supplement 1) (12 August). — С. 11536—11542. — . — doi:10.1073/pnas.0802597105. — PMID 18695240.
- George R. McGhee, Jr. Matthew E. Clapham, Peter M. Sheehan, David J. Bottjer, Mary L. Droser. A new ecological-severity ranking of major Phanerozoic biodiversity crises : [англ.] // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2013. — Т. 370 (15 January). — С. 260—270. — doi:10.1016/j.palaeo.2012.12.019.
- John Baez. Extinction (англ.). University of California, Riverside - Department of Mathematics (8 апреля 2006). Дата обращения: 15 марта 2018.
- J. John Sepkoski Jr. Patterns of Phanerozoic Extinction: a Perspective from Global Data Bases / Walliser O.H. (eds). — Berlin, Heidelberg : Springer, 1996. — Кн. Global Events and Event Stratigraphy in the Phanerozoic - Results of the International Interdisciplinary Cooperation in the IGCP-Project 216 “Global Biological Events in Earth History”. — ISBN 978-3-642-79636-4.
- Hallam, 1997, с. 41—49.
- Melott, A. L., B. S. Lieberman, C. M. Laird, L. D. Martin, M. V. Medvedev, B. C. Thomas, J. K. Cannizo, N. Gehrels, and C. H. Jackman. Did a gamma-ray burst initiate the late Ordovician mass extinction? : [англ.] // International Journal of Astrobiology. — 2004. — Т. 3, вып. 1. — С. 55—61. — doi:10.1017/S1473550404001910.
- Otto H. Walliser. Global Events in the Devonian and Carboniferous : [англ.]. — Berlin : Springer-Verlag, 1996. — Кн. Global Events and Event Stratigraphy in the Phanerozoic. — С. 225—250.
- George R McGhee. Extinction: Late Devonian Mass Extinction / John Wiley & Sons Ltd. — Wiley, 2012. — Кн. Encyclopedia of Life Sciences. — (Major Reference Works). — ISBN 0470066512.
- L. H. Tanner, S. G. Lucas, M. G. Chapman. Assessing the record and causes of Late Triassic extinctions : [англ.] // Earth-Science Reviews. — 2004. — Т. 65, вып. 1—2 (March). — doi:10.1016/S0012-8252(03)00082-5.
- Anthony Hallam. How catastrophic was the end‐Triassic mass extinction? : [англ.] // Lethaia. — 2007. — Т. 35 (2 January). — С. 147—157. — doi:10.1111/j.1502-3931.2002.tb00075.x.
- Massive submarine eruption bled Earth's oceans of oxygen : [англ.] // New Scientist. — 2008. — Т. 199, вып. 2665 (19 July). — С. 16.
- R. Mark Leckie, Timothy J. Bralower, Richard Cashman. Oceanic anoxic events and plankton evolution: Biotic response to tectonic forcing during the mid-Cretaceous : [англ.] // Paleoceanography. — 2002. — Т. 17, № 3 (23 August). — С. 13—1—13—29. — doi:10.1029/2001PA000623.
- С. А. Вишневский. Импактные события и вымирания организмов. Meteorite web site (2001). Дата обращения: 21 марта 2018.
- J. J. Hooker, M. E. Collinson, N. P. Sille. Eocene–Oligocene mammalian faunal turnover in the Hampshire Basin, UK: calibration to the global time scale and the major cooling event : [англ.] // Journal of the Geological Society. — 2004. — Т. 161, вып. 2 (March). — С. 161—172. — doi:10.1144/0016-764903-091.
- Главной причиной позднечетвертичного вымирания все-таки были люди, а не климат • Новости науки. «Элементы». Дата обращения: 17 августа 2020.
- Frédérik Saltré, Marta Rodríguez-Rey, Barry W. Brook, Christopher N Johnson, Chris S. M. Turney. Climate change not to blame for late Quaternary megafauna extinctions in Australia // Nature Communications. — 2016-01-29. — Т. 7. — ISSN 2041-1723. — doi:10.1038/ncomms10511.
- Gerardo Ceballos, Paul R. Ehrlich, Anthony D. Barnosky, Andrés García, Robert M. Pringle. Accelerated modern human–induced species losses: Entering the sixth mass extinction (англ.) // Science Advances. — 2015-06-01. — Vol. 1, iss. 5. — P. e1400253. — ISSN 2375-2548. — doi:10.1126/sciadv.1400253.
- Экологи подтвердили наступление эпохи шестого массового вымирания - Вести.Наука. web.archive.org (9 декабря 2019). Дата обращения: 16 марта 2020.
- Розанн Скербл. Исчезновение видов. Мир и регионы. Голос Америки (26 июля 2014). Дата обращения: 29 июля 2014.
- Bjørn Lomborg. The Skeptical Environmentalist: Measuring the Real State of the World. — Cambridge, UK : Cambridge U. Press, 2001. — ISBN 0 521 80447 7.
- Грозит ли Земле "шестое вымирание". Российская газета. Дата обращения: 16 марта 2020.
- Hallam, 1997, с. 31—32.
- Matthew R. Saltzman, Robert L. Ripperdan, M. D. Brasier, Kyger C. Lohmann, Richard A. Robison, W. T. Chang, Shanchi Peng, E. K. Ergaliev, Bruce Runnegar. A global carbon isotope excursion (SPICE) during the Late Cambrian: relation to trilobite extinctions, organic-matter burial and sea level : [англ.] // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2000. — Т. 162, вып. 3–4 (October). — С. 211—223. — doi:10.1016/S0031-0182(00)00128-0.
- Axel Munnecke, Christian Samtleben, Torsten Bickert. The Ireviken Event in the lower Silurian of Gotland, Sweden — relation to similar Palaeozoic and Proterozoic events : [англ.] // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2003. — Т. 195, вып. 1. — С. 99—124. — doi:10.1016%2FS0031-0182%2803%2900304-3.
- Stanley, S. M., Yang, X. A double mass extinction at the end of the paleozoic era : [англ.] // Science. — 1994. — Т. 266, вып. 5189 (25 November). — С. 1340—1344.
- Alvarez, W. Comparing the evidence relevant to impact and flood basalt at times of major mass extinctions : [англ.] // Astrobiology. — 2003. — Т. 3, № 1. — С. 153—161. — doi:10.1089/153110703321632480.
- Stephen E. Grasby, Benoit Beauchamp, Jochen Knies. Early Triassic productivity crises delayed recovery from world’s worst mass extinction : [англ.] // Geology. — 2016. — Т. 44, вып. 9 (1 September). — С. 779—782. — doi:10.1130/G38141.1.
- A. Ruffell, M. J. Simms, P. B. Wignall. The Carnian Humid Episode of the late Triassic: a review : [англ.] // Geological Magazine. — 2015. — Т. 153, вып. 2 (3 August). — С. 271—284. — doi:10.1017/S0016756815000424.
- Crispin T. S. Little, Michael J. Benton. Early Jurassic mass extinction: A global long-term event : [англ.] // Geology. — 1995. — Т. 23, вып. 6 (1 June). — С. 495—498. — doi:10.1130/0091-7613(1995)023<0495:EJMEAG>2.3.CO;2.
- Andrew H. Knoll, Richard K. Bambach, Jonathan L. Payne, Sara Pruss, Woodward W. Fischer. Paleophysiology and end-Permian mass extinction : [англ.] // Earth and Planetary Science Letters. — 2007. — Т. 256, вып. 3—4 (30 April). — С. 295—313. — doi:10.1016/j.epsl.2007.02.018.
- Newell, Norman D. Revolutions in the history of life : [англ.] // Geological Society of America Special Papers. — 1967. — Т. 89. — С. 63—92.
- Shanan E. Peters. Environmental determinants of extinction selectivity in the fossil record : [англ.] // Nature. — 2008. — Т. 454 (31 July). — С. 626—630. — doi:10.1038/nature07032.
- Terry Devitt. Ebb and flow of the sea drives world's big extinction events (англ.), EurekAlert! (15 June 2008).
- Slah Boulila, Jacques Laskar, Bilal U. Haq, Bruno Galbrun, Nathan Hara. Long-term cyclicities in Phanerozoic sea-level sedimentary record and their potential drivers : [англ.] // Global and Planetary Change. — 2018. — Т. 165 (June). — С. 128—136. — doi:10.1016/j.gloplacha.2018.03.004.
- A. Hallam, P. B. Wignall. Mass extinctions and sea-level changes : [англ.] // Earth-Science Reviews. — 1999. — Т. 48. — С. 217—250. — doi:10.1016/S0012-8252(99)00055-0.
- Andrew R. Cossins; K. Bowler. Temperature biology of animals : [англ.]. — New York : Chapman and Hall, 1987. — С. 30. — 339 с. — ISBN 978-94-010-7906-8.
- Peter J. Mayhew, Gareth B. Jenkins, Timothy G. Benton. A long-term association between global temperature and biodiversity, origination and extinction in the fossil record : [англ.] // Proceedings of the Royal Society B: Biological Sciences. — 2008. — Т. 275, вып. 1630 (7 January). — С. 47—53. — doi:10.1098/rspb.2007.1302.
- Geerat J. Vermeij. Ecological avalanches and the two kinds of extinction : [англ.] // Evolutionary Ecology Research. — 2004. — Т. 6, № 3 (March). — С. 315—337. — ISSN 1522–0613.
- Martin Schobben, Alan Stebbins, Abbas Ghaderi, Harald Strauss, Dieter Korn, Christoph Korte. Eutrophication, microbial-sulfate reduction and mass extinctions : [англ.] // Communicative & Integrative Biology. — 2016. — Т. 9, вып. 1 (January). — С. e1115162. — doi:10.1080/19420889.2015.1115162.
- Anatoly D. Erlykin, David A. T. Harper, Terry Sloan, Arnold W. Wolfendale. Mass extinctions over the last 500 myr: an astronomical cause? : [англ.] // Palaeontology. — 2017. — Т. 60, вып. 2 (13 February). — С. 159—167. — doi:10.1111/pala.12283.
- J. David Archibald et. al. Cretaceous Extinctions: Multiple Causes : [англ.] // Science. — 2010. — Т. 328, вып. 5981 (21 May). — С. 973. — doi:10.1126/science.328.5981.973-a.
- Owen B. Toon, Kevin Zahnle, David Morrison, Richard P. Turco, Curt Covey. Environmental perturbations caused by the impacts of asteroids and comets : [англ.] // Reviews of Geophysics. — 1997. — Т. 35, вып. 1 (1 February). — С. 41—78. — ISSN 1944-9208. — doi:10.1029/96RG03038.
- Adrian P. Jones. Impact volcanism and mass extinctions : [англ.] / Gerta Keller, Andrew C. Kerr. — Boulder, Colorado, USA : Geological Society of America, 2014. — Кн. Volcanism, Impacts, and Mass Extinctions: Causes and Effects. — С. 369—383. — 455 с. — ISBN 978-0-81372505-5.
- Earth Impact Database (англ.) (недоступная ссылка). Planetary and Space Science Centre University of New Brunswick Fredericton, New Brunswick, Canada. Дата обращения: 14 июня 2018. Архивировано 7 февраля 2015 года.
- Luis W. Alvarez, Walter Alvarez, Frank Asaro, Helen V. Michel. Extraterrestrial Cause for the Cretaceous-Tertiary Extinction : [англ.] // Science. — 1980. — Т. 208, вып. 4448 (6 June). — С. 1095—1108. — doi:10.1126/science.208.4448.1095.
- Paul R. Renne, Alan L. Deino, Frederik J. Hilgen, Klaudia F. Kuiper, Darren F. Mark, William S. Mitchell III, Leah E. Morgan, Roland Mundil, Jan Smit. Time Scales of Critical Events Around the Cretaceous-Paleogene Boundary : [англ.] // Science. — 2013. — Т. 339, вып. 6120 (8 February). — С. 684—687. — doi:10.1126/science.1230492.
- Wolf U. Reimold, Simon P. Kelley, Sarah C. Sherlock, Herbert Henkel, Christian Koeberl. Laser argon dating of melt breccias from the Siljan impact structure, Sweden: Implications for a possible relationship to Late Devonian extinction events : [англ.] // Meteoritics & Planetary Science. — 2005. — Т. 40, вып. 4 (26 January). — С. 591—607. — doi:10.1111/j.1945-5100.2005.tb00965.x.
- L. Becker, R. J. Poreda, A. R. Basu, K. O. Pope, T. M. Harrison, C. Nicholson, R. Iasky. Bedout: A Possible End-Permian Impact Crater Offshore of Northwestern Australia : [англ.] // Science. — 2004. — Т. 304, вып. 5676 (4 June). — С. 1469—1476. — doi:10.1126/science.1093925.
- Frese, R. von, Potts, L., Wells, S., Leftwich, T., Kim, H., et al. GRACE gravity evidence for an impact basin in Wilkes Land, Antarctica : [англ.] // Geochemistry, Gophysics, and Geosystems. — 2009. — Т. 10, № 2. — С. Q02014. — ISSN 1525-2027. — doi:10.1029/2008GC002149.
- Christian Koeberl, Kenneth A. Farley, Bernhard Peucker-Ehrenbrink, Mark A. Sephton. Geochemistry of the end-Permian extinction event in Austria and Italy: No evidence for an extraterrestrial component : [англ.] // Geology. — 2004. — Т. 32, вып. 12. — С. 1053—1056. — doi:10.1130/G20907.1.
- Gregory J. Retallack, Abbas Seyedolali, Evelyn S. Krull, William T. Holser, Clifford P. Ambers, Frank T. Kyte. Search for evidence of impact at the Permian-Triassic boundary in Antarctica and Australia : [англ.] // Geology. — 1998. — Т. 26, № 11 (November). — С. 979—982. — doi:10.1130/0091-7613(1998)026<0979:SFEOIA>2.3.CO;.
- Schmieder, M.; Buchner, E.; Schwarz, W. H.; Trieloff, M.; Lambert, P. A Rhaetian 40Ar/39Ar age for the Rochechouart impact structure (France) and implications for the latest Triassic sedimentary record : [англ.] // Meteoritics & Planetary Science. — 2010. — Т. 45, № 8 (5 October). — С. 225—1242. — doi:10.1111/j.1945-5100.2010.01070.x.
- Benjamin E. Cohen, Darren F. Mark, Martin R. Lee, Sarah L. Simpson. A new high‐precision 40Ar/39Ar age for the Rochechouart impact structure: At least 5 Ma older than the Triassic–Jurassic boundary : [англ.] // Meteoritics & Planetary Science. — 2017. — Т. 52, вып. 8. — С. 1600—1611. — doi:10.1111/maps.12880.
- G. Keller. Impacts, volcanism and mass extinction: random coincidence or cause and effect? : [англ.] // Australian Journal of Earth Sciences: An International Geoscience Journal of the Geological Society of Australia. — 2005. — Т. 52, вып. 4—5. — С. 725—757. — doi:10.1080/08120090500170393.
- Morgan T. Jones, Dougal A. Jerram, Henrik H. Svensen, Clayton Grove. The effects of large igneous provinces on the global carbon and sulphur cycles : [англ.] // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2016. — Т. 441 (1 January). — С. 4—21. — doi:10.1016/j.palaeo.2015.06.042.
- V. A. Kravchinsky. Paleozoic large igneous provinces of Northern Eurasia: Correlation with massextinction events : [англ.] // Global and Planetary Change. — 2012. — Т. 86—87 (April). — С. 31—36. — ISSN 0921-8181. — doi:10.1016/j.gloplacha.2012.01.007.
- P. B. Wignall. Large igneous provinces and mass extinctions : [англ.] // Earth-Science Reviews. — 2001. — Т. 53, вып. 1—2 (March). — С. 1—33. — . — doi:10.1016/S0012-8252(00)00037-4.
- David P. G. Bond, Paul B. Wignall. Mass extinctions, volcanism and impacts // Large igneous provinces and mass extinctions: an update : [англ.] / ed. by Gerta Keller, Andrew C. Kerr. — Boulder, Colorado, USA : Geological Society of America, 2014. — Кн. Volcanism, Impacts, and Mass Extinctions: Causes and Effects. — С. 29—57. — 455 с. — ISBN 978-0-8137-2505-5.
- Снакин В.В. Географическая изоляция видов как фактор глобальной динамики биоразнообразия (рус.) // Жизнь Земли : журнал. — 2016. — Т. 38, № 1. — С. 52-61. — ISSN 0514-7468.
- Снакин В.В. Массовые вымирания видов животных в истории биосферы Земли: еще одна гипотеза (рус.) // Изв. РАН. Сер. Географ. : журнал. — 2016. — № 5. — С. 82-90. — ISSN 2587-5566.
- [Snakin, V.V. Lithospheric plate tectonics and mass extinctions of biological species // IOP Conference Series: Earth and Environmental Science. – 2021. – V. 946. doi:10.1088/1755-1315/946/1/012009 ].
- [Ceballos G., Ehrlich P., Barnosky A, Garcia A., Pringle R., Palmer T. Accelerated modern human–induced species losses: Entering the sixth mass extinction // Science Advances. – 19 Jun 2015. – V. 1, No. 5 (http://advances.sciencemag.org/content/advances/1/5/e1400253.full.pdf) ].
- Таурон. Планета под ударом (октябрь 2010-4 марта 2011). Дата обращения: 22 марта 2018.
- Mark A. Richards, Walter Alvarez, Stephen Self, Leif Karlstrom, Paul R. Renne, Michael Manga, Courtney J. Sprain, Jan Smit, Loÿc Vanderkluysen, Sally A. Gibson. Triggering of the largest Deccan eruptions by the Chicxulub impact : [англ.] // GSA Bulletin. — 2015. — Т. 127, вып. 11—12 (12 November). — С. 1507—1520. — doi:10.1130/B31167.1.
- Abbott, Dallas H., and Ann E. Isley. Extraterrestrial influences on mantle plume activity : [англ.] // Earth and Planetary Science Letters. — 2002. — Т. 205, № 1—2 (30 December). — С. 53—62. — doi:10.1016/S0012-821X(02)01013-0.
- Rosalind V. White, Andrew D. Saunders. Volcanism, impact and mass extinctions: incredible or credible coincidences? : [англ.] // Lithos. — 2005. — Т. 79. — С. 299 — 316. — doi:10.1016/j.lithos.2004.09.016.
- Medvedev, M.V. & Melott, A.L. Do extragalactic cosmic rays induce cycles in fossil diversity? : [англ.] // The Astrophysical Journal. — 2007. — Т. 664 (1 August). — С. 879—889. — doi:10.1086/518757.
- D. R. Gies, J. W. Helsel. Ice Age Epochs and the Sun's Path through the Galaxy : [англ.] // The Astrophysical Journal. — 2005. — Т. 626, № 2 (June). — С. 844—848. — . — doi:10.1086/430250.
- Nir J. Shaviv. The spiral structure of the Milky Way, cosmic rays, and ice age epochs on Earth : [англ.] // New Astronomy. — 2003. — Т. 8, вып. 1 (January). — С. 39—77. — doi:10.1016/S1384-1076(02)00193-8.
- T. Sloan, A. W. Wolfendale. Cosmic rays and climate change over the past 1000 million years : [англ.] // New Astronomy. — 2013. — Т. 25 (December). — С. 45—49. — doi:10.1016/j.newast.2013.03.008.
- Robert G. V. Baker, Peter G. Flood. The Sun-Earth connect 3: lessons from the periodicities of deep time influencing sea-level change and marine extinctions in the geological record : [англ.] // Springerplus. — 2015. — Т. 4. — С. 285. — doi:10.1186/s40064-015-0942-6. — PMID 26203405.
- Erik M. Leitch, Gautam Vasisht. Mass extinctions and the sun’s encounters with spiral arms : [англ.] // New Astronomy. — 1998. — Т. 3, вып. 1 (February). — С. 51—56. — doi:10.1016/S1384-1076(97)00044-4.
- M. A. Ruderman. Possible Consequences of Nearby Supernova Explosions for Atmospheric Ozone and Terrestrial Life : [англ.] // Science. — 1974. — Т. 184, вып. 4141 (7 June). — С. 1079—1081. — doi:10.1126/science.184.4141.1079.
- Tsvi Piran, Raul Jimenez. Possible Role of Gamma Ray Bursts on Life Extinction in the Universe : [англ.] // Phys. Rev. Lett.. — 2014. — Т. 113, вып. 23 (December). — С. 231102(1)—231102(6). — doi:10.1103/PhysRevLett.113.231102.
- Tim Beardsley. Star-struck? : Impacts' role in the history of life remains contentious : [англ.] // Scientific American. — 1988. — Т. 258, № 4 (April). — С. 37—40. — doi:10.1038/scientificamerican0488-37b.
- Adrian L. Melott. Long-Term Cycles in the History of Life: Periodic Biodiversity in the Paleobiology Database : [англ.] // PLoS ONE. — 2008. — Т. 3, вып. 12. — С. e4044. — doi:10.1371/journal.pone.0004044.
- Michael R. Rampino, Richard B. Stothers. Terrestrial mass extinctions, cometary impacts and the Sun's motion perpendicular to the galactic plane : [англ.] // Nature. — 1984. — Т. 308 (19 April). — С. 709—712. — doi:10.1038/308709a0.
- W. M. Napier. Evidence for cometary bombardment episodes : [англ.] // Monthly Notices of the Royal Astronomical Society. — 2006. — Т. 366, вып. 3 (1 March). — С. 977—982. — . — doi:10.1111/j.1365-2966.2005.09851.x.
- Richard B. Stothers. The period dichotomy in terrestrial impact crater ages : [англ.] // Monthly Notices of the Royal Astronomical Society. — 2006. — Т. 365, вып. 1 (1 January). — С. 178—180. — . — doi:10.1111/j.1365-2966.2005.09720.x.
- M. Gillman, H. Erenler. The galactic cycle of extinction : [англ.] // International Journal of Astrobiology. — 2008. — Т. 7, вып. 1 (11 January). — С. 17—26. — . — doi:10.1017/S1473550408004047.
- Bahcall, J. N.; Bahcall, S. The sun's motion perpendicular to the galactic plane : [англ.] // Nature. — 1985. — Т. 316 (22 August). — С. 706—708. — ISSN 0028-0836. — . — doi:10.1038/316706a0.
- F. Feng and C. A. L. Bailer-Jones. Assessing the Influence of the Solar Orbit on Terrestrial Biodiversity : [англ.] // The Astrophysical Journal. — 2013. — Т. 768, № 2 (25 April). — С. 152. — . — doi:10.1088/0004-637X/768/2/152.
- H. Svensmark. Imprint of Galactic dynamics on Earth's climate : [англ.] // Astronomische Nachrichten. — 2006. — Т. 327, № 9 (November). — С. 866 — 870. — . — doi:10.1002/asna.200610650.
- M. D. Filipoviс, J. Horner, E. J. Crawford, N. F. H. Tothill, G. L. White. Mass Extinction And The Structure Of The Milky Way : [англ.] // Serb. Astron. J.. — 2013. — Т. 187. — С. 43—52. — doi:10.2298/SAJ130819005F.
- Does dark matter cause mass extinctions and geologic upheavals?, News & Press, Royal Astronomical Society (19 февраля 2015). Дата обращения 3 июня 2018.
- Adrian L. Melott and Richard K. Bambach. Nemesis reconsidered : [англ.] // Monthly Notices of the Royal Astronomical Society: Letters. — 2010. — Т. 407, вып. 1 (September). — С. L99—L102. — arXiv:1007.0437. — . — doi:10.1111/j.1745-3933.2010.00913.x.
- Andrew C. Overholt, Adrian L. Melott, Martin Pohl. Testing the link between terrestrial climate change and galactic spiral arm transit : [англ.] // The Astrophysical Journal Letters. — 2009. — Т. 705, № 2 (20 October). — С. L101—L103. — arXiv:0906.2777. — . — doi:10.1088/0004-637X/705/2/L101.
- G. N. Goncharov, V. V. Orlov. Global Repeating Events in the History of the Earth and the Motion of the Sun in the Galaxy : [англ.] // Astronomy Reports. — 2003. — Т. 47, вып. 11 (November). — С. 925—933. — . — doi:10.1134/1.1626195.
- Marc Davis, Piet Hut, Richard A. Muller. Extinction of species by periodic comet showers : [англ.] // Nature. — 1984. — Т. 308 (19 April). — С. 715—717. — . — doi:10.1038/308715a0.
- Richard A. Muller. Measurement of the lunar impact record for the past 3.5 b.y. and implications for the Nemesis theory : [англ.] // Geological Society of America Special Paper. — 2002. — Т. 356. — С. 659—665.
- News Staff / Source. Scientist Links Earth’s Periodic Mass Extinctions to Planet Nine (англ.), Astronomy News, Sci-News.com (31 March 2016). Дата обращения 30 июня 2018.
- Budd, G. E. The Cambrian Fossil Record and the Origin of the Phyla (англ.) // Integrative and Comparative Biology : journal. — Oxford University Press, 2003. — Vol. 43, no. 1. — P. 157—165. — doi:10.1093/icb/43.1.157. — PMID 21680420.
- Andreas Prokoph, Richard E. Ernst, Kenneth L. Buchan. Time‐Series Analysis of Large Igneous Provinces: 3500 Ma to Present : [англ.] // The Journal of Geology. — 2004. — Т. 112, № 1 (January). — С. 1—22. — doi:10.1086/379689.
- Vincent E. Courtillot, Paul R. Renne. On the ages of flood basalt events : [англ.] // Comptes Rendus Geoscience. — 2003. — Т. 335, вып. 1 (January). — С. 113—140. — doi:10.1016/S1631-0713(03)00006-3.
- Andrew B. Smith, Alistair J. McGowan. Cyclicity in the fossil record mirrors rock outcrop area : [англ.] // Biology Letters. — 2005. — Т. 1, вып. 4. — С. 443—445. — ISSN 1744-957X. — doi:10.1098/rsbl.2005.0345. — PMID 17148228.
- Jetsu, L., Pelt, J. Spurious periods in the terrestrial impact crater record : [англ.] // Astronomy and Astrophysics. — 2000. — Т. 353. — С. 409—418. — .
- Nan Crystal Arens, Ian D. West. Press-pulse: a general theory of mass extinction? : [англ.] // Paleobiology. — 2008. — Т. 34, вып. 4 (December). — С. 456—471. — doi:10.1666/07034.1.
- Georg Feulner. Limits to biodiversity cycles from a unified model of mass-extinction events : [англ.] // International Journal of Astrobiology. — 2011. — Т. 10, вып. 2 (April). — С. 123—129. — doi:10.1017/S1473550410000455.
- Ricard V. Solé, Susanna C. Manrubia, Michael Benton, Per Bak. Self-similarity of extinction statistics in the fossil record // Nature. — 1997. — Т. 388 (21 августа). — С. 764—767. — doi:10.1038/41996.
- Hallam, 1997, с. 19—20.
- Steven M. Stanley. Delayed recovery and the spacing of major extinctions : [англ.] // Paleobiology. — 1990. — Т. 16, вып. 4. — С. 401—414. — doi:10.1017/S0094837300010150.
Литература
- Лиза Рэндалл. Тёмная материя и динозавры: Удивительная взаимосвязь событий во Вселенной = Lisa Randall: "Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the Universe". — М.: Альпина Нон-фикшн, 2016. — 506 p. — ISBN 978-5-91671-646-7.
- A. Hallam, P. B. Wignall. Mass Extinctions and Their Aftermath. — New York: Oxford University Press, 1997. — 324 с. — ISBN 0-19-854916.
- Volcanism, Impacts, and Mass Extinctions: Causes and Effects : Special paper 505. Articles from International Conference, March 27-29, 2013 at London's Natural History Museum : [англ.] / editors: Gerta Keller, Andrew C. Kerr. — Boulder, Colorado, USA : Geological Society of America, 2014. — 455 с. — (GSA SPECIAL PAPERS). — ISBN 978-0-8137-2505-5.
- Global Events and Event Stratigraphy in the Phanerozoic : Results of the International Interdisciplinary Cooperation in the IGCP-Project 216 “Global Biological Events in Earth History” : [англ.] / Otto H. Walliser (Ed.). — Berlin Heidelberg : Springer-Verlag, 1996. — 333 с. — ISBN 978-3-642-79636-4.
- Anthony Hallam. Catastrophes and Lesser Calamities: The Causes of Mass Extinctions : [англ.]. — Oxford : Oxford University Press, 2004. — 240 с. — ISBN 9780192806680.
- Элизабет Колберт (Elizabeth Kolbert). Шестое вымирание. Неестественная история (The Sixth Extinction: An Unnatural History). 2014. ISBN 978-0-8050-9299-8
- Earth’s sixth mass extinction event under way, scientists warn (Guardian, 10 July 2017)
Ссылки
- Массовые вымирания — elementy.ru
- Массовые вымирания в фанерозое — Докторская диссертация Алексеева А. С., 1998 г.
- Снакин В. В. Экология, глобальные природные процессы и эволюция биосферы. Энциклопедический словарь, 2020 — https://www.rfbr.ru/rffi/ru/books/o_2101835
| В библиографических каталогах |
|---|