Остракодермы
Остракодермы, или щитковые[1][2] (лат. Ostracodermi, от др.-греч. ὄστρᾰκον — панцирь и δέρμα — кожа[3]), — группа палеозойских бесчелюстных. Получили название из-за хорошо развитого экзоскелета (панцирь из крупных костных пластин или мелких чешуй[4]).
| Парафилетическая группа животных | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
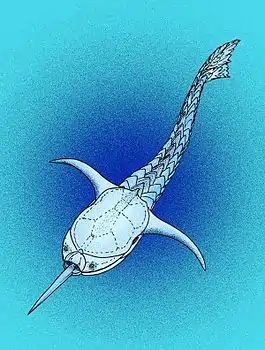 Doryaspis arctica из группы разнощитковых | |||||||||||||||||||||||||||||||||||||||||
| Название | |||||||||||||||||||||||||||||||||||||||||
| † Остракодермы | |||||||||||||||||||||||||||||||||||||||||
| Статус названия | |||||||||||||||||||||||||||||||||||||||||
| Устаревшее таксономическое | |||||||||||||||||||||||||||||||||||||||||
| Научное название | |||||||||||||||||||||||||||||||||||||||||
| Ostracodermi Cope, 1889 | |||||||||||||||||||||||||||||||||||||||||
| Родительский таксон | |||||||||||||||||||||||||||||||||||||||||
| Надкласс Бесчелюстные | |||||||||||||||||||||||||||||||||||||||||
| Представители | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология 510—370 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
По внешнему виду остракодермы были более или менее рыбообразными животными, хотя часто и без парных плавников. Их длина составляла от 2 см до 1,5 м[5], а возможно, и до 2 м[4][6]. Жили в прибрежных зонах морей и в пресных водоёмах. Известны от верхнего кембрия до верхнего девона[4], примерно 510—370 млн лет назад. По всей видимости, предки челюстноротых[7][8] (но не круглоротых, как часто считали ранее)[7][9].
Иногда рассматриваются как класс[10][11], но в большинстве современных систем не имеют таксономического ранга[7][9][12].
Описание
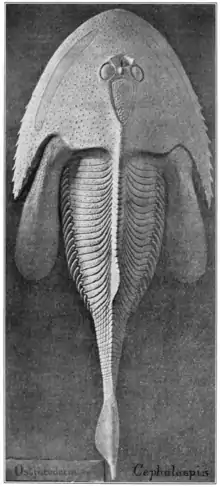
Скелет
Остракодермы имели панцирь из костных пластин или чешуйчатый покров (переднюю часть тела чаще всего защищал панцирь). Толщина его пластин у некоторых из них достигает 3 см[13]. Часто эти пластины были покрыты дентином различных видов, один из которых был практически идентичен дентину зубов человека[8][14]. У некоторых костнопанцирных верхний слой дентина панциря эмалеподобен[15].
У некоторых остракодерм (костнопанцирных и галеаспид) в голове был и внутренний скелет из кости или минерализированного хряща, и его изучение даёт много информации о морфологии органов головы (мозга, глаз, пинеального комплекса, структур внутреннего уха, обонятельной системы, гипофиза, нервов, кровеносных сосудов и жабр)[9].
У некоторых разнощитковых[16], костнощитковых[17] и бесщитковых[18] обнаружены следы дуг позвонков. У костнощитковых был плечевой пояс, слитый с черепом[19], и скелет грудных плавников в виде хрящевой пластинки (без радиалий) в каждом из них[20].
Челюстей не известно ни у кого из остракодерм[9]. Настоящих зубов у них тоже не было, но у некоторых рот был окружён специализированными зубоподобными чешуями, которые годились для соскребания корма с различных поверхностей[18], у других известны ротовые пластинки, которые могли работать как совок, а у третьих — пластинки, способные к дроблению тонких раковин[8].
Плавники
Хвостовой плавник остракодерм обычно был гетероцеркальный (разнолопастный), причём у большинства из них (у телодонтов, разнощитковых, галеаспид и анаспид), в отличие от большинства других позвоночных, гипоцеркальный: хорда заходила в нижнюю лопасть, которая при этом обычно была больше верхней. Этот плавник и был их движителем. У многих видов он был единственным плавником[4][21][8].
Спинной плавник (иногда два) был у большинства костнопанцирных и телодонтов. Анальный плавник встречался в этих же группах и у беспанцирных[6].
Парные грудные плавники были у большинства костнопанцирных, у питуриаспид, у некоторых телодонтов и беспанцирных. Настоящих брюшных плавников у остракодерм (как и у всех бесчелюстных) не было[4][12], но некоторые беспанцирные[9] и телодонты[6] имели парные плавниковые складки на брюшной стороне.
Органы чувств
Глаза были у большинства остракодерм. В головном щите многих из них есть отверстие и для теменного глаза, расположенное примерно между парными глазницами. Оно, хотя и никогда не превышало их по размеру, часто было довольно крупным[22]. У арандаспид такое отверстие было парным[21][16], но по другим интерпретациям оно у них не имело отношения к теменному глазу[23].
У многих остракодерм известна сейсмосенсорная система (органы боковой линии). У разнощитковых она компенсировала их слабое (а иногда и отсутствующее) зрение[8]. На головном щите костнопанцирных видны структуры, которые интерпретируются как рецепторы давления[4], электрические органы[24] или электрорецепторы[25]. Это два боковых и одно спинное поле мелких костных пластинок, к которым изнутри подходят ветвящиеся каналы, соединяющиеся с внутренним ухом[26][27][4].
Обонятельная система большинства[28] непарноноздрёвых, одной из двух основных групп остракодерм, была (как и у круглоротых) тесно связана с гипофизом. Ноздря, соединенная с гипофизарным проходом, образовывала в головном панцире непарноноздрёвых назогипофизарное отверстие[8][4].
Образ жизни
Остракодермы населяли прибрежные зоны морей и пресные водоёмы. Для видов с уплощённой формой тела и тяжёлым костным панцирем предполагается придонный образ жизни. Некоторые из них — костнощитковые, подобные цефаласпису, — имели хорошо развитую мускулатуру задней части тела и парные плавники, что указывает на относительно большую подвижность[8]. У беспанцирных было веретенообразное тело и лёгкий гибкий чешуйчатый покров; возможно, они жили в толще воды[4]. Это предполагается и для телодонтов. Хотя приспособления остракодерм к плаванию были, по всей видимости, сравнительно малоэффективны, гидродинамическое моделирование для некоторых из них показывает, что выпуклость брюшной стороны могла создавать подъёмную силу при движении (которое обеспечивалось хвостовым плавником)[8]. По крайней мере большинство остракодерм не были способны к продолжительному активному плаванию, но могли планировать в толще воды.
Питались остракодермы, вероятно, только мелкой пищей[29] путём всасывания, подхватывания[30] или отцеживания через жаберные щели[2]. Возможно, некоторые разнощитковые могли нападать на червей[2], соскребать обрастателей с разных твёрдых поверхностей[18] и даже дробить раковины[8]. Некоторые имели рострум, возможно, позволявший выкапывать спрятавшихся в песке мелких животных.
Ни у каких групп остракодерм не заметно увеличения со временем количества сильно специализированных форм. Среди последних видов (средне- и верхнедевонских) таких было особенно мало. Это указывает на то, что каждая из их групп в течение всего своего существования сохраняла примерно постоянный образ жизни и экологическую нишу[29].
Время жизни
Самые ранние известные остракодермы относятся к верхнему кембрию и известны только по мелким костным фрагментам. Кембрийскими представителями остракодерм были Anatolepis, известный из Северной Америки, Гренландии и Шпицбергена начиная с франконского века (около 495 млн лет назад)[31][4][16] и, возможно, безымянный организм[32] из отложений возрастом 510 млн лет[33] в Австралии.
Хорошо сохранившиеся окаменелости остракодерм относятся ко времени от раннего ордовика до позднего девона (480—370 млн лет назад)[34]. Расцвет этой группы был в позднем силуре — раннем девоне[30] (около 420 млн лет назад)[35]. Для девонских отложений они служат руководящими ископаемыми[8][30].
В среднем девоне (эйфельский век, около 390 млн лет назад) начался упадок самых многочисленных ранее групп остракодерм: разнощитковых, костнощитковых, галеаспид и телодонтов. Беспанцирные исчезли ещё в начале девона, около 420 млн лет назад (хотя иногда к ним относят и ряд родов, доживших до верхнего девона). В позднем девоне (360—380 млн лет назад) вымерли последние остракодермы[29].
Причины вымирания
Упадок и вымирание остракодерм обычно объясняют вытеснением их челюстноротыми[4][9]. С другой стороны, есть местонахождения, где одновременно была богатая фауна тех и других. Хищничество со стороны челюстноротых вряд ли было очень интенсивным: находок, которые его подтверждают, известно (на 2004 год) только две. Это мелкий экземпляр костнопанцирного в желудке акантоды из нижнего девона и след укуса (вероятно, лопастепёрой рыбы) на панцире разнощиткового из верхнего девона[29].
Возможно, какую-то роль в вымирании остракодерм сыграла трансгрессия моря в середине девона, которая, по некоторым данным[29], привела к сокращению площади подходящих для них местообитаний. На телодонтов, которые, видимо, лучше других были приспособлены к жизни в открытом море, это подействовало меньше[29]. Ещё одна гипотеза основывается на том, что упадок остракодерм был одновременным с распространением на суше сосудистых растений. Оно привело к формированию закреплённых почв и, как следствие, к уменьшению количества смываемых в моря частиц органики, которыми эти животные, вероятно, в основном и питались[29].
Распространение
Остракодермы жили в прибрежных зонах морей и пресных водоёмах, и широкие моря были для них преградой. Поэтому большинство их видов (и многие более крупные таксоны) были эндемиками небольших областей[29][8].
До конца ордовика — середины силура (440—430 млн лет назад[35]) большинство остракодерм (в частности, астраспиды, разнощитковые и телодонты) встречались только в водах Лаврентии. Вероятно, там жил и последний общий предок всех остракодерм[36]. Но арандаспиды, тоже одна из древнейших групп (известны с нижнего ордовика[16]), жили только в водах Гондваны (их окаменелости находят в Австралии и Южной Америке)[36].
В кембрии и ордовике не было космополитичных групп остракодерм: в разных регионах планеты их фауна сильно отличалась. Ареал большинства групп оставался нешироким до среднего силура, когда сближение Лаврентии с другими континентами (в первую очередь с Балтикой) сделало возможным его увеличение[36][27]. Так, телодонты к концу ордовика распространились из вод Лаврентии в воды Балтики и Гондваны, а к середине силура — и Ангариды[36]. Начиная с раннего девона, фауна телодонтов почти по всему миру стала одинаковой[27]. В Гондване они прожили на несколько десятков миллионов лет дольше, чем в Еврамерике[37], но большинство остальных остракодерм Гондваны исчезло в верхнем ордовике (возможно, из-за оледенения)[36].
Появившиеся позже непарноноздрёвые остракодермы распространились не менее широко (чего нельзя сказать про отдельные их виды и роды). Беспанцирные известны из Еврамерики, костнопанцирные, кроме того, из Сибири и Центральной Азии. Но общих родов между европейской и североамериканской фауной костнопанцирных даже в девоне почти не было[29]. Ареал остальных групп непарноноздрёвых был не таким большим: окаменелости галеаспид находят только на юге Китая и на севере Вьетнама, а питуриаспид — только в одном местонахождении в Австралии[27][34][17].
Классификация
Раньше остракодерм (как и других бесчелюстных) иногда включали в состав рыб, а иногда и объединяли с плакодермами под названием «панцирные рыбы»[3] (сейчас под ним понимают только плакодерм)[38].
Первые исследователи считали остракодерм костными рыбами[34][8]. К бесчелюстным их отнёс в 1889 году Эдвард Коп, который и ввёл термин «остракодермы». Он включил эту группу вместе с миногообразными и миксинообразными в состав бесчелюстных (остракодермы у Копа имели ранг подкласса, а бесчелюстные — класса). Коп выделял два отряда остракодерм: Arrhina («безноздрёвые») и Diplorhina («парноноздрёвые»)[39]. Отряд Arrhina включал костнощитковых и разнощитковых[40][7], а отряд Diplorhina — животное Mycterops, впоследствии оказавшееся ракоскорпионом[41].
В 1924 году Йохан Киер поделил бесчелюстных на парноноздрёвых (Diplorhina), включавших разнощитковых и телодонтов, и непарноноздрёвых (Monorhina), включавших костнопанцирных, беспанцирных, миногообразных и миксинообразных. Эрик Стеншё в своих работах 1927 и 1932 года предложил другую систему: в группу Cephalaspidomorphi попали костнопанцирные, беспанцирные и миногообразные, а в группу Pteraspidomorphi — разнощитковые и миксинообразные. Бо́льшую популярность среди палеонтологов получила система Киера[40]. Но сейчас Monorhina обычно синонимизируют с Cephalaspidomorphi, а Diplorhina — с Pteraspidomorphi[4][30][16] и придают этим группам (если считают их таксонами) ранг классов[42][30][12][21] или подклассов[4]. При этом миногообразных[42][43], а иногда и миксинообразных[44][13], включают в состав Monorhina, а иногда не относят ни к тем, ни к другим[30]. Таким образом, в этих системах остракодермы не образуют таксон. Исследователи, которые придерживаются кладистического подхода, тоже не придают им таксономического ранга из-за их парафилетичности[34][7][9]. Но некоторые другие учёные рассматривают их как класс[10][11].
Обычно к остракодермам относят следующие группы[4][7][9]:
- Парноноздрёвые (Diplorhina, Pteraspidomorphi). Из двух основных групп остракодерм эта возникла первой. Эти животные имели парные носовые мешки и отверстия ноздрей (по крайней мере, в тех случаях, когда это известно)[4]. Их обонятельная система не была связана с гипофизом. Элементы экзоскелета состоят из дентина и аспидина; клеточной костной ткани и перихондральных окостенений не было[45].

- Арандаспиды[6][21] (Arandaspida). Иногда их включают в состав разнощитковых[4][46]. 2—4 монотипических рода из ордовика Южной Америки и Австралии[23]. В длину достигали около 20 см[27], иногда до 35 см[47][6]. Передняя часть тела покрыта двумя большими, но тонкими костными пластинами: спинной и брюшной (с рядом мелких пластинок между ними), задняя — чешуёй[6][23]. Внутренних окостенений почти не было. Имели только один плавник — хвостовой. Жаберных отверстий много (более 10 пар). Примечательный признак — пара отверстий в передней части спинного щита, которые обычно интерпретируются как парные пинеальное[6][21][26] или пинеальное с парапинеальным[48][16] отверстия (очень редкая, хотя и не уникальная[49], ситуация среди позвоночных), а иногда — как выходы эндолимфатических протоков[23].

- Астраспиды[6][21] (Astraspida). Иногда их включают в состав разнощитковых[4][46]. 1—3 рода из ордовика и, возможно, силура Северной Америки и Сибири. Размер этих животных достигал 15 см[6]. Голова покрыта массивным панцирем, который образовывался из сраставшихся мелких пластинок, задняя часть — чешуёй. Вероятно, был и внутренний скелет (хрящевой). Как и у арандаспид, единственным плавником был хвостовой[50]. Жаберных отверстий 8—10 (до 13)[6] пар[16][21][51].
- Eriptychiida. Иногда их включают в состав астраспид, иногда — разнощитковых. Есть предположение о близости их к челюстноротым[21][16]. Один род с двумя видами из ордовика Северной Америки. Известны только по мелким обломкам. Строением головного панциря напоминают астраспид, микроструктурой — разнощитковых, но отличаются от них хорошо минерализированным внутренним скелетом. Внешне были, скорее всего, похожи на астраспид[52].

- Разнощитковые[6][21] или гетеростраки[4][53] (Heterostraci). Большая и разнообразная группа. Известно около 300 видов из силура и девона Европы, Сибири и Северной Америки. Самые маленькие имели размер 2 см, а самые большие — 1,5 м[5], а возможно, и 2 м[4][6]. Костное покрытие головы состояло из различного числа пластин — от одной до многих. Эндоскелета почти[8] не было[26]. Настоящих плавников, кроме хвостового, не имели, но у некоторых по бокам тела были парные костные пластинки, которые работали как рули. Глаза очень маленькие, по бокам головы. Жаберных отверстий только одна пара[21][54].

- Телодонты[4][21] (Thelodonti), ранее[2] известны как целолепиды[4] (Coelolepida)[9]. Иногда их выделяют из состава Pteraspidomorphi[21]. Разнообразная и, возможно, не монофилетическая группа. Известно 22 рода[6]. Жили с верхнего ордовика по верхний девон и были распространены очень широко: окаменелости встречаются и в Евразии, и в Северной Америке, и на континентах, входивших в состав Гондваны. Размер — 10—20 см[55], иногда от 2 и до 40 см[56]. Были целиком покрыты мелкими чешуйками. Внутренних окостенений не было[4]. Спинная и брюшная сторона у многих почти не отличались. Часто были, помимо хвостового, спинной и анальный плавник, а также парные грудные плавники[55]. Жаберных отверстий (когда их число известно) 7—8 пар[56][4][21][37].
- Непарноноздрёвые (Monorhina, Cephalaspidomorphi). Появились позже парноноздрёвых (в силуре). У этих животных (как и у круглоротых) ноздря была одна (но у галеаспид[28][8][4] и питуриаспид[57][6] в неё открывались два носовых мешка). В экзоскелете, кроме дентина и аспидина, у некоторых есть костная ткань с остеоцитами, а в эндоскелете — перихондральные окостенения[44].
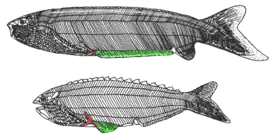

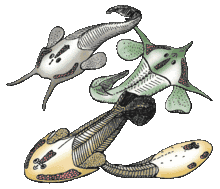
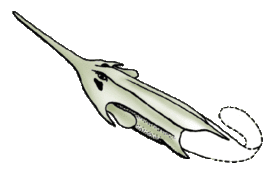
- Беспанцирные[58], бесщитковые[6][21] или анаспиды[4][59] (Anaspida). Филогенетическое положение точно не установлено[60], иногда их выделяют из состава Cephalaspidomorphi[21]. Известны около 20 родов (в некоторых системах больше)[6] из силура и девона Европы и Северной Америки. Длина — 10—15 см. Окостенения слабо развиты (что сильно затрудняет изучение этих животных), вместо массивного головного щита были небольшие пластинки и чешуя. Кроме хвостового плавника, у некоторых был маленький анальный, а кроме того, парные плавники — грудные или тянувшиеся на большую длину вдоль нижней стороны тела[60][61][21]. Жаберных отверстий 6—15 пар. Форма тела веретеновидная, и из-за этого они напоминали рыб сильнее всех остальных остракодерм[4].
- Галеаспиды[4][6] (Galeaspida). Их тоже не всегда включают в состав Cephalaspidomorphi[8]. Разнообразная группа из силура и девона Китая и Вьетнама. Около 30 родов[6] и 70 видов[62]. Имели цельный массивный головной щит, иногда с длинными выростами вперёд или вбок, длина которого составляла от 2 до 30 см[6]. Были окостенения и внутри головы. Никаких плавников, кроме хвостового, не было. Жаберных отверстий от 6 до 45 пар (рекорд среди позвоночных). Уникальный признак — большое отверстие на голове спереди сверху, которое соединяло носовые полости, глотку и внешнюю среду[28][21][62][63][4].
- Костнощитковые, костнопанцирные[6][21], остеостраки[64] или цефаласпиды[4] (Osteostraci). Изучены лучше всех остальных остракодерм. Большая группа (около 40 родов[6] и 200 видов) из силура и девона Евразии и Северной Америки[65]. В длину достигали от 2 см[64] до 60 см[21] (по другим данным, и до 1 м[65]). Имели массивный череп с внутренними окостенениями, покрытый пластинами дермальной кости (головным щитом), обычно с выростами по бокам. Кроме хвостового плавника, некоторые имели один или два спинных, и, возможно, анальный; обычно были и мускулистые парные грудные плавники. Вероятно, относительно хорошо плавали[6]. Жаберных отверстий около 10 пар[64][21][4]. Самый заметный уникальный признак — три поля на головном щите, которые интерпретируются как следы электрических органов[24], электрорецепторов[25] или рецепторов давления[4].
- Питуриаспиды[6] (Pituriaspida). Маленькая и плохо изученная группа из девона Австралии, известно два монотипических рода. Видимо, близки к галеаспидам и остеостракам, и условно включаются в состав Cephalaspidomorphi[21][17][66]. Имели тяжёлый головной щит[27] длиной до 9 см[6] с длинным выростом спереди и более короткими по бокам. По-видимому, были хорошо развитые парные грудные плавники[57]. Уникальный признак — отверстие с неизвестной функцией на нижней стороне головы. Про заднюю часть тела ничего не известно[17].
Этим группам придают ранг отрядов[2][27][21], надотрядов[66], подклассов[21][67][6], классов[21][28][66] или надклассов[21].
Кроме того, иногда остракодермами называют даже две группы бесчелюстных, не имевших панциря: Euphaneropidae (возможно, относятся к анаспидам[9]) и конодонтоносителей[7][27].
Эволюция
Считается установленным, что остракодермы произошли от близкого к конодонтоносителям предка и дали начало челюстноротым[9]. Но в эволюционных связях разных групп остракодерм до сих пор остаётся много неясного[34].
Возможно, челюстноротые произошли от остракодерм, близких к костнопанцирным[34][27][9][7]. Есть и предположение об их происхождении от телодонтов[68] или близких к ним форм[8]. Миногообразные и миксинообразные, которых раньше часто считали потомками остракодерм, скорее всего, таковыми не являются[7][9].
Изучение остракодерм внесло существенный вклад в современные представления об эволюции позвоночных. По всей видимости, кожные окостенения возникли раньше внутренних[7], а отсутствие костных клеток, как и парность ноздрей и многочисленность жаберных отверстий, для позвоночных первичны[4]. Строение костнопанцирных — по некоторым представлениям, самых близких родственников челюстноротых, — показывает, что ещё до возникновения челюстей у позвоночных появились клеточная костная ткань, перихондральные кости, склеротические кольца (окостенения в глазах), парные грудные плавники, плечевой пояс и эпицеркальный хвостовой плавник[7] (хотя есть мнение, что по крайней мере некоторые из этих структур у костнопанцирных и челюстноротых возникли независимо[4]). Но о деталях происхождения челюстей[9] и парных плавников, а также внутреннего уха, окаменелости остракодерм говорят очень мало или ничего[26].
Представления о родственных отношениях разных групп остракодерм у разных авторов сильно отличаются. Особенно неопределённо положение беспанцирных, потому что отсутствие у них хорошо развитых окостенений сильно затрудняет изучение их строения[9]. По одной версии, их предки отделились от «ствола» филогенетического дерева остракодерм первыми[9], по другой — после предков астраспид, арандаспид и разнощитковых[27][69], по третьей — после телодонтов[34][60], по четвёртой — беспанцирные вместе с костнопанцирными и круглоротыми образуют ветвь, сестринскую по отношению к группе, включающей остальных остракодерм и челюстноротых[70].
Кладограмма низших позвоночных по Ф. Жанвье, 2008[9] (группы, относящиеся к остракодермам, выделены жирным):
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма низших позвоночных по Л. И. Новицкой, 2004[70] (группы, относящиеся к остракодермам, выделены жирным):
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
- Остракодермы — статья из Большой советской энциклопедии
- Ромер А., Парсонс Т. Анатомия позвоночных. — М.: Мир, 1992. — Т. 1. — С. 57—61. — 358 с. — ISBN 5-03-000291-X.
- Остракодермы // Геологический словарь: в 2-х томах / К. Н. Паффенгольц и др. — 2-е изд., исправл. — М.: Недра, 1978. — Т. 2. — С. 51.
- Кэрролл Р. Палеонтология и эволюция позвоночных. — М.: Мир, 1992. — Т. 1. — С. 36—55. — 280 с. — ISBN 5-03-001819-0.
- Janvier, Philippe. Heterostraci. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Иванов А. О., Черепанов Г. О. Ископаемые низшие позвоночные: учебное пособие. — 2. — СПб.: Издательство Санкт-Петербургского университета, 2007. — С. 54—87. — 228 с. — ISBN 978-5-288-04342-0.
- McKenzie D. J., Farrell A. P., Brauner C. J. Volume 26. Primitive Fishes // Fish Physiology. — Academic Press, 2007. — P. 31—33. — 576 p. — ISBN 978-0-12-373671-0.
- Новицкая Л. И. Бесчелюстные (Agnatha) как древнейшие позвоночные: основные результаты исследований // Эволюция биосферы и биоразнообразия: к 70-летию А.Ю. Розанова / С. В. Рожнов. — М.: Товарищество научных знаний КМК, 2006. — С. 193—207. — 600 с. — ISBN 5-87317-299-4.
- Janvier, Philippe. Early Jawless Vertebrates and Cyclostome Origins (англ.) // Zoological Science. — Zoological Society of Japan, 2008. — Vol. 25, no. 10. — P. 1045—1056. — doi:10.2108/zsj.25.1045. — PMID 19267641.
- Kotpal R. L. Modern Text Book of Zoology: Vertebrates. — New Delhi: Rastogi Publications, 2001. — P. 111—114. — 864 p. — ISBN 81-7133-891-7.
- Shukla J. P. Fish and Fisheries (неопр.). — 2. — New Delhi: Rastogi Publications, 2010. — С. 640. — ISBN 81-7133-800-3.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Agnatha. Бесчелюстные, с. 10.
- Михайлова И. А., Бондаренко О. Б. 1 // Палеонтология. — М.: Издательство МГУ, 1997. — С. 360—364. — 448 с. — ISBN 5-211-03841-X.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Подкласс Heterostraci. Гетеростраки, с. 100.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Подкласс Osteostraci. Остеостраки, с. 213.
- Pteraspidomorphi. Anatolepis, Arandaspids & Astraspids (англ.). Palaeos: Life Through Deep Time. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Cephalaspidomorphi: Osteostraci (англ.). Palaeos: Life Through Deep Time. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Kardong K. V. 7: Skeletal System: The Skull // Vertebrates: Comparative Anatomy, Function, Evolution. — 4. — McGraw-Hill College, 2005. — P. 239—250. — 800 p. — ISBN 0070607508.
- Zhu M., Yu X., Choo B., Qu Q., Jia L., Zhao W., Lu J. Fossil Fishes from China Provide First Evidence of Dermal Pelvic Girdles in Osteichthyans (англ.) // PLOS One : journal. — Public Library of Science, 2012. — Vol. 7, no. 4. — P. e35103. — doi:10.1371/journal.pone.0035103. — PMID 22509388.
- Janvier P., Arsenault M., Desbiens S. Calcified cartilage in the paired fins of the osteostracan Escuminaspis laticeps (Traquair 1880), from the Late Devonian of Miguasha (Quebec, Canada), with a consideration of the early evolution of the pectoral fin endoskeleton in vertebrates (англ.) // Journal of Vertebrate Paleontology : journal. — Society of Vertebrate Paleontology, 2004. — Vol. 24, no. 4. — P. 773—779. — doi:10.1671/0272-4634(2004)024[0773:CCITPF]2.0.CO;2.
- Нельсон Д. С. Рыбы мировой фауны / Пер. 4-го перераб. англ. изд. Н. Г. Богуцкой, науч. ред-ры А. М. Насека, А. С. Герд. — М.: Книжный дом «ЛИБРОКОМ», 2009. — С. 73–80. — ISBN 978-5-397-00675-0.
- Ромер А., Парсонс Т. Анатомия позвоночных. — М.: Мир, 1992. — Т. 2. — С. 226—227. — 406 с. — ISBN 5-03-000290-1.
- Janvier, Philippe. Arandaspida. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Лабас Ю. А., Глухова Л. Н. Механизм расширения и смены биологических функций в филогенезе на примере специализированных электрогенераторных тканей рыб // Журнал общей биологии : журнал. — 1999. — Т. 60, № 5. — С. 510—526.
- Шабанов Д. А. III. Vertebrata-01. Бесчелюстные. Лекторий-библиотека Batrachos.com. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Janvier P. Skull of Jawless Fishes // The Skull (неопр.) / J. Hanken, B. K. Hall. — University of Chicago Press, 1993. — Т. 2: Patterns of Structural and Systematic Diversity. — С. 152—188. — ISBN 0-226-31568-1.
- Benton, Michael J. Vertebrate Palaeontology (неопр.). — 3. — Blackwell Publishing, 2005. — ISBN 978-0632056378. Архивированная копия (недоступная ссылка). Дата обращения: 21 ноября 2012. Архивировано 19 октября 2008 года.
- Gai Zh., Donoghue P. C. J., Zhu M., Janvier, P., Stampanoni M. Fossil jawless fish from China foreshadows early jawed vertebrate anatomy (англ.) // Nature : journal. — 2011. — Vol. 476, no. 7360. — P. 324—327. — doi:10.1038/nature10276. — PMID 21850106.
- Janvier P., Newman M. J. On Cephalaspis magnifica Traquair, 1893, from the Middle Devonian of Scotland, and the relationships of the last osteostracans (англ.) // Transactions of the Royal Society of Edinburgh: Earth Sciences : journal. — 2004. — Vol. 95, no. 3—4. — P. 511—525. — doi:10.1017/S0263593300001188.
- Бесчелюстные // Биологический энциклопедический словарь / М. С. Гиляров и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986. — С. 59. — 831 с.
- Smith M. P., Sansom I. J. The affinity of Anatolepis Bockelie & Fortey (неопр.) // Geobios. — 1995. — Т. 28. — С. 61—63. — doi:10.1016/S0016-6995(95)80088-3.
- Young, G. C., Karatajute-Talimaa V. N., Smith M. M. A possible Late Cambrian vertebrate from Australia (англ.) // Nature. — 1996. — Vol. 383, no. 6603. — P. 810—812. — doi:10.1038/383810a0.
- Gee, Henry. Fishy fragments tip the scales (англ.) (недоступная ссылка). University of California: Richard Cowen pages. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Janvier, Philippe. Vertebrata. Animals with backbones. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 11 декабря 2012 года.
- International Chronostratigraphic Chart (англ.). International Commission on Stratigraphy (январь 2015). Дата обращения: 11 апреля 2015. Архивировано 11 апреля 2015 года.
- Smith M. P., Donoghue P. C. J., Sansom I. J. The spatial and temporal diversification of Early Palaeozoic vertebrates (англ.) // Geological Society, London, Special Publications : journal. — 2002. — Vol. 194, no. 1. — P. 69—83. — doi:10.1144/GSL.SP.2002.194.01.06.
- Janvier, Philippe. Thelodonti. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Панцирные рыбы // Биологический энциклопедический словарь / М. С. Гиляров и др. — 2-е изд., исправл. — Москва: Сов. Энциклопедия, 1986. — С. 446. — 831 с.
- Cope E. D. Synopsis of the Families of Vertebrata (англ.) // The American Naturalist. — University of Chicago Press, 1889. — Vol. 23, no. 274. — P. 849—877. — doi:10.1086/275018.
- Tarlo, Lambert Beverley. The classification and evolution of the Heterostraci (англ.) // Acta Palaeontologica Polonica : journal. — Polish Academy of Sciences, 1962. — Vol. 7, no. 1—2. — P. 249—290.
- Jordan, David Starr. The genera of fishes. Part III... A contribution to the stability of scientific nomenclature (англ.). — Stanford University, 1917. — P. 434. — doi:10.5962/bhl.title.14748.
- Система рыб мировой фауны. Зоологический институт Российской академии наук. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Matthews, Iain. Agnatha (англ.) (недоступная ссылка). University of St Andrews, School of Biology. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Класс Monorhina. Непарноноздрёвые, с. 209.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, класс Diplorhina. Парноноздрёвые, с. 11.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Подкласс Heterostraci. Гетеростраки, с. 90.
- Gagnier P.-Y, Blieck A. On Sacabambaspis janvieri and the vertebrate diversity in Ordovician seas // Fossil fishes as living animals. Proc. 2nd Intern. Coll. Middle Palaeoz. Fishes. Academia, vol. 1 / E. Mark-Kurik. — Tallinn: Acad. Sci. Estonia, 1989. — P. 9—20.
- Blieck A., Elliott D. K., Gagnier P. Y. Some questions concerning the phylogenetic relationships of heterostracans, Ordovician to Devonian jawless vertebrates // Early vertebrates and related problems of evolutionary biology (Intern. Symp. Beijing, 1987) / Chang Mee-mann, Liu Yu-hai, Zhang Guo-rui. — Beijing: Science Press, 1991. — P. 12—13.
- Eakin R. M. The Third Eye. — Berkeley: University of California Press, 1973. — P. 16—17. — 168 p. — ISBN 0-520-02413-3.
- Janvier, Philippe. Pteraspidomorphi. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Janvier, Philippe. Astraspida. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Janvier, Philippe. Eriptychiida. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Подкласс Heterostraci. Гетеростраки, с. 69.
- The Heterostraci (англ.). Palaeos: Life Through Deep Time. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Thelodonti. Nikoliviids and Furcacaudiforms (англ.). Palaeos: Life Through Deep Time. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Подкласс Thelodonti. Телодонты, с. 20.
- Janvier, Philippe. Pituriaspida. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Анаспиды // Биологический энциклопедический словарь / М. С. Гиляров и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986. — С. 26. — 831 с.
- Анаспиды — статья из Большой советской энциклопедии
- Janvier, Philippe. Anaspida. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 11 декабря 2012 года.
- The Anaspida - unarmoured "ostracoderms" (англ.) (недоступная ссылка). Palaeos: Life Through Deep Time. Дата обращения: 21 ноября 2012. Архивировано 11 декабря 2012 года.
- Janvier, Philippe. Galeaspida. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Galeaspida (англ.). Palaeos: Life Through Deep Time. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Подкласс Osteostraci. Остеостраки, с. 210.
- Janvier, Philippe. Osteostraci. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Fish (англ.). Fishindex on users.atw.hu. Дата обращения: 21 ноября 2012. Архивировано 11 декабря 2012 года.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Содержание, с. 423.
- Cephalaspidomorphi Overview (англ.). Palaeos: Life Through Deep Time. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Mikko Haaramo. Vertebrata - animals with backbones. Mikko's Phylogeny Archive (7 декабря 2011). Дата обращения: 21 ноября 2012. Архивировано 11 декабря 2012 года.
- Бесчелюстные и древние рыбы / Л. И. Новицкая, 2004, Подкласс Heterostraci. Гетеростраки, с. 96.
Литература
- Бесчелюстные и древние рыбы / Л. И. Новицкая. — М.: ГЕОС, 2004. — С. 10—268. — 436 с. — ISBN 5-89118-368-7.
- Новицкая Л. И. Бесчелюстные (Agnatha) как древнейшие позвоночные: основные результаты исследований // Эволюция биосферы и биоразнообразия: к 70-летию А.Ю. Розанова / С. В. Рожнов. — М.: Товарищество научных знаний КМК, 2006. — 600 с. — ISBN 5-87317-299-4.
- Иванов А. О., Черепанов Г. О. Ископаемые низшие позвоночные: учебное пособие. — 2. — СПб.: Издательство Санкт-Петербургского университета, 2007. — С. 54—87. — 228 с. — ISBN 978-5-288-04342-0.
- Кэрролл Р. Палеонтология и эволюция позвоночных. — М.: Мир, 1992. — Т. 1. — С. 37—55. — 280 с. — ISBN 5-03-001819-0.
- Janvier P. Skull of Jawless Fishes // The Skull (неопр.) / J. Hanken, B. K. Hall. — University of Chicago Press, 1993. — Т. 2: Patterns of Structural and Systematic Diversity. — С. 152—188. — ISBN 0-226-31568-1.
- Janvier, Philippe. Early Jawless Vertebrates and Cyclostome Origins (англ.) // Zoological Science. — Zoological Society of Japan, 2008. — Vol. 25, no. 10. — P. 1045—1056. — doi:10.2108/zsj.25.1045. — PMID 19267641.
- Nelson J. S. Fishes of the World (неопр.). — 4. — John Wiley & Sons, 2006. — С. 704. — ISBN 978-0-471-25031-9.
Ссылки
- Pteraspidomorphi Overview (англ.). Palaeos: Life Through Deep Time. — обзор парноноздрёвых со ссылками на страницы про дочерние таксоны. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Cephalaspidomorphi Overview (англ.). Palaeos: Life Through Deep Time. — обзор непарноноздрёвых со ссылками на страницы про дочерние таксоны. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Janvier, Philippe. Vertebrata. Animals with backbones. Version 01 January 1997 (under construction) (англ.). The Tree of Life Web Project. — Обзор позвоночных со ссылками на страницы про дочерние таксоны. Дата обращения: 21 ноября 2012. Архивировано 11 декабря 2012 года.
- Шабанов Д. А. III. Vertebrata-01. Бесчелюстные. Лекторий-библиотека Batrachos.com. Дата обращения: 21 ноября 2012. Архивировано 23 января 2013 года.
- Остракодермы на марках разных стран. biostamps.narod.ru. Дата обращения: 21 ноября 2012.
| Таксономия |
|---|